
Interaction Between Malat1 and miR-499-5p Regulates Meis1 Expression and Function with a Net Impact on Cell Proliferation
 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Oligonucleotide Transfection
2.2. RNA Extraction and Quantification
2.3. Reverse Transcription and qPCR
2.4. Shotgun Proteomics Analysis
2.5. Nano LC-MS/MS Analysis
2.6. Proteomics Data Analysis
2.7. Western Blotting
2.8. Luciferase Reporter Assay
2.9. BrdU Incorporation Assay
2.10. Bioinformatic and Statistical Analysis
3. Results
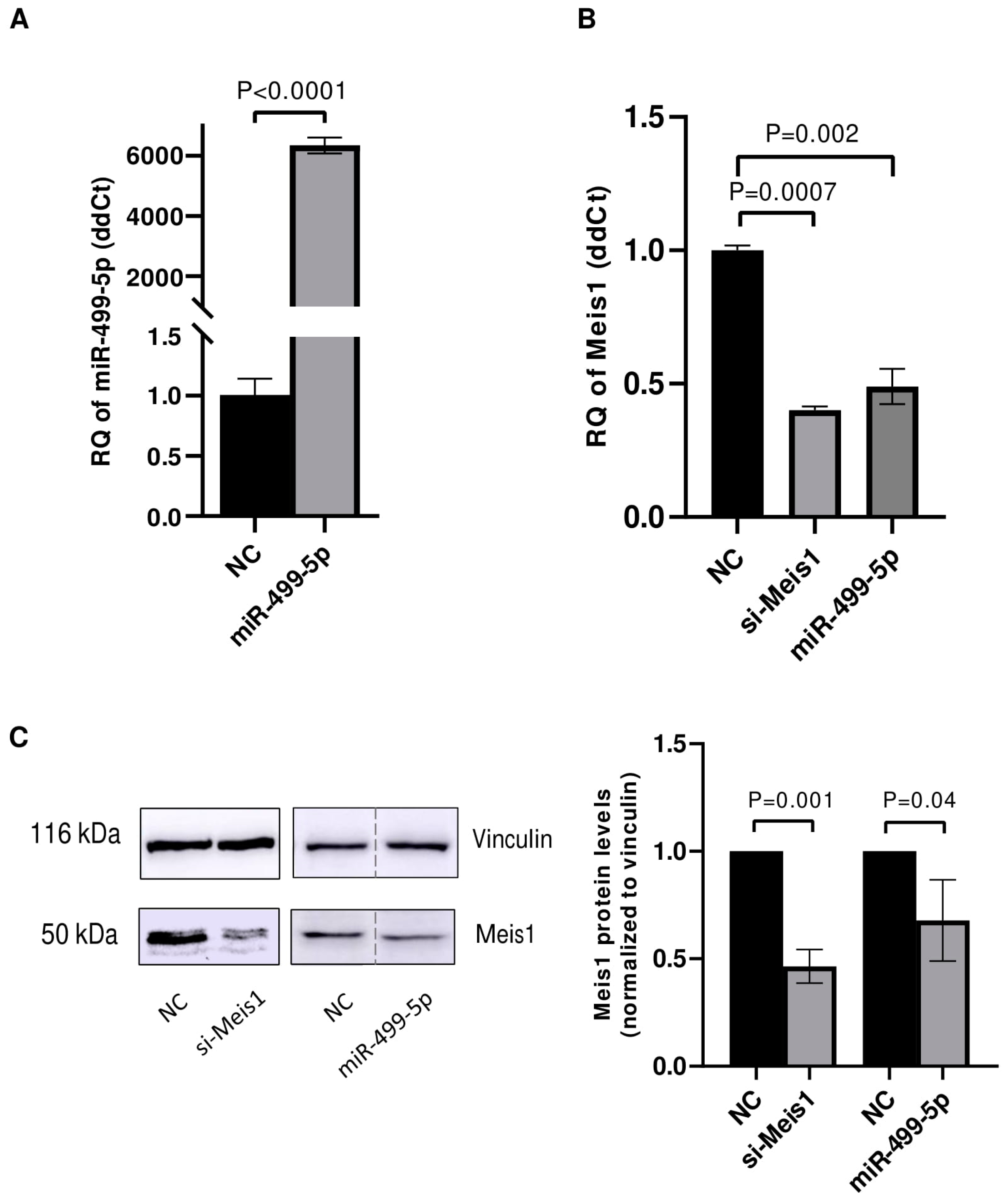
3.1. miR-499-5p Acts as an Upstream Regulator of Meis1
3.2. miR-499-5p Affects Meis1 mRNA and Protein Expression
3.3. Malat1 Sponges miR-499-5p to Regulate Meis1 mRNA and Protein Expression
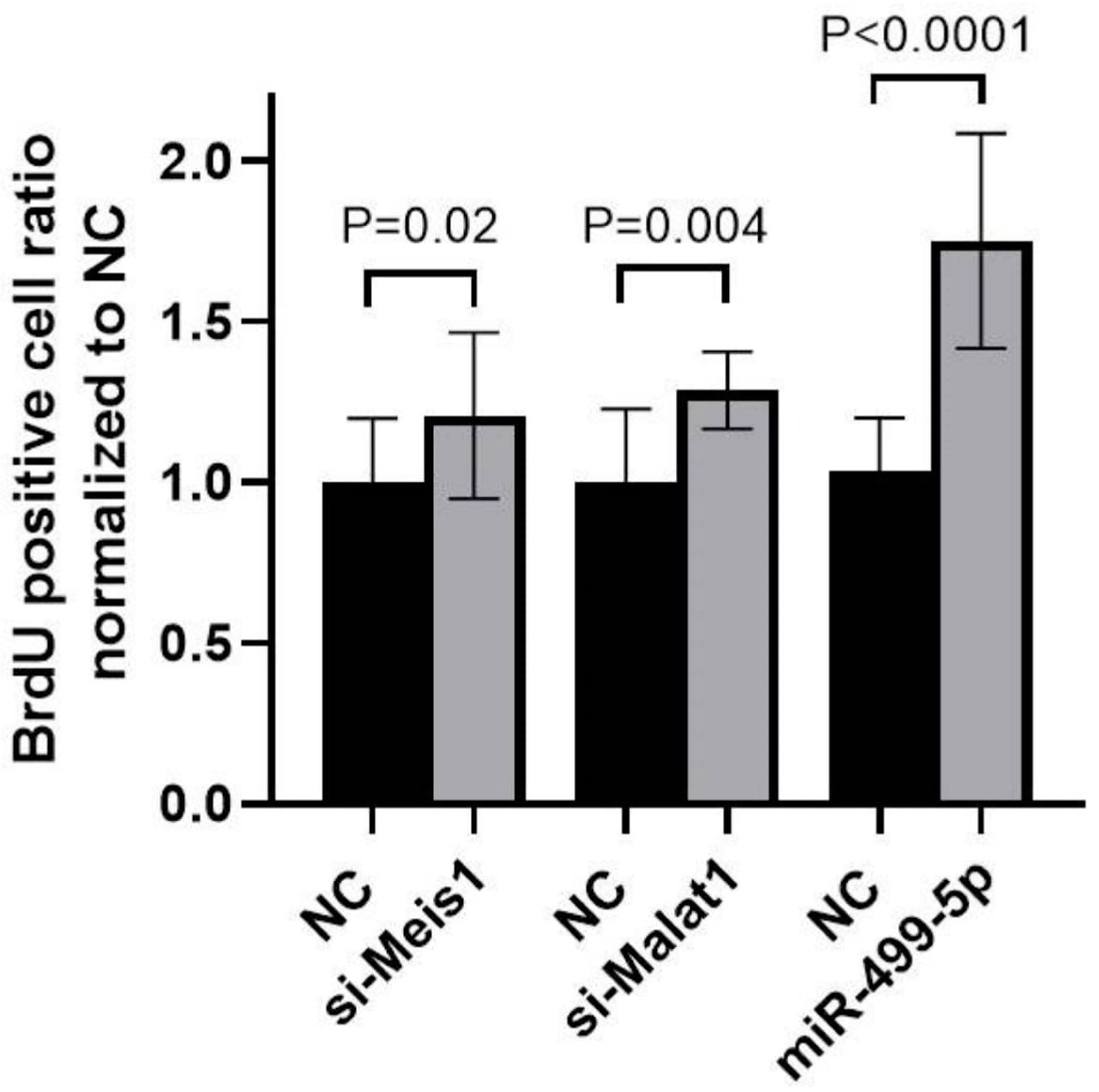
3.4. Impact of Malat1, miR-499-5p and Meis1 on Cell Proliferation
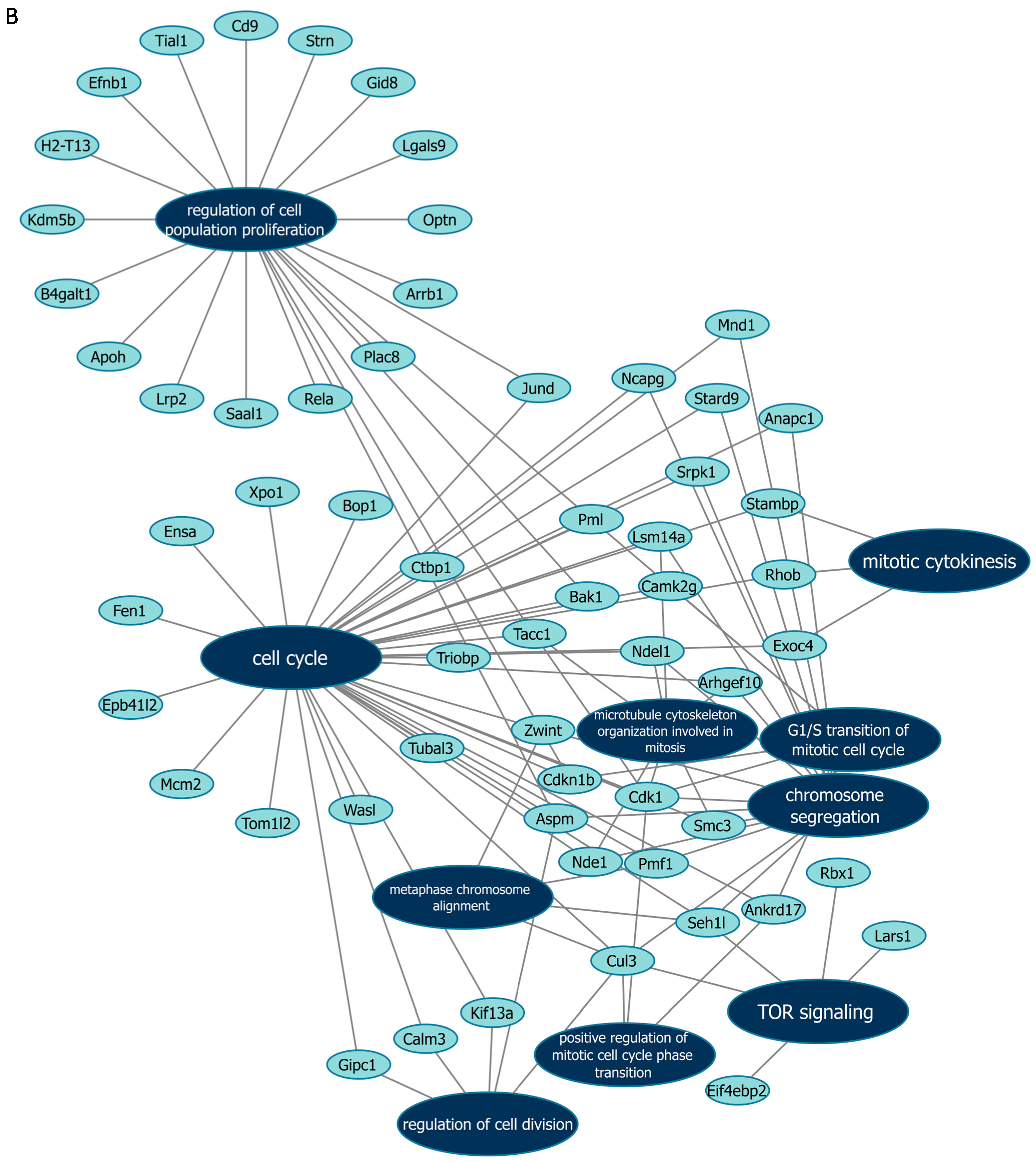
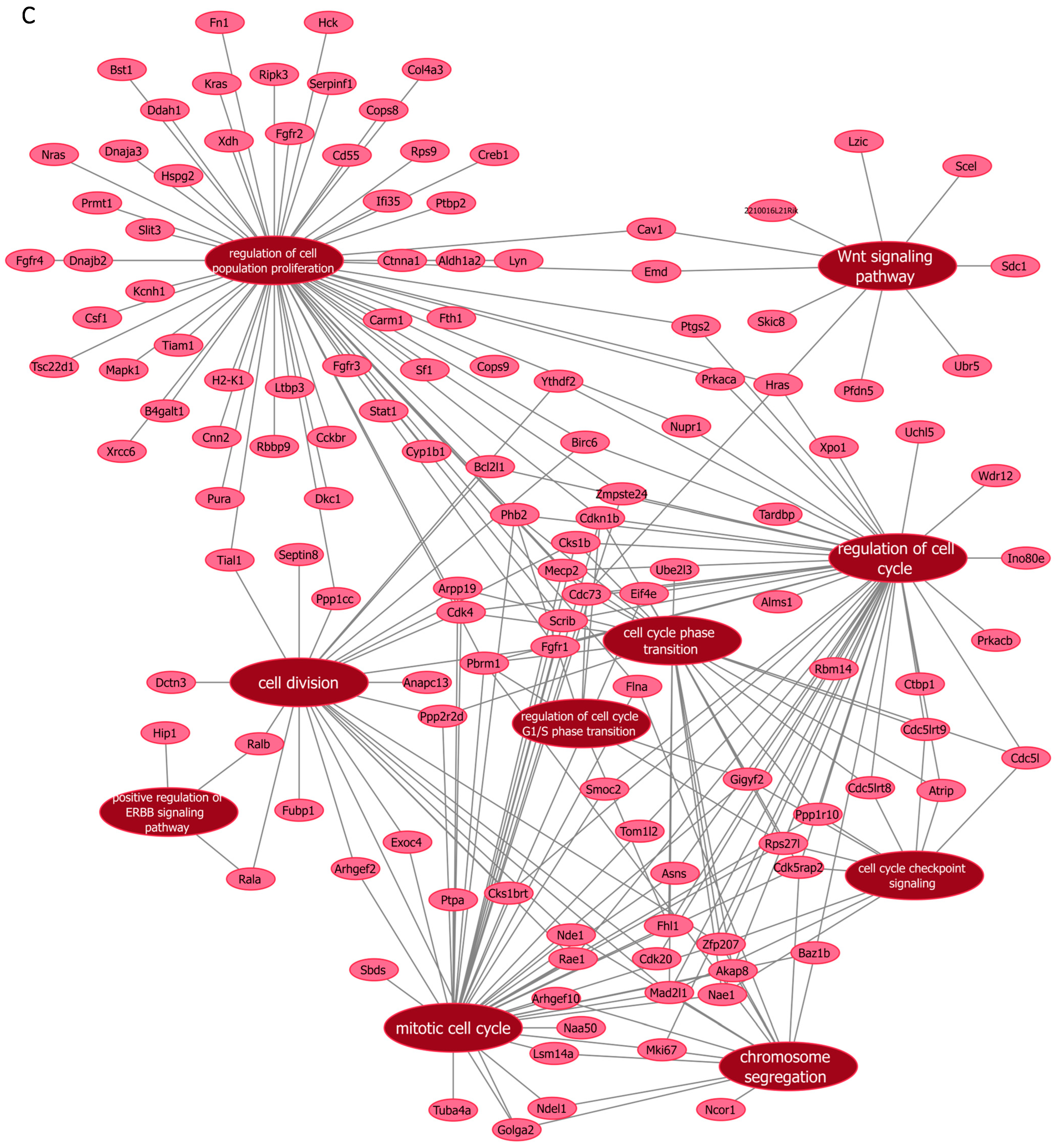
3.5. Functional Enrichment Analyses Using Mass Spectrometry After Manipulation of Malat1, miR-499-5p and Meis1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moskow, J.J.; Bullrich, F.; Huebner, K.; Daar, I.O.; Buchberg, A.M. Meis1, a PBX1-Related Homeobox Gene Involved in Myeloid Leukemia in BXH-2 Mice. Mol. Cell. Biol. 1995, 15, 5434–5443. [Google Scholar] [CrossRef]
- Jiang, M.; Xu, S.; Bai, M.; Zhang, A. The Emerging Role of MEIS1 in Cell Proliferation and Differentiation. Am. J. Physiol. Cell Physiol. 2021, 320, C264–C269. [Google Scholar] [CrossRef]
- Cardoso, A.C.; Pereira, A.H.M.; Sadek, H.A. Mechanisms of Neonatal Heart Regeneration. Curr. Cardiol. Rep. 2020, 22, 33. [Google Scholar] [CrossRef]
- Salminen, A.V.; Lam, D.D.; Winkelmann, J. Role of MEIS1 in Restless Legs Syndrome: From GWAS to Functional Studies in Mice. Adv. Pharmacol. 2019, 84, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Spieker, N.; Van Sluis, P.; Beitsma, M.; Boon, K.; Van Schaik, B.D.C.; Van Kampen, A.H.C.; Caron, H.; Versteeg, R. The MEIS1 Oncogene Is Highly Expressed in Neuroblastoma and Amplified in Cell Line IMR32. Genomics 2001, 71, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.L.; Li, J.; Kiriluk, K.J.; Rosen, A.M.; Paner, G.P.; Antic, T.; Lussier, Y.A.; Vander Griend, D.J. Deregulation of a Hox Protein Regulatory Network Spanning Prostate Cancer Initiation and Progression. Clin. Cancer Res. 2012, 18, 4291–4302. [Google Scholar] [CrossRef] [PubMed]
- Crijns, A.P.G.; de Graeff, P.; Geerts, D.; ten Hoor, K.A.; Hollema, H.; van der Sluis, T.; Hofstra, R.M.W.; de Bock, G.H.; de Jong, S.; van der Zee, A.G.J.; et al. MEIS and PBX Homeobox Proteins in Ovarian Cancer. Eur. J. Cancer 2007, 43, 2495–2505. [Google Scholar] [CrossRef]
- Li, W.; Huang, K.; Guo, H.; Cui, G. Meis1 Regulates Proliferation of Non-Small-Cell Lung Cancer Cells. J. Thorac. Dis. 2014, 6, 850. [Google Scholar] [CrossRef]
- Yao, M.; Gu, Y.; Yang, Z.; Zhong, K.; Chen, Z. MEIS1 and Its Potential as a Cancer Therapeutic Target (Review). Int. J. Mol. Med. 2021, 48, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, T.; Nakatake, M.; Kuwata, T.; Couzinet, A.; Goitsuka, R.; Tsutsumi, S.; Aburatani, H.; Valk, P.J.M.; Delwel, R.; Nakamura, T. MEIS1-Mediated Transactivation of Synaptotagmin-like 1 Promotes CXCL12/CXCR4 Signaling and Leukemogenesis. J. Clin. Investig. 2016, 126, 1664–1678. [Google Scholar] [CrossRef]
- Liu, J.; Qin, Y.Z.; Yang, S.; Wang, Y.; Chang, Y.J.; Zhao, T.; Jiang, Q.; Huang, X.J. Meis1 Is Critical to the Maintenance of Human Acute Myeloid Leukemia Cells Independent of MLL Rearrangements. Ann. Hematol. 2017, 96, 567–574. [Google Scholar] [CrossRef]
- Rosales-Avĩa, J.A.; Torres-Flores, J.; Aguilar-Lemarroy, A.; Gurrola-Díaz, C.; Hernández-Flores, G.; Ortiz-Lazareno, P.C.; Lerma-Díaz, J.M.; De Celis, R.; González-Ramella, Ó.; Barrera-Chaires, E.; et al. MEIS1, PREP1, and PBX4 Are Differentially Expressed in Acute Lymphoblastic Leukemia: Association of MEIS1 Expression with Higher Proliferation and Chemotherapy Resistance. J. Exp. Clin. Cancer Res. 2011, 30, 112. [Google Scholar] [CrossRef]
- Crist, R.C.; Roth, J.J.; Waldman, S.A.; Buchberg, A.M. A Conserved Tissue-Specific Homeodomain-Less Isoform of MEIS1 Is Downregulated in Colorectal Cancer. PLoS ONE 2011, 6, e23665. [Google Scholar] [CrossRef]
- Song, F.; Wang, H.; Wang, Y. Myeloid Ecotropic Viral Integration Site 1 Inhibits Cell Proliferation, Invasion or Migration in Human Gastric Cancer. Oncotarget 2017, 8, 90050–90060. [Google Scholar] [CrossRef]
- Mahmoud, A.I.; Kocabas, F.; Muralidhar, S.A.; Kimura, W.; Koura, A.S.; Thet, S.; Porrello, E.R.; Sadek, H.A. Meis1 Regulates Postnatal Cardiomyocyte Cell Cycle Arrest. Nature 2013, 497, 249. [Google Scholar] [CrossRef]
- Nguyen, N.U.N.; Canseco, D.C.; Xiao, F.; Nakada, Y.; Li, S.; Lam, N.T.; Muralidhar, S.A.; Savla, J.J.; Hill, J.A.; Le, V.; et al. A Calcineurin–Hoxb13 Axis Regulates Growth Mode of Mammalian Cardiomyocytes. Nature 2020, 582, 271–276. [Google Scholar] [CrossRef]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96. [Google Scholar] [CrossRef]
- Sufianov, A.; Beilerli, A.; Begliarzade, S.; Ilyasova, T.; Kudriashov, V.; Liang, Y.; Beylerli, O. The Role of Noncoding RNAs in the Osteogenic Differentiation of Human Periodontal Ligament-Derived Cells. Noncoding RNA Res. 2022, 8, 89–95. [Google Scholar] [CrossRef]
- Lanzillotti, C.; De Mattei, M.; Mazziotta, C.; Taraballi, F.; Rotondo, J.C.; Tognon, M.; Martini, F. Long Non-Coding RNAs and MicroRNAs Interplay in Osteogenic Differentiation of Mesenchymal Stem Cells. Front. Cell Dev. Biol. 2021, 9, 646032. [Google Scholar] [CrossRef]
- Subhramanyam, C.S.; Hu, Q. Non-Coding RNA in Brain Development and Disorder. Curr. Med. Chem. 2017, 24, 1983–1997. [Google Scholar] [CrossRef]
- Shi, H.; He, Y.; Li, X.; Du, Y.; Zhao, J.; Ge, C. Regulation of Non-Coding RNA in the Growth and Development of Skeletal Muscle in Domestic Chickens. Genes 2022, 13, 1033. [Google Scholar] [CrossRef] [PubMed]
- Bueno, M.J.; Malumbres, M. MicroRNAs and the Cell Cycle. Biochim. Biophys. Acta 2011, 1812, 592–601. [Google Scholar] [CrossRef]
- Stojic, L.; Lun, A.T.L.; Mascalchi, P.; Ernst, C.; Redmond, A.M.; Mangei, J.; Barr, A.R.; Bousgouni, V.; Bakal, C.; Marioni, J.C.; et al. A High-Content RNAi Screen Reveals Multiple Roles for Long Noncoding RNAs in Cell Division. Nat. Commun. 2020, 11, 1851. [Google Scholar] [CrossRef] [PubMed]
- Hime, G.R.; Somers, W.G. Micro-RNA Mediated Regulation of Proliferation, Self-Renewal and Differentiation of Mammalian Stem Cells. Cell Adhes. Migr. 2009, 3, 425–432. [Google Scholar] [CrossRef]
- Hwang, H.W.; Mendell, J.T. MicroRNAs in Cell Proliferation, Cell Death, and Tumorigenesis. Br. J. Cancer 2006, 94, 776–780. [Google Scholar] [CrossRef] [PubMed]
- De Martino, M.; Esposito, F.; Pallante, P. Long Non-Coding RNAs Regulating Multiple Proliferative Pathways in Cancer Cell. Transl. Cancer Res. 2021, 10, 3140–3157. [Google Scholar] [CrossRef] [PubMed]
- Romania, P.; Lulli, V.; Pelosi, E.; Biffoni, M.; Peschle, C.; Marziali, G. MicroRNA 155 Modulates Megakaryopoiesis at Progenitor and Precursor Level by Targeting Ets-1 and Meis1 Transcription Factors. Br. J. Haematol. 2008, 143, 570–580. [Google Scholar] [CrossRef]
- Palma, C.A.; Al Sheikha, D.; Lim, T.K.; Bryant, A.; Vu, T.T.; Jayaswal, V.; Ma, D.D.F. MicroRNA-155 as an Inducer of Apoptosis and Cell Differentiation in Acute Myeloid Leukaemia. Mol. Cancer 2014, 13, 79. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Huang, H.; Chen, P.; He, M.; Li, Y.; Arnovitz, S.; Jiang, X.; He, C.; Hyjek, E.; Zhang, J.; et al. MiR-196b Directly Targets Both HOXA9/MEIS1 Oncogenes and FAS Tumour Suppressor in MLL-Rearranged Leukaemia. Nat. Commun. 2012, 3, 688. [Google Scholar] [CrossRef]
- Su, Z.; Si, W.; Li, L.; Zhou, B.; Li, X.; Xu, Y.; Xu, C.; Jia, H.; Wang, Q.K. MiR-144 Regulates Hematopoiesis and Vascular Development by Targeting Meis1 during Zebrafish Development. Int. J. Biochem. Cell Biol. 2014, 49, 53–63. [Google Scholar] [CrossRef]
- Pandey, R.; Yang, Y.; Jackson, L.; Ahmed, R.P.H. MicroRNAs Regulating Meis1 Expression and Inducing Cardiomyocyte Proliferation. Cardiovasc. Regen. Med. 2016, 3, e1468. [Google Scholar]
- Li, Y.; Gan, Y.; Liu, J.; Li, J.; Zhou, Z.; Tian, R.; Sun, R.; Liu, J.; Xiao, Q.; Li, Y.; et al. Downregulation of MEIS1 Mediated by ELFN1-AS1/EZH2/DNMT3a Axis Promotes Tumorigenesis and Oxaliplatin Resistance in Colorectal Cancer. Signal Transduct. Target. Ther. 2022, 7, 87. [Google Scholar] [CrossRef]
- Sameh, M.; Khalaf, H.M.; Anwar, A.M.; Osama, A.; Ahmed, E.A.; Mahgoub, S.; Ezzeldin, S.; Tanios, A.; Alfishawy, M.; Said, A.F.; et al. Integrated Multiomics Analysis to Infer COVID-19 Biological Insights. Sci. Rep. 2023, 13, 1802. [Google Scholar] [CrossRef]
- Zhou, X.; Zheng, B.; Pang, L.; Che, Y.; Qi, X. Suppression of MALAT1 Alleviates Neurocyte Apoptosis and Reactive Oxygen Species Production through the MiR-499-5p/SOX6 Axis in Subarachnoid Hemorrhage. J. Mol. Histol. 2022, 53, 85–96. [Google Scholar] [CrossRef]
- Chuang, C.-Y.; Wang, B.-W.; Yu, Y.-J.; Fang, W.-J.; Lin, C.-M.; Shyu, K.-G.; Chua, S.-K. Exosomal MALAT1 from Rapid Electrical Stimulation-Treated Atrial Fibroblasts Enhances Sox-6 Expression by Downregulating MiR-499a-5p. Cells 2024, 13, 1942. [Google Scholar] [CrossRef] [PubMed]
- Grant, S.; Qiao, L.; Dent, P. Roles of ERBB Family Receptor Tyrosine Kinases, and Downstream Signaling Pathways, in the Control of Cell Growth and Survival. Front. Biosci. 2002, 7, d376–d389. [Google Scholar] [CrossRef] [PubMed]
- Holbro, T.; Civenni, G.; Hynes, N.E. The ErbB Receptors and Their Role in Cancer Progression. Exp. Cell Res. 2003, 284, 99–110. [Google Scholar] [CrossRef]
- Staal, F.J.T.; Luis, T.C. Wnt Signaling in Hematopoiesis: Crucial Factors for Self-Renewal, Proliferation, and Cell Fate Decisions. J. Cell Biochem. 2010, 109, 844–849. [Google Scholar] [CrossRef]
- Davidson, G. The Cell Cycle and Wnt. Cell Cycle 2010, 9, 1667–1668. [Google Scholar] [CrossRef]
- Wei, Z.; Liu, H.T. MAPK Signal Pathways in the Regulation of Cell Proliferation in Mammalian Cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef]
- Fingar, D.C.; Blenis, J. Target of Rapamycin (TOR): An Integrator of Nutrient and Growth Factor Signals and Coordinator of Cell Growth and Cell Cycle Progression. Oncogene 2004, 23, 3151–3171. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Liu, R.-D.; Lei, D.-Q.; Shang, Q.-C.; Li, H.-F.; Hu, X.-G.; Zheng, H.; Jin, G. MiR-499a-5p Promotes 5-FU Resistance and the Cell Proliferation and Migration through Activating PI3K/Akt Signaling by Targeting PTEN in Pancreatic Cancer. Ann. Transl. Med. 2021, 9, 1798. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.H.; Zhang, S.G.; Zhou, P.; Wei, X.; Mao, X.D.; Lin, S.G.; Liu, C. LncRNA MALAT1 Affects High Glucose-Induced Endothelial Cell Proliferation, Apoptosis, Migration and Angiogenesis by Regulating the PI3K/Akt Signaling Pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8551–8559. [Google Scholar] [CrossRef]
- Gutschner, T.; Hämmerle, M.; Diederichs, S. MALAT1—A Paradigm for Long Noncoding RNA Function in Cancer. J. Mol. Med. 2013, 91, 791–801. [Google Scholar] [CrossRef]
- Blasi, F.; Bruckmann, C.; Penkov, D.; Dardaei, L. A Tale of TALE, PREP1, PBX1, and MEIS1: Interconnections and Competition in Cancer. Bioessays 2017, 39, 1600245. [Google Scholar] [CrossRef]
- Aksoz, M.; Turan, R.D.; Albayrak, E.; Kocabas, F. Emerging Roles of Meis1 in Cardiac Regeneration, Stem Cells and Cancer. Curr. Drug Targets 2018, 19, 181–190. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed]
- YiRen, H.; YingCong, Y.; Sunwu, Y.; Keqin, L.; Xiaochun, T.; Senrui, C.; Ende, C.; XiZhou, L.; Yanfan, C. Long Noncoding RNA MALAT1 Regulates Autophagy Associated Chemoresistance via MiR-23b-3p Sequestration in Gastric Cancer. Mol. Cancer 2017, 16, 174. [Google Scholar] [CrossRef]
- Su, K.; Wang, N.; Shao, Q.; Liu, H.; Zhao, B.; Ma, S. The Role of a CeRNA Regulatory Network Based on LncRNA MALAT1 Site in Cancer Progression. Biomed. Pharmacother. 2021, 137, 111389. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Lin, C.; Liu, W.; Zhang, J.; Ohgi, K.A.; Grinstein, J.D.; Dorrestein, P.C.; Rosenfeld, M.G. NcRNA- and Pc2 Methylation-Dependent Gene Relocation between Nuclear Structures Mediates Gene Activation Programs. Cell 2011, 147, 773–788. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, V.; Shen, Z.; Chakraborty, A.; Giri, S.; Freier, S.M.; Wu, X.; Zhang, Y.; Gorospe, M.; Prasanth, S.G.; Lal, A.; et al. Long Noncoding RNA MALAT1 Controls Cell Cycle Progression by Regulating the Expression of Oncogenic Transcription Factor B-MYB. PLoS Genet. 2013, 9, e1003368. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, J.N.; Ensminger, A.W.; Clemson, C.M.; Lynch, C.R.; Lawrence, J.B.; Chess, A. A Screen for Nuclear Transcripts Identifies Two Linked Noncoding RNAs Associated with SC35 Splicing Domains. BMC Genom. 2007, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, V.; Ellis, J.D.; Shen, Z.; Song, D.Y.; Pan, Q.; Watt, A.T.; Freier, S.M.; Bennett, C.F.; Sharma, A.; Bubulya, P.A.; et al. The Nuclear-Retained Noncoding RNA MALAT1 Regulates Alternative Splicing by Modulating SR Splicing Factor Phosphorylation. Mol. Cell 2010, 39, 925–938. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ning, J.; Teng, H.; Deng, Y.; Sheldon, M.; Shi, L.; Martinez, C.; Zhang, J.; Tian, A.; Sun, Y.; et al. Long Noncoding RNA Malat1 Protects against Osteoporosis and Bone Metastasis. Nat. Commun. 2024, 15, 2384. [Google Scholar] [CrossRef]
- Chen, F.; Li, W.; Zhang, D.; Fu, Y.; Yuan, W.; Luo, G.; Liu, F.; Luo, J. MALAT1 Regulates Hypertrophy of Cardiomyocytes by Modulating the MiR-181a/HMGB2 Pathway. Eur. J. Histochem. 2022, 66. [Google Scholar] [CrossRef]
- Sun, Y.; Gu, R.; Shen, Z.; Ye, Y. MALAT1 Knockdown Alleviates Myocardial Injury in Mice with Severe Acute Pancreatitis via the MiR-374a/Sp1/Wnt/β-Catenin Pathway. Am. J. Transl. Res. 2023, 15, 3928. [Google Scholar]
- Hajibabaei, S.; Nafissi, N.; Azimi, Y.; Mahdian, R.; Rahimi-Jamnani, F.; Valizadeh, V.; Rafiee, M.H.; Azizi, M. Targeting Long Non-Coding RNA MALAT1 Reverses Cancerous Phenotypes of Breast Cancer Cells through MicroRNA-561-3p/TOP2A Axis. Sci. Rep. 2023, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Terroba, E.; Plasek-Hegde, L.M.; Chiotakakos, I.; Li, V.; de Miguel, F.J.; Robles-Oteiza, C.; Tyagi, A.; Politi, K.; Zamudio, J.R.; Dimitrova, N. Overexpression of Malat1 Drives Metastasis through Inflammatory Reprogramming of the Tumor Microenvironment. Sci. Immunol. 2024, 9, eadh5462. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, X.; Wu, H.; Ni, P.; Gu, Z.; Qiao, Y.; Chen, N.; Sun, F.; Fan, Q. CREB Up-Regulates Long Non-Coding RNA, HULC Expression through Interaction with MicroRNA-372 in Liver Cancer. Nucleic Acids Res. 2010, 38, 5366. [Google Scholar] [CrossRef]
- Jiang, X.; Zhou, Y.; Sun, A.J.; Xue, J.L. NEAT1 Contributes to Breast Cancer Progression through Modulating MiR-448 and ZEB1. J. Cell Physiol. 2018, 233, 8558–8566. [Google Scholar] [CrossRef] [PubMed]
- Ghini, F.; Rubolino, C.; Climent, M.; Simeone, I.; Marzi, M.J.; Nicassio, F. Endogenous Transcripts Control MiRNA Levels and Activity in Mammalian Cells by Target-Directed MiRNA Degradation. Nat. Commun. 2018, 9, 3119. [Google Scholar] [CrossRef]
- Kleaveland, B.; Shi, C.Y.; Stefano, J.; Bartel, D.P. A Network of Noncoding Regulatory RNAs Acts in the Mammalian Brain. Cell 2018, 174, 350–362.e17. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Chang, M.W.; Tsitsipatis, D.; Yang, X.; Martindale, J.L.; Munk, R.; Cheng, A.; Izydore, E.; Pandey, P.R.; Piao, Y.; et al. LncRNA OIP5-AS1-Directed MiR-7 Degradation Promotes MYMX Production during Human Myogenesis. Nucleic Acids Res. 2022, 50, 7115–7133. [Google Scholar] [CrossRef]
- de la Mata, M.; Gaidatzis, D.; Vitanescu, M.; Stadler, M.B.; Wentzel, C.; Scheiffele, P.; Filipowicz, W.; Großhans, H. Potent Degradation of Neuronal Mi RNA s Induced by Highly Complementary Targets. EMBO Rep. 2015, 16, 500–511. [Google Scholar] [CrossRef]
- Bosson, A.D.; Zamudio, J.R.; Sharp, P.A. Endogenous MiRNA and Target Concentrations Determine Susceptibility to Potential CeRNA Competition. Mol. Cell 2014, 56, 347–359. [Google Scholar] [CrossRef]
- Denzler, R.; McGeary, S.E.; Title, A.C.; Agarwal, V.; Bartel, D.P.; Stoffel, M. Impact of MicroRNA Levels, Target-Site Complementarity, and Cooperativity on Competing Endogenous RNA-Regulated Gene Expression. Mol. Cell 2016, 64, 565–579. [Google Scholar] [CrossRef]
- Sluijter, J.P.G.; Van Mil, A.; Van Vliet, P.; Metz, C.H.G.; Liu, J.; Doevendans, P.A.; Goumans, M.J. MicroRNA-1 and-499 Regulate Differentiation and Proliferation in Human-Derived Cardiomyocyte Progenitor Cells. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Huang, L.; Su, P.; Song, T.; Zhang, W.; Fan, J.; Liu, Y. MicroRNA-499a-5p Inhibits Osteosarcoma Cell Proliferation and Differentiation by Targeting Protein Phosphatase 1D through Protein Kinase B/Glycogen Synthase Kinase 3β Signaling. Oncol. Lett. 2018, 15, 4113–4120. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Dong, M.; Liu, Z.; Yang, J.; Shi, Y. MiR-499a-5p Inhibits Proliferation, Invasion, Migration, and Epithelial-Mesenchymal Transition, and Enhances Radiosensitivity of Cervical Cancer Cells via Targeting EIF4E. Onco Targets Ther. 2020, 13, 2913–2924. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Shi, P.; Jia, X.; Xue, Q. Long Non-coding RNA LINC01018 Inhibits the Progression of Acute Myeloid Leukemia by Targeting MiR-499a-5p to Regulate PDCD4. Oncol. Lett. 2021, 22, 1–11. [Google Scholar] [CrossRef]
- Zong, Z.; Song, Y.; Xue, Y.; Ruan, X.; Liu, X.; Yang, C.; Zheng, J.; Cao, S.; Li, Z.; Liu, Y. Knockdown of LncRNA SCAMP1 Suppressed Malignant Biological Behaviours of Glioma Cells via Modulating MiR-499a-5p/LMX1A/NLRC5 Pathway. J. Cell Mol. Med. 2019, 23, 5048–5062. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Li, Z.; Yu, Y.; Zeng, Q.; Cheng, Y.; Ji, W.; Xia, W.; Lu, S. Exosomal MiR-499a-5p Promotes Cell Proliferation, Migration and EMT via MTOR Signaling Pathway in Lung Adenocarcinoma. Exp. Cell Res. 2019, 379, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.; Yang, Z.; Feng, Z.; Wang, Y.; Ji, N. MicroRNA-499-5p Promotes Vascular Smooth Muscle Cell Proliferation and Migration via Inhibiting SOX6. Physiol. Genom. 2023, 55, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Aslan, G.S.; Jaé, N.; Manavski, Y.; Fouani, Y.; Shumliakivska, M.; Kettenhausen, L.; Kirchhof, L.; Günther, S.; Fischer, A.; Luxán, G.; et al. Malat1 Deficiency Prevents Neonatal Heart Regeneration by Inducing Cardiomyocyte Binucleation. JCI Insight 2023, 8. [Google Scholar] [CrossRef]
- Chen, Y.; Li, S.; Zhang, Y.; Wang, M.; Li, X.; Liu, S.; Xu, D.; Bao, Y.; Jia, P.; Wu, N.; et al. The LncRNA Malat1 Regulates Microvascular Function after Myocardial Infarction in Mice via MiR-26b-5p/Mfn1 Axis-Mediated Mitochondrial Dynamics. Redox Biol. 2021, 41, 101910. [Google Scholar] [CrossRef]
- Wang, S.; Ren, L.; Shen, G.; Liu, M.; Luo, J. The Knockdown of MALAT1 Inhibits the Proliferation, Invasion and Migration of Hemangioma Endothelial Cells by Regulating MiR-206/VEGFA Axis. Mol. Cell Probes 2020, 51, 101540. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wang, Y.; Hou, W.; Ding, X.; Wang, W. Long Non-Coding RNA MALAT1 Promotes Cell Proliferation, Migration and Invasion by Targeting MiR-590-3p in Osteosarcoma. Exp. Ther. Med. 2022, 24, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.A.; Valinezhad, K.; Adel, E.; Munirathinam, G. MALAT-1 Is a Key Regulator of Epithelial-Mesenchymal Transition in Cancer: A Potential Therapeutic Target for Metastasis. Cancers 2024, 16, 234. [Google Scholar] [CrossRef]
- Zhang, L.S.; Kang, X.; Lu, J.; Zhang, Y.; Wu, X.; Wu, G.; Zheng, J.; Tuladhar, R.; Shi, H.; Wang, Q.; et al. Installation of a Cancer Promoting WNT/SIX1 Signaling Axis by the Oncofusion Protein MLL-AF9. EBioMedicine 2019, 39, 145–158. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligonucleotide Name | Sequence (5′ to 3′) |
|---|---|
| MEIS1-499 WT1 F | CAGCTTGGAAATCAACAGTCTTATTACCTCATCATGGAT |
| MEIS1-499 WT1 R | CTAGATCCATGATGAGGTAATAAGACTGTTGATTTCCAAGCTGAGCT |
| MEIS1-499 WT2 F | CGCTGTTGCAGGCAGTGTCTTAAGGAGACTGGTAGGAGT |
| MEIS1-499 WT2 R | CTAGACTCCTACCAGTCTCCTTAAGACACTGCCTGCAACAGCGAGCT |
| MEIS1-499 Mut2 F | CGCTGTTGCAGGCAGTGGAGACTGGTAGGAGT |
| MEIS1-499 Mut2 R | CTAGACTCCTACCAGTCTCCACTGCCTGCAACAGCGAGCT |
| Manipulation | GO Term | Gene Symbol |
|---|---|---|
| si-Malat1 | Cell cycle G1/S phase transition | Rrm2, Cdk2, Eif4g1, Gigyf2, Men1, Plrg1 |
| Cell cycle G2/M phase transition | Calm3, Usp47, Mta3, Arpp19, Cdk2, Mre11a | |
| Cell division | Cul3, Specc1l, Calm3, Epb41l2, Exoc8, Fubp1, Igf2, Pmf1, Anapc7, Arpp19, Cks1b, Cdk2, Cks1brt, Exoc4, Men1, Nde1, Mapre2, Washc5, Wnk1, Zwint | |
| DNA replication | Hras, Rrm2, Faf1, Cdk2, Pole4, Carm1, Mre11a | |
| ERK1 and ERK2 cascade | Arrb1, Fermt2, Hras, Crkl, Map2k2, Trf | |
| MAPK cascade | Arrb1, Nenf, Fermt2, Qars1, Gdf6, Hras, Igf2, Crkl, Map2k2, Gsn, Men1, Nrk, Pak4, Trf | |
| Mitotic cell cycle | Cul3, Calm3, Exoc8, Igf2, Usp47, Mta3, Rrm2, Rnf2, Anapc7, Arpp19, Cks1b, Cdk2, Cks1brt, Eif4g1, Exoc4, Gigyf2, Men1, Nde1, Mre11a, Plrg1, Ppp1r10, Wnk1, Zwint | |
| Positive regulation of cell division | Cul3, Igf2, Men1, Wnk1 | |
| Regulation of cell population proliferation | Arrb1, Serpine2, Celf1, Efnb2, Fermt2, Hras, Kras, Igf2, Mta3, Pdcd2, Smarca2, Sp1, Ccn4, Crkl, Cdk2, Carm1, Ifi35, Men1, Lims2, Nrk, Prdx4, Ptbp2, Pramel20, Ctcf, Trf | |
| Chromosome separation | Cul3, Anapc7, Csnk2a2, Plscr1, Zwint | |
| miR-499-5p | G1/S transition of mitotic cell cycle | Ankrd17, Camk2g, Cdk1, Cdkn1b, Pml |
| Chromosome segregation | Anapc1, Cdk1, Smc3, Lsm14a, Pmf1, Arhgef10, Aspm, Mnd1, Ndel1, Nde1, Zwint, Cul3, Ncapg, Srpk1, Seh1l, Stard9 | |
| Cell cycle | Ankrd17, Ctbp1, Ensa, Fen1, Camk2g, Anapc1, Tacc1, Mcm2, Cdk1, Jund, Cdkn1b, Smc3, Lsm14a, Pmf1, Bak1, Arhgef10, Aspm, Mnd1, Ndel1, Nde1, Tubal3, Pml, Zwint, Xpo1, Calm3, Rhob, Exoc4, Gipc1, Cul3, Tom1l2, Wasl, Triobp, Ncapg, Bop1, Stambp, Srpk1, Epb41l2, Seh1l, Stard9, Kif13a | |
| Mitotic cytokinesis | Rhob, Exoc4, Stambp | |
| Microtubule cytoskeleton organization involved in mitosis | Tacc1, Cdk1, Smc3, Lsm14a, Arhgef10, Ndel1, Nde1 | |
| Positive regulation of mitotic cell cycle phase transition | Ankrd17, Cdk1, Cul3 | |
| Metaphase chromosome alignment | Pmf1, Zwint, Cul3, Seh1l | |
| Regulation of cell division | Aspm, Calm3, Gipc1, Cul3, Kif13a | |
| Regulation of cell population proliferation | Rela, Plac8, Cdk1, Jund, Gid8, Cdkn1b, Tial1, Bak1, Kdm5b, Aspm, Lrp2, Pml, Arrb1, Optn, Lgals9, Strn, Cd9, Efnb1, H2-T13, B4galt1, Apoh, Saal1 | |
| TOR signaling | Rbx1, Eif4ebp2, Cul3, Lars1, Seh1l | |
| si-Meis1 | Cell cycle checkpoint signaling | Cdk5rap2, Atrip, Zfp207, Rps27l, Mad2l1, Cdc5l, Cdc5lrt9, Cdc5lrt8, Ppp1r10, Nae1, Gigyf2 |
| Cell cycle phase transition | Cdk5rap2, Phb2, Atrip, Zfp207, Rps27l, Pbrm1, Mad2l1, Eif4e, Cks1b, Arpp19, Fhl1, Cdc73, Cdc5l, Ube2l3, Cdc5lrt9, Cdc5lrt8, Ppp2r2d, Ppp1r10, Nae1, Mecp2, Gigyf2, Cdkn1b, Cdk4, Akap8 | |
| Cell division | Fgfr1, Cdk20, Arhgef2, Fubp1, Zfp207, Ythdf2, Nde1, Mad2l1, Exoc4, Cks1brt, Cks1b, Birc6, Arpp19, Dctn3, Anapc13, Golga2, Ppp1cc, Tial1, Septin8, Ralb, Rala, Rae1, Ppp2r2d, Cdk4, Bcl2l1 | |
| Chromosome segregation | Cdk5rap2, Zfp207, Pbrm1, Ndel1, Nde1, Ncor1, Mad2l1, Baz1b, Arhgef10, Golga2, Flna, Mki67, Lsm14a, Akap8 | |
| Mitotic cell cycle | Ptpa, Fgfr1, Arhgef2, Cdk5rap2, Scrib, Tuba4a, Phb2, Asns, Zmpste24, Zfp207, Tom1l2, Smoc2, Sbds, Rps27l, Pbrm1, Ndel1, Nde1, Naa50, Mad2l1, Exoc4, Eif4e, Cks1brt, Cks1b, Baz1b, Arpp19, Arhgef10, Fhl1, Golga2, Flna, Cdc73, Rae1, Ppp2r2d, Ppp1r10, Nae1, Mki67, Mecp2, Lsm14a, Gigyf2, Cdkn1b, Cdk4, Akap8 | |
| Positive regulation of ERBB signaling pathway | Ralb, Rala, Hip1 | |
| Regulation of cell cycle | Prkacb, Prkaca, Hras, Fgfr1, Cdk20, Cdk5rap2, Scrib, Phb2, Atrip, Asns, Zmpste24, Zfp207, Ythdf2, Wdr12, Uchl5, Tom1l2, Smoc2, Rps27l, Ptgs2, Pbrm1, Nupr1, Mad2l1, Ino80e, Eif4e, Cks1brt, Cks1b, Birc6, Baz1b, Alms1, Rbm14, Ctbp1, Fhl1, Cdc73, Cdc5l, Cdc5lrt9, Cdc5lrt8, Xpo1, Tardbp, Rae1, Ppp1r10, Nae1, Mki67, Mecp2, Gigyf2, Cdkn1b, Cdk4, Bcl2l1 | |
| Regulation of cell cycle G1/S phase transition | Phb2, Rps27l, Pbrm1, Fhl1, Cdc73, Gigyf2, Cdkn1b | |
| Regulation of cell population proliferation | Slit3, Cnn2, Ptbp2, Prkaca, Hras, Mapk1, Lyn, Hck, Fgfr4, Fgfr3, Fgfr2, Fgfr1, Rbbp9, Rps9, Kras, Scrib, Pura, Phb2, Fn1, Tsc22d1, Zmpste24, Xdh, Tiam1, Stat1, Serpinf1, Ptgs2, Prmt1, Pbrm1, Nupr1, Nras, Ltbp3, Ifi35, Hspg2, H2-K1, Fth1, Emd, Dnaja3, Csf1, Cops9, Col4a3, Cav1, Carm1, Birc6, Kcnh1, Cckbr, Cops8, Ddah1, Flna, Dkc1, Cdc73, Bst1, Xrcc6, Ctnna1, Ppp1cc, Tial1, Sf1, Ripk3, Mecp2, Dnajb2, Cyp1b1, Creb1, Cdkn1b, Cdk4, Cd55, Bcl2l1, B4galt1, Aldh1a2 | |
| Wnt signaling pathway | 2210016L21Rik, Skic8, Pfdn5, Emd, Cav1, Ubr5, Cdc73, Sdc1, Scel, Lzic |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fahim, S.A.; Ragheb, M.; Fayed, I.H.; Osama, A.; Karam, A.; Magdeldin, S.; Metwale, R.; Elsayed, M.D.A.A.; Abdellatif, A.; Sadek, H.A.; et al. Interaction Between Malat1 and miR-499-5p Regulates Meis1 Expression and Function with a Net Impact on Cell Proliferation. Cells 2025, 14, 125. https://doi.org/10.3390/cells14020125
Fahim SA, Ragheb M, Fayed IH, Osama A, Karam A, Magdeldin S, Metwale R, Elsayed MDAA, Abdellatif A, Sadek HA, et al. Interaction Between Malat1 and miR-499-5p Regulates Meis1 Expression and Function with a Net Impact on Cell Proliferation. Cells. 2025; 14(2):125. https://doi.org/10.3390/cells14020125
Chicago/Turabian StyleFahim, Salma A., Manon Ragheb, Ibrahim Hassan Fayed, Aya Osama, Ahmed Karam, Sameh Magdeldin, Rana Metwale, Mohamed Dief Allah Abdalmoneam Elsayed, Ahmed Abdellatif, Hesham A. Sadek, and et al. 2025. "Interaction Between Malat1 and miR-499-5p Regulates Meis1 Expression and Function with a Net Impact on Cell Proliferation" Cells 14, no. 2: 125. https://doi.org/10.3390/cells14020125
APA StyleFahim, S. A., Ragheb, M., Fayed, I. H., Osama, A., Karam, A., Magdeldin, S., Metwale, R., Elsayed, M. D. A. A., Abdellatif, A., Sadek, H. A., El Sobky, S. A., El-Ekiaby, N., Fawzy, I. O., & Abdelaziz, A. I. (2025). Interaction Between Malat1 and miR-499-5p Regulates Meis1 Expression and Function with a Net Impact on Cell Proliferation. Cells, 14(2), 125. https://doi.org/10.3390/cells14020125