Tissue-Specific Actions of Glucocorticoids on Apoptosis: A Double-Edged Sword

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction


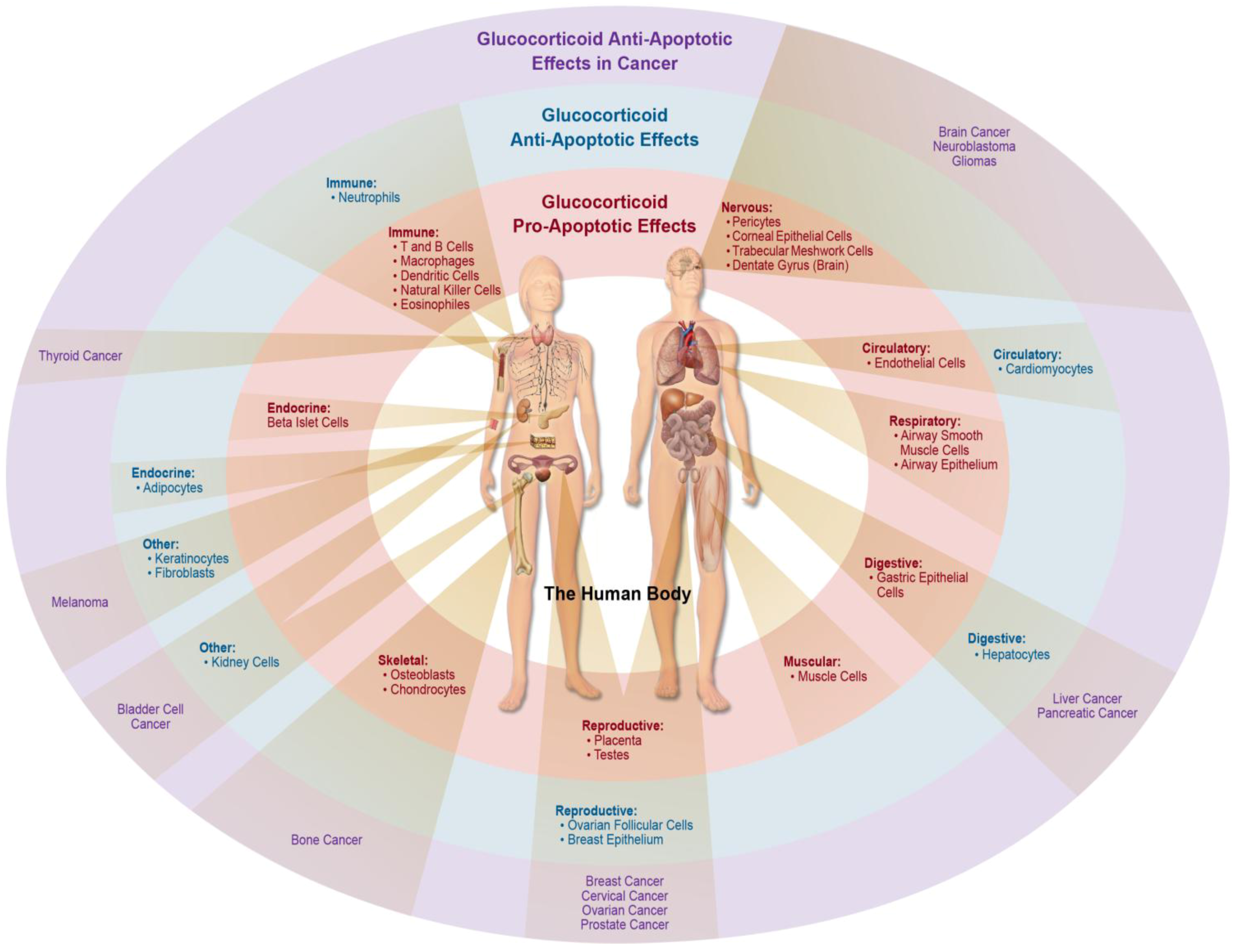
2. GC-Induced Apoptosis in Organ Systems
2.1. Skeletal System
2.2. Skeletal Muscular System
2.3. Respiratory System
2.4. Circulatory System
2.5. Nervous System
2.6. Digestive System
2.7. Endocrine System
2.8. Reproductive System
2.9. Immune System
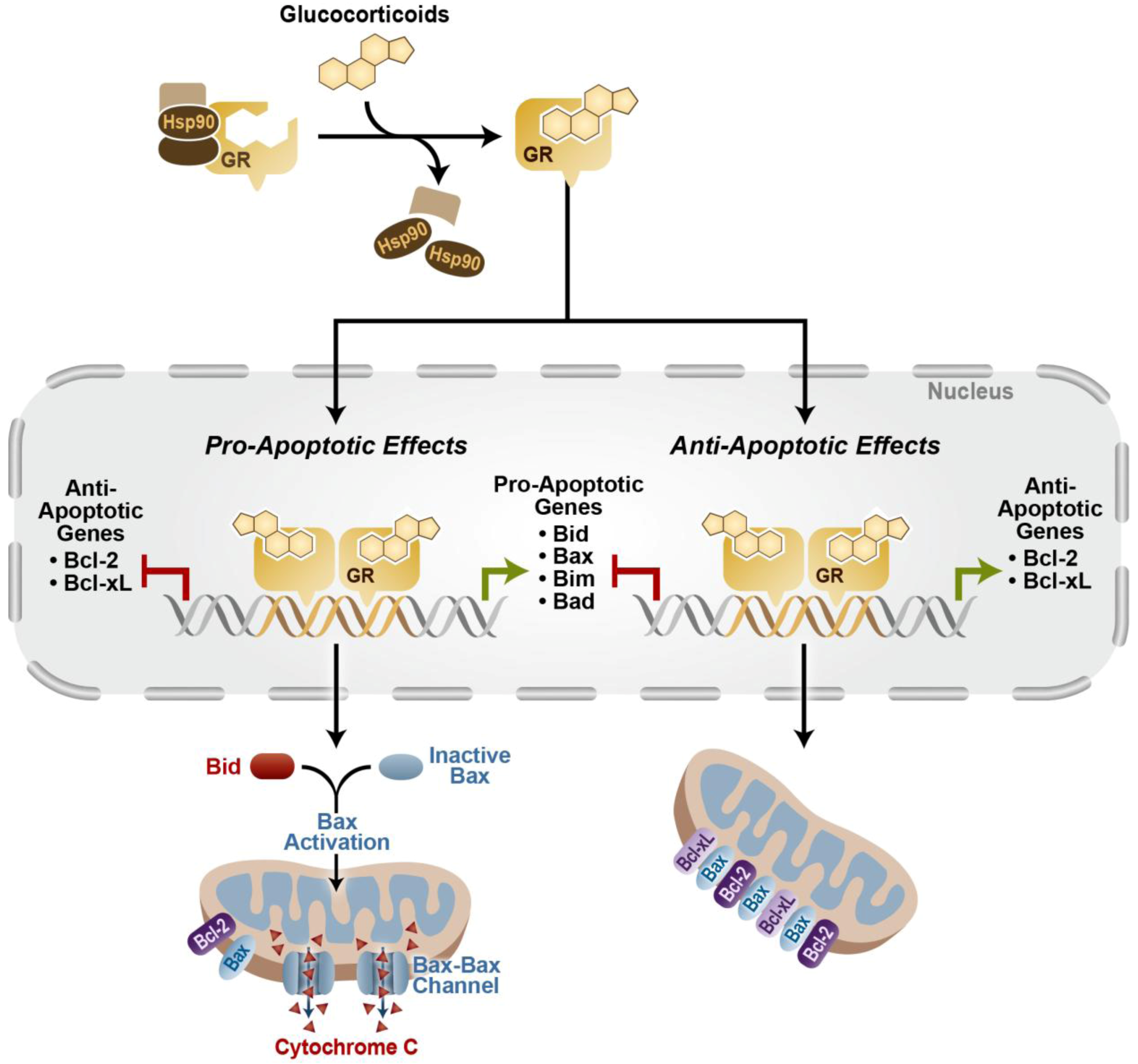
3. Anti-Apoptotic Effects of Glucocorticoid Signaling
3.1. Anti-Apoptotic Effects of Glucocorticoid Signaling in Normal Tissue
3.2. Resistance and Anti-Apoptotic Effects of Glucocorticoid Signaling in Cancer
4. Other Clinical Implications and Concluding Remarks

Acknowledgments
Conflict of Interest
References
- Cole, T.J.; Blendy, J.A.; Monaghan, A.P.; Krieglstein, K.; Schmid, W.; Aguzzi, A.; Fantuzzi, G.; Hummler, E.; Unsicker, K.; Schutz, G. Targeted disruption of the glucocorticoid receptor gene blocks adrenergic chromaffin cell development and severely retards lung maturation. Genes Dev. 1995, 9, 1608–1621. [Google Scholar] [CrossRef]
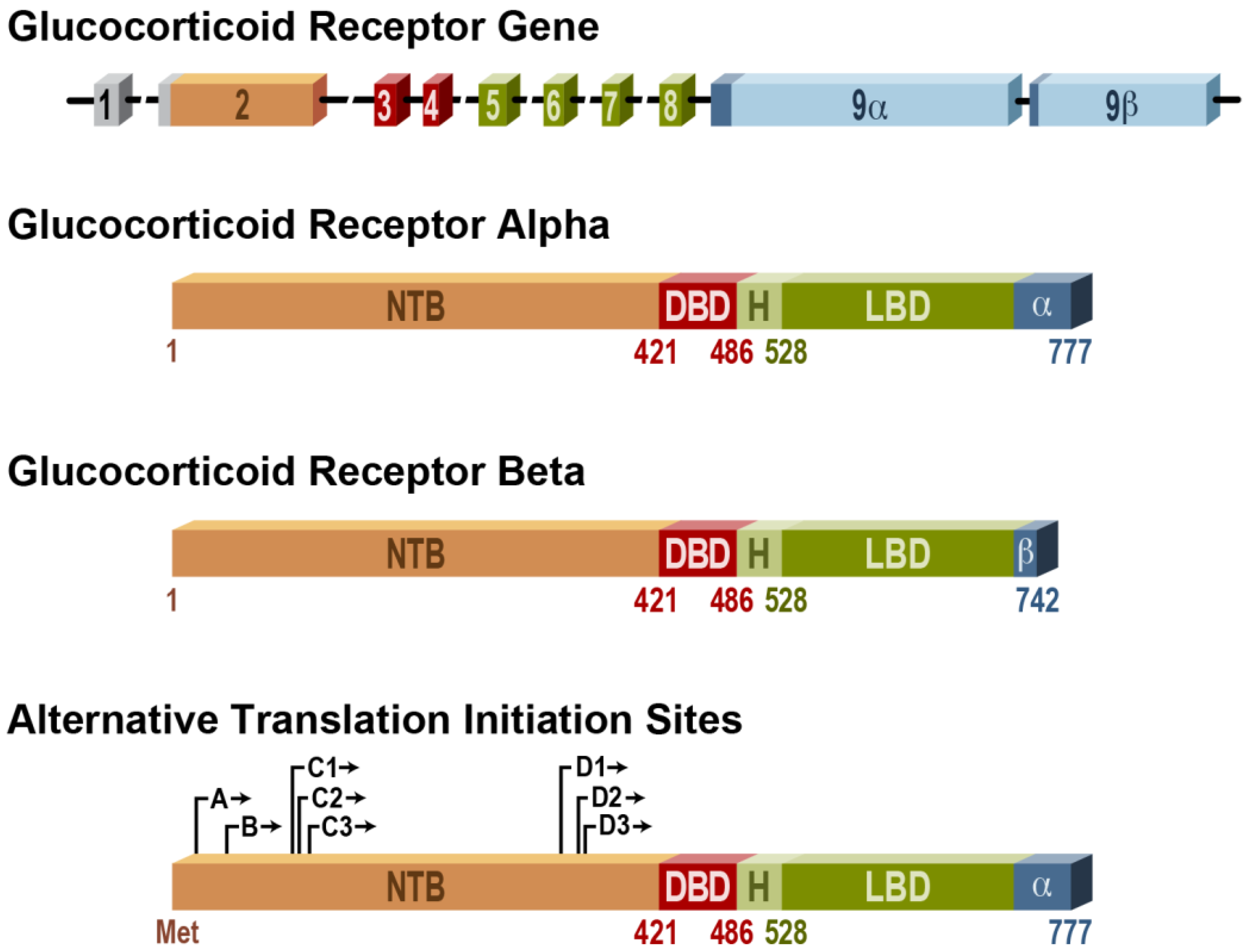
- Lu, N.Z.; Cidlowski, J.A. The origin and functions of multiple human glucocorticoid receptor isoforms. Ann. NY Acad. Sci. 2004, 1024, 102–123. [Google Scholar] [CrossRef]
- Oakley, R.H.; Sar, M.; Cidlowski, J.A. The human glucocorticoid receptor beta isoform. Expression, biochemical properties, and putative function. J. Biol. Chem. 1996, 271, 9550–9559. [Google Scholar]
- Smith, L.K.; Cidlowski, J.A. Glucocorticoid-induced apoptosis of healthy and malignant lymphocytes. Prog. Brain Res. 2010, 182, 1–30. [Google Scholar] [CrossRef]
- Lu, N.Z.; Collins, J.B.; Grissom, S.F.; Cidlowski, J.A. Selective regulation of bone cell apoptosis by translational isoforms of the glucocorticoid receptor. Mol. Cell Biol. 2007, 27, 7143–7160. [Google Scholar] [CrossRef]
- Mann, C.L.; Cidlowski, J.A. Glucocorticoids regulate plasma membrane potential during rat thymocyte apoptosis in vivo and in vitro. Endocrinology 2001, 142, 421–429. [Google Scholar] [CrossRef]
- Reichardt, H.M.; Kaestner, K.H.; Tuckermann, J.; Kretz, O.; Wessely, O.; Bock, R.; Gass, P.; Schmid, W.; Herrlich, P.; Angel, P.; et al. DNA binding of the glucocorticoid receptor is not essential for survival. Cell 1998, 93, 531–541. [Google Scholar] [CrossRef]
- Herr, I.; Gassler, N.; Friess, H.; Buchler, M.W. Regulation of differential pro- and anti-apoptotic signaling by glucocorticoids. Apoptosis 2007, 12, 271–291. [Google Scholar] [CrossRef]
- Saleh, A.; Srinivasula, S.M.; Acharya, S.; Fishel, R.; Alnemri, E.S. Cytochrome c and dATP-mediated oligomerization of Apaf-1 is a prerequisite for procaspase-9 activation. J. Biol. Chem. 1999, 274, 17941–17945. [Google Scholar]
- Moutsatsou, P.; Kassi, E.; Papavassiliou, A.G. Glucocorticoid receptor signaling in bone cells. Trends Mol. Med. 2012, 18, 348–359. [Google Scholar] [CrossRef]
- Li, H.; Qian, W.; Weng, X.; Wu, Z.; Zhuang, Q.; Feng, B.; Bian, Y. Glucocorticoid receptor and sequential P53 activation by dexamethasone mediates apoptosis and cell cycle arrest of osteoblastic MC3T3-E1 cells. PLoS One 2012, 7, e37030. [Google Scholar]
- Zaman, F.; Chrysis, D.; Huntjens, K.; Fadeel, B.; Savendahl, L. Ablation of the pro-apoptotic protein Bax protects mice from glucocorticoid-induced bone growth impairment. PLoS One 2012, 7, e33168. [Google Scholar]
- Smink, J.J.; Gresnigt, M.G.; Hamers, N.; Koedam, J.A.; Berger, R.; van Buul-Offers, S.C. Short-term glucocorticoid treatment of prepubertal mice decreases growth and IGF-I expression in the growth plate. J. Endocrinol. 2003, 177, 381–388. [Google Scholar] [CrossRef]
- Chrysis, D.; Zaman, F.; Chagin, A.S.; Takigawa, M.; Savendahl, L. Dexamethasone induces apoptosis in proliferative chondrocytes through activation of caspases and suppression of the Akt-phosphatidylinositol 3'-kinase signaling pathway. Endocrinology 2005, 146, 1391–1397. [Google Scholar]
- Dirks-Naylor, A.J.; Griffiths, C.L. Glucocorticoid-induced apoptosis and cellular mechanisms of myopathy. J. Steroid. Biochem. Mol. Biol. 2009, 117, 1–7. [Google Scholar] [CrossRef]
- Lofberg, E.; Gutierrez, A.; Wernerman, J.; Anderstam, B.; Mitch, W.E.; Price, S.R.; Bergstrom, J.; Alvestrand, A. Effects of high doses of glucocorticoids on free amino acids, ribosomes and protein turnover in human muscle. Eur. J. Clin. Invest. 2002, 32, 345–353. [Google Scholar] [CrossRef]
- Lee, M.C.; Wee, G.R.; Kim, J.H. Apoptosis of skeletal muscle on steroid-induced myopathy in rats. J. Nutr. 2005, 135, 1806S–1808S. [Google Scholar]
- Oshima, Y.; Kuroda, Y.; Kunishige, M.; Matsumoto, T.; Mitsui, T. Oxidative stress-associated mitochondrial dysfunction in corticosteroid-treated muscle cells. Muscle Nerve 2004, 30, 49–54. [Google Scholar] [CrossRef]
- Orzechowski, A.; Jank, M.; Gajkowska, B.; Sadkowski, T.; Godlewski, M.M.; Ostaszewski, P. Delineation of signalling pathway leading to antioxidant-dependent inhibition of dexamethasone-mediated muscle cell death. J. Muscle Res. Cell. Motil. 2003, 24, 33–53. [Google Scholar] [CrossRef]
- Bulfield, G.; Siller, W.G.; Wight, P.A.; Moore, K.J. X chromosome-linked muscular dystrophy (mdx) in the mouse. Proc. Natl. Acad. Sci. USA 1984, 81, 1189–1192. [Google Scholar] [CrossRef]
- Kojima, S.; Takagi, A.; Watanabe, T. Effect of prednisolone on apoptosis and cellular infiltration in mdx mouse muscle. Rinsho Shinkeigaku 1999, 39, 1109–1113. [Google Scholar]
- Ding, M.J.; Wang, L.X.; Dai, Y.R. Changes of airway smooth muscle cell apoptosis in asthmatic airway remodeling and the effect of dexamethasone in rats. Zhonghua Jie He He Hu Xi Za Zhi 2008, 31, 607–610. [Google Scholar]
- Dorscheid, D.R.; Wojcik, K.R.; Sun, S.; Marroquin, B.; White, S.R. Apoptosis of airway epithelial cells induced by corticosteroids. Am. J. Respir. Crit. Care Med. 2001, 164, 1939–1947. [Google Scholar]
- Vogt, C.J.; Schmid-Schonbein, G.W. Microvascular endothelial cell death and rarefaction in the glucocorticoid-induced hypertensive rat. Microcirculation 2001, 8, 129–139. [Google Scholar]
- Gaytan, F.; Morales, C.; Bellido, C.; Sanchez-Criado, J.E. Selective apoptosis of luteal endothelial cells in dexamethasone-treated rats leads to ischemic necrosis of luteal tissue. Biol. Reprod. 2002, 66, 232–240. [Google Scholar] [CrossRef]
- Newton, C.J.; Ran, G.; Xie, Y.X.; Bilko, D.; Burgoyne, C.H.; Adams, I.; Abidia, A.; McCollum, P.T.; Atkin, S.L. Statin-induced apoptosis of vascular endothelial cells is blocked by dexamethasone. J. Endocrinol. 2002, 174, 7–16. [Google Scholar] [CrossRef]
- Williams, T.A.; Verhovez, A.; Milan, A.; Veglio, F.; Mulatero, P. Protective effect of spironolactone on endothelial cell apoptosis. Endocrinology 2006, 147, 2496–2505. [Google Scholar] [CrossRef]
- Katychev, A.; Wang, X.; Duffy, A.; Dore-Duffy, P. Glucocorticoid-induced apoptosis in CNS microvascular pericytes. Dev. Neurosci. 2003, 25, 436–446. [Google Scholar] [CrossRef]
- Wordinger, R.J.; Clark, A.F. Effects of glucocorticoids on the trabecular meshwork: Towards a better understanding of glaucoma. Prog. Retin. Eye Res. 1999, 18, 629–667. [Google Scholar] [CrossRef]
- Gu, Y.; Zeng, S.; Qiu, P.; Peng, D.; Yan, G. Apoptosis of bovine trabecular meshwork cells induced by dexamethasone. Zhonghua Yan Ke Za Zhi 2002, 38, 302–304. [Google Scholar]
- Chen, W.L.; Lin, C.T.; Yao, C.C.; Huang, Y.H.; Chou, Y.B.; Yin, H.S.; Hu, F.R. In-vitro effects of dexamethasone on cellular proliferation, apoptosis, and Na+-K+-ATPase activity of bovine corneal endothelial cells. Ocul. Immunol. Inflamm. 2006, 14, 215–223. [Google Scholar] [CrossRef]
- Bourcier, T.; Forgez, P.; Borderie, V.; Scheer, S.; Rostene, W.; Laroche, L. Regulation of human corneal epithelial cell proliferation and apoptosis by dexamethasone. Invest. Ophthalmol. Vis. Sci. 2000, 41, 4133–4141. [Google Scholar]
- Hassan, A.H.; von Rosenstiel, P.; Patchev, V.K.; Holsboer, F.; Almeida, O.F. Exacerbation of apoptosis in the dentate gyrus of the aged rat by dexamethasone and the protective role of corticosterone. Exp. Neurol. 1996, 140, 43–52. [Google Scholar] [CrossRef]
- Brummelte, S.; Galea, L.A. Chronic high corticosterone reduces neurogenesis in the dentate gyrus of adult male and female rats. Neuroscience 2010, 168, 680–690. [Google Scholar] [CrossRef]
- Yu, S.; Holsboer, F.; Almeida, O.F. Neuronal actions of glucocorticoids: Focus on depression. J. Steroid Biochem. Mol. Biol. 2008, 108, 300–309. [Google Scholar] [CrossRef]
- Lu, J.; Goula, D.; Sousa, N.; Almeida, O.F. Ionotropic and metabotropic glutamate receptor mediation of glucocorticoid-induced apoptosis in hippocampal cells and the neuroprotective role of synaptic N-methyl-D-aspartate receptors. Neuroscience 2003, 121, 123–131. [Google Scholar] [CrossRef]
- Crochemore, C.; Lu, J.; Wu, Y.; Liposits, Z.; Sousa, N.; Holsboer, F.; Almeida, O.F. Direct targeting of hippocampal neurons for apoptosis by glucocorticoids is reversible by mineralocorticoid receptor activation. Mol. Psychiatry 2005, 10, 790–798. [Google Scholar] [CrossRef] [Green Version]
- Lupien, S.J.; McEwen, B.S.; Gunnar, M.R.; Heim, C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 2009, 10, 434–445. [Google Scholar] [CrossRef]
- Kempermann, G.; Krebs, J.; Fabel, K. The contribution of failing adult hippocampal neurogenesis to psychiatric disorders. Curr. Opin. Psychiatry 2008, 21, 290–295. [Google Scholar] [CrossRef]
- Sloviter, R.S.; Sollas, A.L.; Dean, E.; Neubort, S. Adrenalectomy-induced granule cell degeneration in the rat hippocampal dentate gyrus: Characterization of an in vivo model of controlled neuronal death. J. Comp. Neurol. 1993, 330, 324–336. [Google Scholar] [CrossRef]
- Yu, S.; Patchev, A.V.; Wu, Y.; Lu, J.; Holsboer, F.; Zhang, J.Z.; Sousa, N.; Almeida, O.F. Depletion of the neural precursor cell pool by glucocorticoids. Ann. Neurol. 2010, 67, 21–30. [Google Scholar] [CrossRef]
- Noguchi, K.K.; Walls, K.C.; Wozniak, D.F.; Olney, J.W.; Roth, K.A.; Farber, N.B. Acute neonatal glucocorticoid exposure produces selective and rapid cerebellar neural progenitor cell apoptotic death. Cell Death Differ. 2008, 15, 1582–1592. [Google Scholar] [CrossRef]
- Noguchi, K.K.; Lau, K.; Smith, D.J.; Swiney, B.S.; Farber, N.B. Glucocorticoid receptor stimulation and the regulation of neonatal cerebellar neural progenitor cell apoptosis. Neurobiol. Dis. 2011, 43, 356–363. [Google Scholar] [CrossRef]
- Roberts, D.; Dalziel, S. Antenatal corticosteroids for accelerating fetal lung maturation for women at risk of preterm birth. Cochrane Database Syst. Rev. 2006. [Google Scholar] [CrossRef] [Green Version]
- Tam, E.W.; Chau, V.; Ferriero, D.M.; Barkovich, A.J.; Poskitt, K.J.; Studholme, C.; Fok, E.D.; Grunau, R.E.; Glidden, D.V.; Miller, S.P. Preterm cerebellar growth impairment after postnatal exposure to glucocorticoids. Sci. Transl. Med. 2011, 3, 105ra105. [Google Scholar] [CrossRef]
- Tani, S.; Matsuda, K.; Tanaka, T. Induction of apoptosis in cultured rat gastric epithelial cells by ipriflavone: Comparison with indomethacin. Biol. Pharm. Bull. 2004, 27, 647–651. [Google Scholar] [CrossRef]
- Zhang, J.; Ke, Y.; Ning, T. Glucocorticoid-induced apoptosis of human gastric epithelial cells transfected with p53 genes. Zhonghua Zhong Liu Za Zhi 1996, 18, 328–330. [Google Scholar]
- Bandyopadhyay, U.; Biswas, K.; Bandyopadhyay, D.; Ganguly, C.K.; Banerjee, R.K. Dexamethasone makes the gastric mucosa susceptible to ulceration by inhibiting prostaglandin synthetase and peroxidase—Two important gastroprotective enzymes. Mol. Cell Biochem. 1999, 202, 31–36. [Google Scholar]
- Di Dalmazi, G.; Pagotto, U.; Pasquali, R.; Vicennati, V. Glucocorticoids and type 2 diabetes: From physiology to pathology. J. Nutr. Metab. 2012, 2012, 525093. [Google Scholar]
- Ranta, F.; Avram, D.; Berchtold, S.; Dufer, M.; Drews, G.; Lang, F.; Ullrich, S. Dexamethasone induces cell death in insulin-secreting cells, an effect reversed by exendin-4. Diabetes 2006, 55, 1380–1390. [Google Scholar] [CrossRef]
- Reich, E.; Tamary, A.; Sionov, R.V.; Melloul, D. Involvement of thioredoxin-interacting protein (TXNIP) in glucocorticoid-mediated beta cell death. Diabetologia 2012, 55, 1048–1057. [Google Scholar] [CrossRef]
- Roma, L.P.; Souza, K.L.; Carneiro, E.M.; Boschero, A.C.; Bosqueiro, J.R. Pancreatic islets from dexamethasone-treated rats show alterations in global gene expression and mitochondrial pathways. Gen. Physiol. Biophys. 2012, 31, 65–76. [Google Scholar] [CrossRef]
- Waddell, B.H.; Hisheh, S.; Dharmarajan, A.M.; Burton, P.J. Apoptosis in rat placenta is zone-dependent and stimulated by glucocorticoids. Biol. Reprod. 2000, 63, 1913–1917. [Google Scholar] [CrossRef]
- Poulain, M.; Frydman, N.; Duquenne, C.; N'Tumba-Byn, T.; Benachi, A.; Habert, R.; Rouiller-Fabre, V.; Livera, G. Dexamethasone induces germ cell apoptosis in the human fetal ovary. J. Clin. Endocrinol. Metab. 2012, 97, E1890–E1897. [Google Scholar] [CrossRef]
- Orazizadeh, M.; Khorsandi, L.S.; Hashemitabar, M. Toxic effects of dexamethasone on mouse testicular germ cells. Andrologia 2010, 42, 247–253. [Google Scholar] [CrossRef]
- Mahmoud, H.; Mahmoud, O.; Layasadat, K.; Naeim, A. Dexamethasone effects on Bax expression in the mouse testicular germ cells. Folia Histochem. Cytobiol. 2009, 47, 237–241. [Google Scholar]
- Yazawa, H.; Sasagawa, I.; Nakada, T. Apoptosis of testicular germ cells induced by exogenous glucocorticoid in rats. Hum. Reprod. 2000, 15, 1917–1920. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Q.; Wang, F.F.; Gao, H.B.; Zhang, P. Stress induces glucocorticoid-mediated apoptosis of rat Leydig cells in vivo. Stress 2012, 15, 74–84. [Google Scholar]
- Gao, H.B.; Tong, M.H.; Hu, Y.Q.; You, H.Y.; Guo, Q.S.; Ge, R.S.; Hardy, M.P. Mechanisms of glucocorticoid-induced Leydig cell apoptosis. Mol. Cell. Endocrinol. 2003, 199, 153–163. [Google Scholar] [CrossRef]
- Andric, S.A.; Kojic, Z.; Bjelic, M.M.; Mihajlovic, A.I.; Baburski, A.Z.; Sokanovic, S.J.; Janjic, M.M.; Stojkov, N.J.; Stojilkovic, S.S.; Kostic, T.S. The opposite roles of glucocorticoid and alpha1-adrenergic receptors in stress triggered apoptosis of rat Leydig cells. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E51–E59. [Google Scholar] [CrossRef]
- Zen, M.; Canova, M.; Campana, C.; Bettio, S.; Nalotto, L.; Rampudda, M.; Ramonda, R.; Iaccarino, L.; Doria, A. The kaleidoscope of glucorticoid effects on immune system. Autoimmun. Rev. 2011, 10, 305–310. [Google Scholar] [CrossRef]
- Cidlowski, J.A.; King, K.L.; Evans-Storms, R.B.; Montague, J.W.; Bortner, C.D.; Hughes, F.M., Jr. The biochemistry and molecular biology of glucocorticoid-induced apoptosis in the immune system. Recent. Prog. Horm. Res. 1996, 51, discussion 490–491. [Google Scholar]
- Montague, J.W.; Cidlowski, J.A. Glucocorticoid-induced death of immune cells: Mechanisms of action. Curr. Top. Microbiol. Immunol. 1995, 200, 51–65. [Google Scholar] [CrossRef]
- Ashwell, J.D.; Lu, F.W.; Vacchio, M.S. Glucocorticoids in T cell development and function*. Annu. Rev. Immunol. 2000, 18, 309–345. [Google Scholar] [CrossRef]
- Vacchio, M.S.; Ashwell, J.D. Glucocorticoids and thymocyte development. Semin. Immunol. 2000, 12, 475–485. [Google Scholar]
- Caron-Leslie, L.M.; Schwartzman, R.A.; Gaido, M.L.; Compton, M.M.; Cidlowski, J.A. Identification and characterization of glucocorticoid-regulated nuclease(s) in lymphoid cells undergoing apoptosis. J. Steroid Biochem. Mol. Biol. 1991, 40, 661–671. [Google Scholar] [CrossRef]
- Cohen, J.J. Glucocorticoid-induced apoptosis in the thymus. Semin. Immunol. 1992, 4, 363–369. [Google Scholar]
- Brunetti, M.; Martelli, N.; Colasante, A.; Piantelli, M.; Musiani, P.; Aiello, F.B. Spontaneous and glucocorticoid-induced apoptosis in human mature T lymphocytes. Blood 1995, 86, 4199–4205. [Google Scholar]
- Lill-Elghanian, D.; Schwartz, K.; King, L.; Fraker, P. Glucocorticoid-induced apoptosis in early B cells from human bone marrow. Exp. Biol. Med. (Maywood) 2002, 227, 763–770. [Google Scholar]
- Zhou, J.Y.; Zhong, H.J.; Yang, C.; Yan, J.; Wang, H.Y.; Jiang, J.X. Corticosterone exerts immunostimulatory effects on macrophages via endoplasmic reticulum stress. Br. J. Surg. 2010, 97, 281–293. [Google Scholar] [CrossRef]
- Cao, Y.; Bender, I.K.; Konstantinidis, A.K.; Shin, S.C.; Jewell, C.M.; Cidlowski, J.A.; Schleimer, R.P.; Lu, N.Z. Glucocorticoid receptor translational isoforms underlie maturational stage-specific glucocorticoid sensitivities of dendritic cells in mice and humans. Blood 2013, 121, 1553–1562. [Google Scholar] [CrossRef]
- Meagher, L.C.; Cousin, J.M.; Seckl, J.R.; Haslett, C. Opposing effects of glucocorticoids on the rate of apoptosis in neutrophilic and eosinophilic granulocytes. J. Immunol. 1996, 156, 4422–4428. [Google Scholar]
- Migliorati, G.; Pagliacci, C.; Moraca, R.; Crocicchio, F.; Nicoletti, I.; Riccardi, C. Glucocorticoid-induced apoptosis of natural killer cells and cytotoxic T lymphocytes. Pharmacol. Res. 1992, 26, 26–27. [Google Scholar]
- Saffar, A.S.; Ashdown, H.; Gounni, A.S. The molecular mechanisms of glucocorticoids-mediated neutrophil survival. Curr. Drug Targets 2011, 12, 556–562. [Google Scholar] [CrossRef]
- Woodward, M.J.; de Boer, J.; Heidorn, S.; Hubank, M.; Kioussis, D.; Williams, O.; Brady, H.J. Tnfaip8 is an essential gene for the regulation of glucocorticoid-mediated apoptosis of thymocytes. Cell Death Differ. 2010, 17, 316–323. [Google Scholar] [CrossRef]
- Kumar, D.; Whiteside, T.L.; Kasid, U. Identification of a novel tumor necrosis factor-alpha-inducible gene, SCC-S2, containing the consensus sequence of a death effector domain of fas-associated death domain-like interleukin-1beta-converting enzyme-inhibitory protein. J. Biol. Chem. 2000, 275, 2973–2978. [Google Scholar] [CrossRef]
- Smith, L.K.; Shah, R.R.; Cidlowski, J.A. Glucocorticoids modulate microRNA expression and processing during lymphocyte apoptosis. J. Biol. Chem. 2010, 285, 36698–36708. [Google Scholar] [CrossRef]
- Talaber, G.; Boldizsar, F.; Bartis, D.; Palinkas, L.; Szabo, M.; Berta, G.; Setalo, G., Jr.; Nemeth, P.; Berki, T. Mitochondrial translocation of the glucocorticoid receptor in double-positive thymocytes correlates with their sensitivity to glucocorticoid-induced apoptosis. Int. Immunol. 2009, 21, 1269–1276. [Google Scholar] [CrossRef]
- Sasson, R.; Shinder, V.; Dantes, A.; Land, A.; Amsterdam, A. Activation of multiple signal transduction pathways by glucocorticoids: Protection of ovarian follicular cells against apoptosis. Biochem. Biophys. Res. Commun. 2003, 311, 1047–1056. [Google Scholar] [CrossRef]
- Sasson, R.; Tajima, K.; Amsterdam, A. Glucocorticoids protect against apoptosis induced by serum deprivation, cyclic adenosine 3',5'-monophosphate and p53 activation in immortalized human granulosa cells: Involvement of Bcl-2. Endocrinology 2001, 142, 802–811. [Google Scholar] [CrossRef]
- Feng, Z.; Marti, A.; Jehn, B.; Altermatt, H.J.; Chicaiza, G.; Jaggi, R. Glucocorticoid and progesterone inhibit involution and programmed cell death in the mouse mammary gland. J. Cell Biol. 1995, 131, 1095–1103. [Google Scholar] [CrossRef]
- Stojadinovic, O.; Lee, B.; Vouthounis, C.; Vukelic, S.; Pastar, I.; Blumenberg, M.; Brem, H.; Tomic-Canic, M. Novel genomic effects of glucocorticoids in epidermal keratinocytes: Inhibition of apoptosis, interferon-gamma pathway, and wound healing along with promotion of terminal differentiation. J. Biol. Chem. 2007, 282, 4021–4034. [Google Scholar]
- Wen, L.P.; Madani, K.; Fahrni, J.A.; Duncan, S.R.; Rosen, G.D. Dexamethasone inhibits lung epithelial cell apoptosis induced by IFN-gamma and Fas. Am. J. Physiol. 1997, 273, L921–L929. [Google Scholar]
- Bailly-Maitre, B.; de Sousa, G.; Zucchini, N.; Gugenheim, J.; Boulukos, K.E.; Rahmani, R. Spontaneous apoptosis in primary cultures of human and rat hepatocytes: Molecular mechanisms and regulation by dexamethasone. Cell Death Differ. 2002, 9, 945–955. [Google Scholar]
- Oh, H.Y.; Namkoong, S.; Lee, S.J.; Por, E.; Kim, C.K.; Billiar, T.R.; Han, J.A.; Ha, K.S.; Chung, H.T.; Kwon, Y.G.; et al. Dexamethasone protects primary cultured hepatocytes from death receptor-mediated apoptosis by upregulation of cFLIP. Cell Death Differ. 2006, 13, 512–523. [Google Scholar] [CrossRef]
- Zhang, H.H.; Kumar, S.; Barnett, A.H.; Eggo, M.C. Dexamethasone inhibits tumor necrosis factor-alpha-induced apoptosis and interleukin-1 beta release in human subcutaneous adipocytes and preadipocytes. J. Clin. Endocrinol. Metab. 2001, 86, 2817–2825. [Google Scholar] [CrossRef]
- Xu, B.; Strom, J.; Chen, Q.M. Dexamethasone induces transcriptional activation of Bcl-xL gene and inhibits cardiac injury by myocardial ischemia. Eur. J. Pharmacol. 2011, 668, 194–200. [Google Scholar] [CrossRef]
- Pearl, J.M.; Nelson, D.P.; Schwartz, S.M.; Wagner, C.J.; Bauer, S.M.; Setser, E.A.; Duffy, J.Y. Glucocorticoids reduce ischemia-reperfusion-induced myocardial apoptosis in immature hearts. Ann. Thorac. Surg. 2002, 74, 830–837. [Google Scholar]
- Tokudome, S.; Sano, M.; Shinmura, K.; Matsuhashi, T.; Morizane, S.; Moriyama, H.; Tamaki, K.; Hayashida, K.; Nakanishi, H.; Yoshikawa, N.; et al. Glucocorticoid protects rodent hearts from ischemia/reperfusion injury by activating lipocalin-type prostaglandin D synthase-derived PGD2 biosynthesis. J. Clin. Invest. 2009, 119, 1477–1488. [Google Scholar] [CrossRef]
- Ren, R.; Oakley, R.H.; Cruz-Topete, D.; Cidlowski, J.A. Dual role for glucocorticoids in cardiomyocyte hypertrophy and apoptosis. Endocrinology 2012, 153, 5346–5360. [Google Scholar] [CrossRef]
- Chen, Q.M.; Alexander, D.; Sun, H.; Xie, L.; Lin, Y.; Terrand, J.; Morrissy, S.; Purdom, S. Corticosteroids inhibit cell death induced by doxorubicin in cardiomyocytes: Induction of antiapoptosis, antioxidant, and detoxification genes. Mol. Pharmacol. 2005, 67, 1861–1873. [Google Scholar] [CrossRef]
- Kewalramani, G.; Puthanveetil, P.; Wang, F.; Kim, M.S.; Deppe, S.; Abrahani, A.; Luciani, D.S.; Johnson, J.D.; Rodrigues, B. AMP-activated protein kinase confers protection against TNF-{alpha}-induced cardiac cell death. Cardiovasc. Res. 2009, 84, 42–53. [Google Scholar] [CrossRef]
- Forster, A.; Emmler, T.; Schwalm, S.; Ebadi, M.; Heringdorf, D.M.; Nieuwenhuis, B.; Kleuser, B.; Huwiler, A.; Pfeilschifter, J. Glucocorticoids protect renal mesangial cells from apoptosis by increasing cellular sphingosine-1-phosphate. Kidney Int. 2010, 77, 870–879. [Google Scholar] [CrossRef]
- Nieuwenhuis, B.; Luth, A.; Kleuser, B. Dexamethasone protects human fibroblasts from apoptosis via an S1P3-receptor subtype dependent activation of PKB/Akt and Bcl XL. Pharmacol. Res. 2010, 61, 449–459. [Google Scholar] [CrossRef]
- Wada, T.; Pippin, J.W.; Marshall, C.B.; Griffin, S.V.; Shankland, S.J. Dexamethasone prevents podocyte apoptosis induced by puromycin aminonucleoside: Role of p53 and Bcl-2-related family proteins. J. Am. Soc. Nephrol. 2005, 16, 2615–2625. [Google Scholar] [CrossRef]
- Messmer, U.K.; Winkel, G.; Briner, V.A.; Pfeilschifter, J. Suppression of apoptosis by glucocorticoids in glomerular endothelial cells: Effects on proapoptotic pathways. Br. J. Pharmacol. 2000, 129, 1673–1683. [Google Scholar] [CrossRef]
- Kumar, S.; Allen, D.A.; Kieswich, J.E.; Patel, N.S.; Harwood, S.; Mazzon, E.; Cuzzocrea, S.; Raftery, M.J.; Thiemermann, C.; Yaqoob, M.M. Dexamethasone ameliorates renal ischemia-reperfusion injury. J. Am. Soc. Nephrol. 2009, 20, 2412–2425. [Google Scholar] [CrossRef]
- Sionov, R.V.; Spokoini, R.; Kfir-Erenfeld, S.; Cohen, O.; Yefenof, E. Mechanisms regulating the susceptibility of hematopoietic malignancies to glucocorticoid-induced apoptosis. Adv. Cancer. Res. 2008, 101, 127–248. [Google Scholar] [CrossRef]
- Schmidt, S.; Rainer, J.; Ploner, C.; Presul, E.; Riml, S.; Kofler, R. Glucocorticoid-induced apoptosis and glucocorticoid resistance: Molecular mechanisms and clinical relevance. Cell Death Differ. 2004, 11, S45–S55. [Google Scholar] [CrossRef]
- Schlossmacher, G.; Stevens, A.; White, A. Glucocorticoid receptor-mediated apoptosis: Mechanisms of resistance in cancer cells. J. Endocrinol. 2011, 211, 17–25. [Google Scholar] [CrossRef]
- Gross, K.L.; Oakley, R.H.; Scoltock, A.B.; Jewell, C.M.; Cidlowski, J.A. Glucocorticoid receptor alpha isoform-selective regulation of antiapoptotic genes in osteosarcoma cells: A new mechanism for glucocorticoid resistance. Mol. Endocrinol. 2011, 25, 1087–1099. [Google Scholar] [CrossRef]
- Aapro, M.S. Present role of corticosteroids as antiemetics. Recent Results Cancer Res. 1991, 121, 91–100. [Google Scholar] [CrossRef]
- Kirkbride, P.; Bezjak, A.; Pater, J.; Zee, B.; Palmer, M.J.; Wong, R.; Cross, P.; Gulavita, S.; Blood, P.; Sun, A.; et al. Dexamethasone for the prophylaxis of radiation-induced emesis: A National Cancer Institute of Canada Clinical Trials Group phase III study. J. Clin. Oncol. 2000, 18, 1960–1966. [Google Scholar]
- Zhang, C.; Beckermann, B.; Kallifatidis, G.; Liu, Z.; Rittgen, W.; Edler, L.; Buchler, P.; Debatin, K.M.; Buchler, M.W.; Friess, H.; et al. Corticosteroids induce chemotherapy resistance in the majority of tumour cells from bone, brain, breast, cervix, melanoma and neuroblastoma. Int. J. Oncol. 2006, 29, 1295–1301. [Google Scholar]
- Zhang, C.; Mattern, J.; Haferkamp, A.; Pfitzenmaier, J.; Hohenfellner, M.; Rittgen, W.; Edler, L.; Debatin, K.M.; Groene, E.; Herr, I. Corticosteroid-induced chemotherapy resistance in urological cancers. Cancer Biol. Ther. 2006, 5, 59–64. [Google Scholar] [CrossRef]
- Zheng, Y.; Izumi, K.; Li, Y.; Ishiguro, H.; Miyamoto, H. Contrary regulation of bladder cancer cell proliferation and invasion by dexamethasone-mediated glucocorticoid receptor signals. Mol. Cancer Ther. 2012, 11, 2621–2632. [Google Scholar] [CrossRef]
- Ni Chonghaile, T.; Concannon, C.G.; Szegezdi, E.; Gorman, A.M.; Samali, A. Dexamethasone inhibits apoptosis in C6 glioma cells through increased expression of Bcl-XL. Apoptosis 2006, 11, 1247–1255. [Google Scholar] [CrossRef]
- Chang, T.C.; Hung, M.W.; Jiang, S.Y.; Chu, J.T.; Chu, L.L.; Tsai, L.C. Dexamethasone suppresses apoptosis in a human gastric cancer cell line through modulation of bcl-x gene expression. FEBS Lett. 1997, 415, 11–15. [Google Scholar] [CrossRef]
- Zhang, C.; Kolb, A.; Buchler, P.; Cato, A.C.; Mattern, J.; Rittgen, W.; Edler, L.; Debatin, K.M.; Buchler, M.W.; Friess, H.; et al. Corticosteroid co-treatment induces resistance to chemotherapy in surgical resections, xenografts and established cell lines of pancreatic cancer. BMC Cancer 2006, 6, 61. [Google Scholar] [CrossRef]
- Runnebaum, I.B.; Bruning, A. Glucocorticoids inhibit cell death in ovarian cancer and up-regulate caspase inhibitor cIAP2. Clin. Cancer Res. 2005, 11, 6325–6332. [Google Scholar] [CrossRef]
- Mitsiades, N.; Poulaki, V.; Tseleni-Balafouta, S.; Koutras, D.A.; Stamenkovic, I. Thyroid carcinoma cells are resistant to FAS-mediated apoptosis but sensitive to tumor necrosis factor-related apoptosis-inducing ligand. Cancer Res. 2000, 60, 4122–4129. [Google Scholar]
- Petrella, A.; Ercolino, S.F.; Festa, M.; Gentilella, A.; Tosco, A.; Conzen, S.D.; Parente, L. Dexamethasone inhibits TRAIL-induced apoptosis of thyroid cancer cells via Bcl-xL induction. Eur. J. Cancer. 2006, 42, 3287–3293. [Google Scholar] [CrossRef]
- Evans-Storms, R.B.; Cidlowski, J.A. Delineation of an antiapoptotic action of glucocorticoids in hepatoma cells: The role of nuclear factor-kappaB. Endocrinology 2000, 141, 1854–1862. [Google Scholar] [CrossRef]
- Scoltock, A.B.; Heimlich, G.; Cidlowski, J.A. Glucocorticoids inhibit the apoptotic actions of UV-C but not Fas ligand in hepatoma cells: Direct evidence for a critical role of Bcl-xL. Cell Death Differ. 2007, 14, 840–850. [Google Scholar] [CrossRef]
- Wanke, I.; Schwarz, M.; Buchmann, A. Insulin and dexamethasone inhibit TGF-beta-induced apoptosis of hepatoma cells upstream of the caspase activation cascade. Toxicology 2004, 204, 141–154. [Google Scholar] [CrossRef]
- Uen, Y.H.; Ko, P.H.; Yin, P.H.; Liu, T.Y.; Chi, C.W.; Lui, W.Y. Glucocorticoid protects hepatoma cells against metabolic stress-induced cell death. Int. J. Oncol. 2008, 33, 1263–1270. [Google Scholar]
- Yang, N.; Zhang, H.; Si-Ma, H.; Fu, Y.; Zhao, W.; Li, D.; Yang, G. Dexamethasone decreases hepatocellular carcinoma cell sensitivity to cisplatin-induced apoptosis. Hepatogastroenterology 2011, 58, 1730–1735. [Google Scholar]
- Pang, D.; Kocherginsky, M.; Krausz, T.; Kim, S.Y.; Conzen, S.D. Dexamethasone decreases xenograft response to Paclitaxel through inhibition of tumor cell apoptosis. Cancer Biol. Ther. 2006, 5, 933–940. [Google Scholar] [CrossRef]
- Wu, W.; Chaudhuri, S.; Brickley, D.R.; Pang, D.; Karrison, T.; Conzen, S.D. Microarray analysis reveals glucocorticoid-regulated survival genes that are associated with inhibition of apoptosis in breast epithelial cells. Cancer Res. 2004, 64, 1757–1764. [Google Scholar] [CrossRef]
- Beck, I.M.; de Bosscher, K.; Haegeman, G. Glucocorticoid receptor mutants: Man-made tools for functional research. Trends Endocrinol. Metab. 2011, 22, 295–310. [Google Scholar] [CrossRef]
- Reber, L.L.; Daubeuf, F.; Plantinga, M.; de Cauwer, L.; Gerlo, S.; Waelput, W.; van Calenbergh, S.; Tavernier, J.; Haegeman, G.; Lambrecht, B.N.; et al. A dissociated glucocorticoid receptor modulator reduces airway hyperresponsiveness and inflammation in a mouse model of asthma. J. Immunol. 2012, 188, 3478–3487. [Google Scholar] [CrossRef]
- Dewint, P.; Gossye, V.; de Bosscher, K.; Vanden Berghe, W.; van Beneden, K.; Deforce, D.; van Calenbergh, S.; Muller-Ladner, U.; Vander Cruyssen, B.; Verbruggen, G.; et al. A plant-derived ligand favoring monomeric glucocorticoid receptor conformation with impaired transactivation potential attenuates collagen-induced arthritis. J. Immunol. 2008, 180, 2608–2615. [Google Scholar]
- Van Loo, G.; Sze, M.; Bougarne, N.; Praet, J.; Mc Guire, C.; Ullrich, A.; Haegeman, G.; Prinz, M.; Beyaert, R.; de Bosscher, K. Antiinflammatory properties of a plant-derived nonsteroidal, dissociated glucocorticoid receptor modulator in experimental autoimmune encephalomyelitis. Mol. Endocrinol. 2010, 24, 310–322. [Google Scholar] [CrossRef]
- Liberman, A.C.; Antunica-Noguerol, M.; Ferraz-de-Paula, V.; Palermo-Neto, J.; Castro, C.N.; Druker, J.; Holsboer, F.; Perone, M.J.; Gerlo, S.; de Bosscher, K.; et al. Compound A, a dissociated glucocorticoid receptor modulator, inhibits T-bet (Th1) and induces GATA-3 (Th2) activity in immune cells. PLoS One 2012, 7, e35155. [Google Scholar] [CrossRef] [Green Version]
- Schacke, H.; Schottelius, A.; Docke, W.D.; Strehlke, P.; Jaroch, S.; Schmees, N.; Rehwinkel, H.; Hennekes, H.; Asadullah, K. Dissociation of transactivation from transrepression by a selective glucocorticoid receptor agonist leads to separation of therapeutic effects from side effects. Proc. Natl. Acad. Sci. USA 2004, 101, 227–232. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Cavet, M.E.; VanderMeid, K.R.; Salvador-Silva, M.; Lopez, F.J.; Ward, K.W. BOL-303242-X, a novel selective glucocorticoid receptor agonist, with full anti-inflammatory properties in human ocular cells. Mol. Vis. 2009, 15, 2606–2616. [Google Scholar]
- Cavet, M.E.; Harrington, K.L.; Ward, K.W.; Zhang, J.Z. Mapracorat, a novel selective glucocorticoid receptor agonist, inhibits hyperosmolar-induced cytokine release and MAPK pathways in human corneal epithelial cells. Mol. Vis. 2010, 16, 1791–1800. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gruver-Yates, A.L.; Cidlowski, J.A. Tissue-Specific Actions of Glucocorticoids on Apoptosis: A Double-Edged Sword. Cells 2013, 2, 202-223. https://doi.org/10.3390/cells2020202
Gruver-Yates AL, Cidlowski JA. Tissue-Specific Actions of Glucocorticoids on Apoptosis: A Double-Edged Sword. Cells. 2013; 2(2):202-223. https://doi.org/10.3390/cells2020202
Chicago/Turabian StyleGruver-Yates, Amanda L., and John A. Cidlowski. 2013. "Tissue-Specific Actions of Glucocorticoids on Apoptosis: A Double-Edged Sword" Cells 2, no. 2: 202-223. https://doi.org/10.3390/cells2020202
APA StyleGruver-Yates, A. L., & Cidlowski, J. A. (2013). Tissue-Specific Actions of Glucocorticoids on Apoptosis: A Double-Edged Sword. Cells, 2(2), 202-223. https://doi.org/10.3390/cells2020202