
Enhanced Expression of TRIM46 in Ovarian Cancer Cells Induced by Tumor-Associated Macrophages Promotes Invasion via the Wnt/β-Catenin Pathway


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture and Preparation of Conditioned Medium
2.3. Invasion Assay
2.4. Cell Viability Assay
2.5. mRNA Sequencing
2.6. Bioinformatics Analysis
2.7. Real-Time RT-PCR
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. TRIM46 Is Upregulated in Macrophage-Stimualted Ovarian Cancer Cells and Correlates with Poor Prognosis
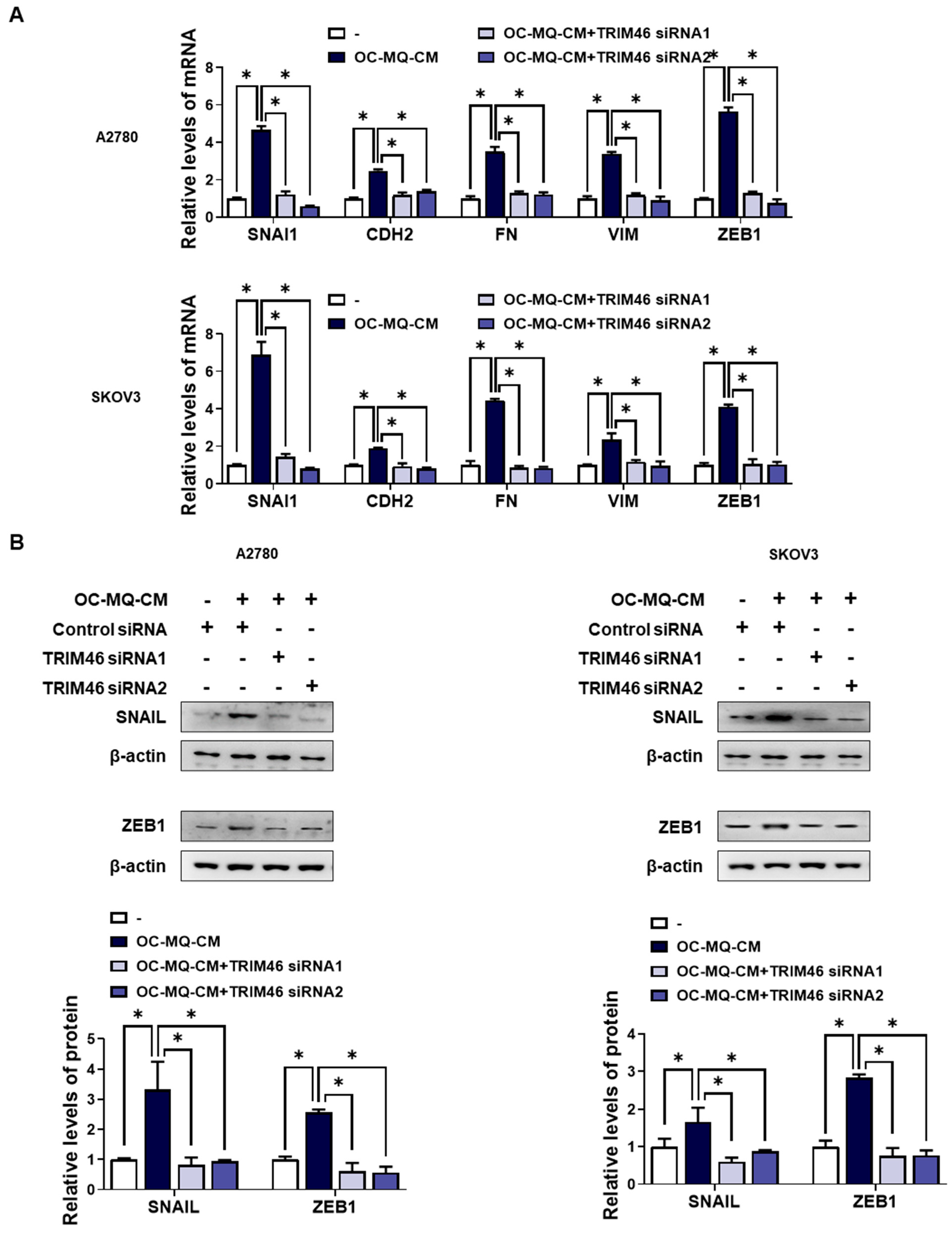
3.2. TRIM46 Is Involved in OC-MQ-Induced Ovarian Cancer Cell Invasion via Epithelial-to-Mesenchymal Transition
3.3. Wnt/β-Catenin Signaling Functions as a Downstream Pathway of TRIM46
3.4. CXCL8 Derived from TAMs Promotes TRIM46 Expression and Ovarian Cancer Cell Invasion
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Duggan, M.A.; Anderson, W.F.; Altekruse, S.; Penberthy, L.; Sherman, M.E. The Surveillance, Epidemiology, and End Results (SEER) Program and Pathology: Toward Strengthening the Critical Relationship. Am. J. Surg. Pathol. 2016, 40, e94–e102. [Google Scholar] [CrossRef] [PubMed]
- Guan, X. Cancer metastases: Challenges and opportunities. Acta Pharm. Sin. B 2015, 5, 402–418. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, Y.; Yang, J.; Zhao, X.; Wei, X. Tumor Microenvironment in Ovarian Cancer: Function and Therapeutic Strategy. Front. Cell Dev. Biol. 2020, 8, 758. [Google Scholar] [CrossRef] [PubMed]
- Worzfeld, T.; Pogge von Strandmann, E.; Huber, M.; Adhikary, T.; Wagner, U.; Reinartz, S.; Müller, R. The unique molecular and cellular microenvironment of ovarian cancer. Front. Oncol. 2017, 7, 24. [Google Scholar] [CrossRef]
- Pan, Y.; Yu, Y.; Wang, X.; Zhang, T. Tumor-Associated Macrophages in Tumor Immunity. Front. Immunol. 2020, 11, 583084. [Google Scholar] [CrossRef] [PubMed]
- Schweer, D.; McAtee, A.; Neupane, K.; Richards, C.; Ueland, F.; Kolesar, J. Tumor-Associated Macrophages and Ovarian Cancer: Implications for Therapy. Cancers 2022, 14, 2220. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, W.; Zheng, L.; Guo, Q. The roles and targeting options of TRIM family proteins in tumor. Front. Pharmacol. 2022, 13, 999380. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Sun, X.; Li, P.; Liu, X.; Zhang, X.; Chen, Q.; Xin, H. TRIM family contribute to tumorigenesis, cancer development, and drug resistance. Exp. Hematol. Oncol. 2022, 11, 75. [Google Scholar] [CrossRef] [PubMed]
- Kapitein, L.C.; Hoogenraad, C.C. Building the neuronal microtubule cytoskeleton. Neuron 2015, 87, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, X.; Dong, W.; Sun, C.; Guo, D.; Zhang, L. Mmu-miR-1894-3p inhibits cell proliferation and migration of breast cancer cells by targeting Trim46. Int. J. Mol. Sci. 2016, 17, 609. [Google Scholar] [CrossRef]
- Cho, S.K.; Lee, K.; Woo, J.H.; Choi, J.H. Macrophages Promote Ovarian Cancer-Mesothelial Cell Adhesion by Upregulation of ITGA2 and VEGFC in Mesothelial Cells. Cells 2023, 12, 384. [Google Scholar] [CrossRef]
- Fogg, K.C.; Olson, W.R.; Miller, J.N.; Khan, A.; Renner, C.; Hale, I.; Weisman, P.S.; Kreeger, P.K. Alternatively activated macrophage-derived secretome stimulates ovarian cancer spheroid spreading through a JAK2/STAT3 pathway. Cancer Lett. 2019, 458, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.; Wang, Y.Y.; Choi, J.Y.; Lim, M.C.; Choi, J.H. CC Chemokine Ligand 7 Derived from Cancer-Stimulated Macrophages Promotes Ovarian Cancer Cell Invasion. Cancers 2021, 13, 2745. [Google Scholar] [CrossRef] [PubMed]
- Steitz, A.M.; Steffes, A.; Finkernagel, F.; Unger, A.; Sommerfeld, L.; Jansen, J.M.; Wagner, U.; Graumann, J.; Muller, R.; Reinartz, S. Tumor-associated macrophages promote ovarian cancer cell migration by secreting transforming growth factor beta induced (TGFBI) and tenascin C. Cell Death Dis. 2020, 11, 249. [Google Scholar] [CrossRef]
- Thompson, E.W.; Newgreen, D.F.; Tarin, D. Carcinoma invasion and metastasis: A role for epithelial-mesenchymal transition? Cancer Res. 2005, 65, 5991–5995; discussion 5995. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.; Duan, L.; Guo, Y.; Zhou, B.; Xu, Q.; Zhang, C.; Liu, W.; Liu, W.; Liu, Z.; Hu, J.; et al. TRIM46 upregulates Wnt/beta-catenin signaling by inhibiting Axin1 to mediate hypoxia-induced epithelial-mesenchymal transition in HK2 cells. Mol. Cell. Biochem. 2022, 477, 2829–2839. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, N.; Kurzrock, R. Targeting the Wnt/beta-catenin pathway in cancer: Update on effectors and inhibitors. Cancer Treat. Rev. 2018, 62, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Shen, J.; Yu, S.; Fei, J.; Zhu, X.; Zhao, J.; Zhai, L.; Sadhukhan, A.; Zhou, J. Tumor-Associated Macrophages (TAMs): A Critical Activator in Ovarian Cancer Metastasis. OncoTargets Ther. 2019, 12, 8687–8699. [Google Scholar] [CrossRef] [PubMed]
- Finkernagel, F.; Reinartz, S.; Lieber, S.; Adhikary, T.; Wortmann, A.; Hoffmann, N.; Bieringer, T.; Nist, A.; Stiewe, T.; Jansen, J.M. The transcriptional signature of human ovarian carcinoma macrophages is associated with extracellular matrix reorganization. Oncotarget 2016, 7, 75339. [Google Scholar] [CrossRef]
- Wen, J.; Zhao, Z.; Huang, L.; Wang, L.; Miao, Y.; Wu, J. IL-8 promotes cell migration through regulating EMT by activating the Wnt/beta-catenin pathway in ovarian cancer. J. Cell. Mol. Med. 2020, 24, 1588–1598. [Google Scholar] [CrossRef]
- Yang, W.; Liu, L.; Li, C.; Luo, N.; Chen, R.; Li, L.; Yu, F.; Cheng, Z. TRIM52 plays an oncogenic role in ovarian cancer associated with NF-kB pathway. Cell Death Dis. 2018, 9, 908. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, Z. TRIM37 promotes the aggressiveness of ovarian cancer cells and increases c-Myc expression by binding to HUWE1. Arch Biochem. Biophys. 2022, 728, 109372. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Fu, Y.; Xing, Y. TRIM47 promotes ovarian cancer cell proliferation, migration, and invasion by activating STAT3 signaling. Clinics 2022, 77, 100122. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Yu, C.; Chen, M.; Tian, S.; Sun, C. Over-expression of TRIM37 promotes cell migration and metastasis in hepatocellular carcinoma by activating Wnt/β-catenin signaling. Biochem. Biophys. Res. Commun. 2015, 464, 1120–1127. [Google Scholar] [CrossRef]
- Tian, H.; Zhao, H.; Qu, B.; Chu, X.; Xin, X.; Zhang, Q.; Li, W.; Yang, S. TRIM24 promotes colorectal cancer cell progression via the Wnt/beta-catenin signaling pathway activation. Am. J. Transl. Res. 2022, 14, 831–848. [Google Scholar]
- Li, K.; Pan, W.; Ma, Y.; Xu, X.; Gao, Y.; He, Y.; Wei, L.; Zhang, J. A novel oncogene TRIM63 promotes cell proliferation and migration via activating Wnt/beta-catenin signaling pathway in breast cancer. Pathol. Res. Pract. 2019, 215, 152573. [Google Scholar] [CrossRef]
- Xu, R.; Hu, J.; Zhang, T.; Jiang, C.; Wang, H.Y. TRIM29 overexpression is associated with poor prognosis and promotes tumor progression by activating Wnt/beta-catenin pathway in cervical cancer. Oncotarget 2016, 7, 28579–28591. [Google Scholar] [CrossRef]
- Tantai, J.; Pan, X.; Chen, Y.; Shen, Y.; Ji, C. TRIM46 activates AKT/HK2 signaling by modifying PHLPP2 ubiquitylation to promote glycolysis and chemoresistance of lung cancer cells. Cell Death Dis. 2022, 13, 285. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Cai, X.; Xu, T.; Liu, K.; Yang, D.; Fan, L.; Li, G.; Yu, X. Tripartite Motif-Containing 46 Promotes Viability and Inhibits Apoptosis of Osteosarcoma Cells by Activating NF-B Signaling Through Ubiquitination of PPAR. Oncol. Res. 2020, 28, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Gatcliffe, T.; Monk, B.; Planutis, K.; Holcombe, R. Wnt signaling in ovarian tumorigenesis. Int. J. Gynecol. Cancer 2008, 18, 954–962. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Inoue, S. TRIM proteins as RING finger E3 ubiquitin ligases. Adv. Exp. Med. Biol. 2012, 770, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Sampson, C.; Wang, Q.; Otkur, W.; Zhao, H.; Lu, Y.; Liu, X.; Piao, H.L. The roles of E3 ubiquitin ligases in cancer progression and targeted therapy. Clin. Transl. Med. 2023, 13, e1204. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Nie, F.; Gan, L.; Wei, X.; Liu, P.; Liu, H.; Zhang, K.; Fang, Z.; Wang, H.; Fang, N. Tripartite motif 31 drives gastric cancer cell proliferation and invasion through activating the Wnt/beta-catenin pathway by regulating Axin1 protein stability. Sci. Rep. 2023, 13, 20099. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Qian, H.; Xu, W.P.; Xiao, M.C.; Ding, C.H.; Liu, F.; Hong, H.Y.; Liu, S.Q.; Zhang, X.; Xie, W.F. Tripartite motif 8 promotes the progression of hepatocellular carcinoma via mediating ubiquitination of HNF1alpha. Cell Death Dis. 2024, 15, 416. [Google Scholar] [CrossRef]
- Ham, S.; Lima, L.G.; Lek, E.; Moller, A. The Impact of the Cancer Microenvironment on Macrophage Phenotypes. Front. Immunol. 2020, 11, 1308. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, R.C.; Zhang, X.L.; Niu, X.L.; Qu, Y.; Li, L.Z.; Meng, X.Y. Interleukin-8 secretion by ovarian cancer cells increases anchorage-independent growth, proliferation, angiogenic potential, adhesion and invasion. Cytokine 2012, 59, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.J.; Li, Y.B.; Yang, L.X.; Cheng, H.J.; Liu, X.; Chen, H. Roles of the CXCL8-CXCR1/2 Axis in the Tumor Microenvironment and Immunotherapy. Molecules 2021, 27, 137. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-Y.; Choi, M.-J.; Kim, J.-H.; Choi, J.-H. Enhanced Expression of TRIM46 in Ovarian Cancer Cells Induced by Tumor-Associated Macrophages Promotes Invasion via the Wnt/β-Catenin Pathway. Cells 2025, 14, 214. https://doi.org/10.3390/cells14030214
Wang Y-Y, Choi M-J, Kim J-H, Choi J-H. Enhanced Expression of TRIM46 in Ovarian Cancer Cells Induced by Tumor-Associated Macrophages Promotes Invasion via the Wnt/β-Catenin Pathway. Cells. 2025; 14(3):214. https://doi.org/10.3390/cells14030214
Chicago/Turabian StyleWang, Yi-Yue, Min-Jun Choi, Jin-Hyung Kim, and Jung-Hye Choi. 2025. "Enhanced Expression of TRIM46 in Ovarian Cancer Cells Induced by Tumor-Associated Macrophages Promotes Invasion via the Wnt/β-Catenin Pathway" Cells 14, no. 3: 214. https://doi.org/10.3390/cells14030214
APA StyleWang, Y.-Y., Choi, M.-J., Kim, J.-H., & Choi, J.-H. (2025). Enhanced Expression of TRIM46 in Ovarian Cancer Cells Induced by Tumor-Associated Macrophages Promotes Invasion via the Wnt/β-Catenin Pathway. Cells, 14(3), 214. https://doi.org/10.3390/cells14030214