Development of a Novel BAFF Responsive Cell Line Suitable for Detecting Bioactive BAFF and Neutralizing Antibodies against BAFF-Pathway Inhibiting Therapeutics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Reagents
2.2. Cell Culture
2.3. Construction of BAFF-TNFR Hybrid Receptor Expression Plasmids
2.4. Transient Transfection
2.5. Establishment of BAFF Responsive Stable Cell Line
2.6. Flow Cytometry Analysis
2.7. Bioassays for BAFF, TNF and Other Cytokines
3. Results and Discussion
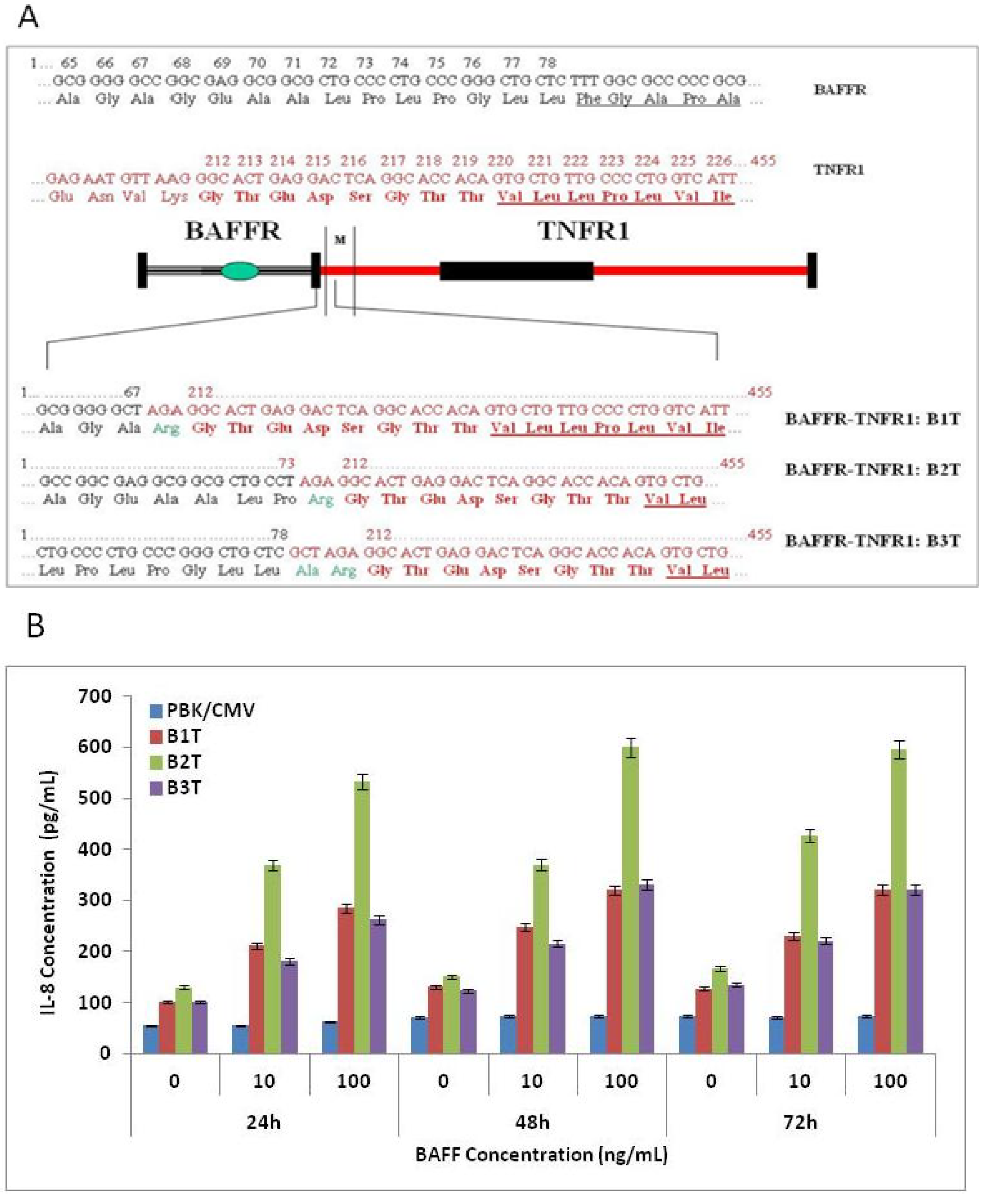
3.1. Construction of BAFF Receptor and TNF Receptor (BAFFR-TNFR) Hybrid Receptors

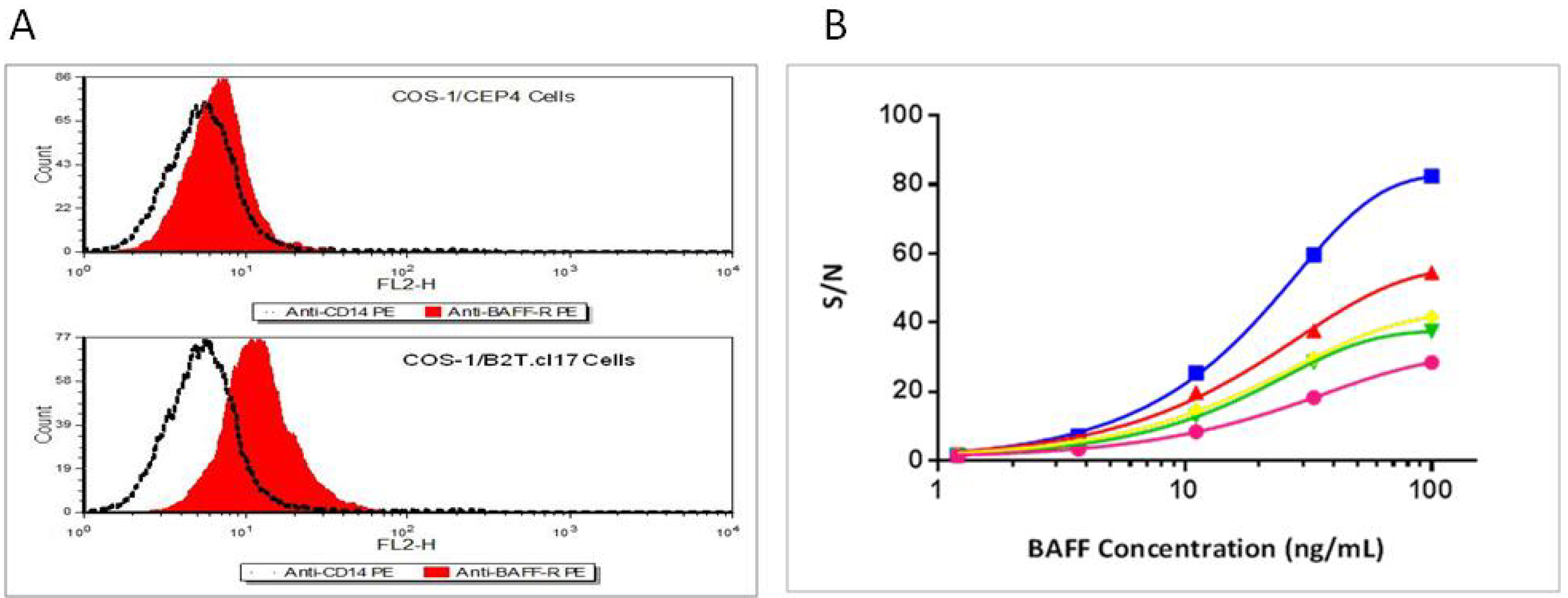
3.2. Hybrid Receptor is Expressed and Located on the Cell Surface of Stably Transfected COS-1 Cells

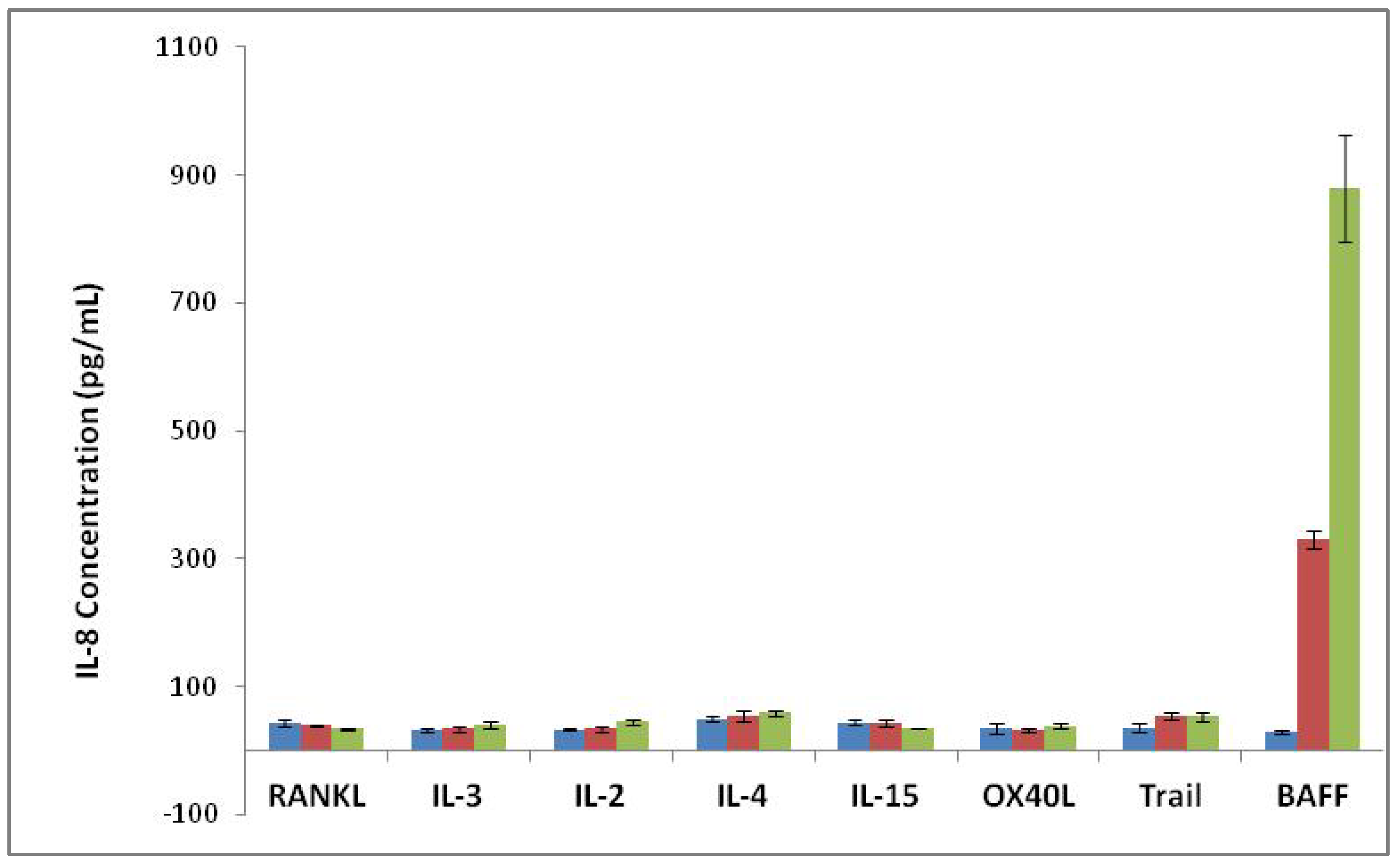
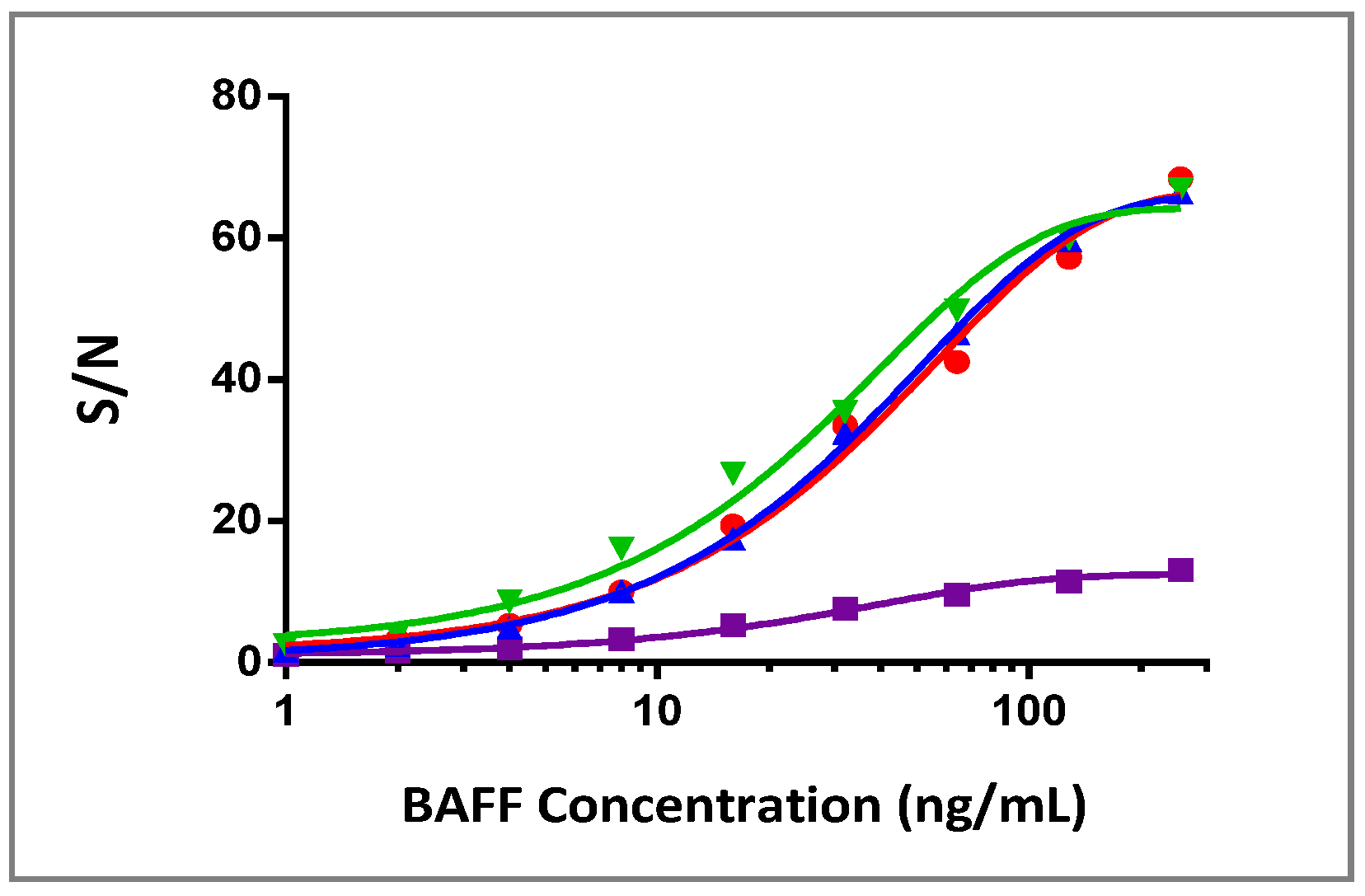
3.3. Specificity of BAFF Assay


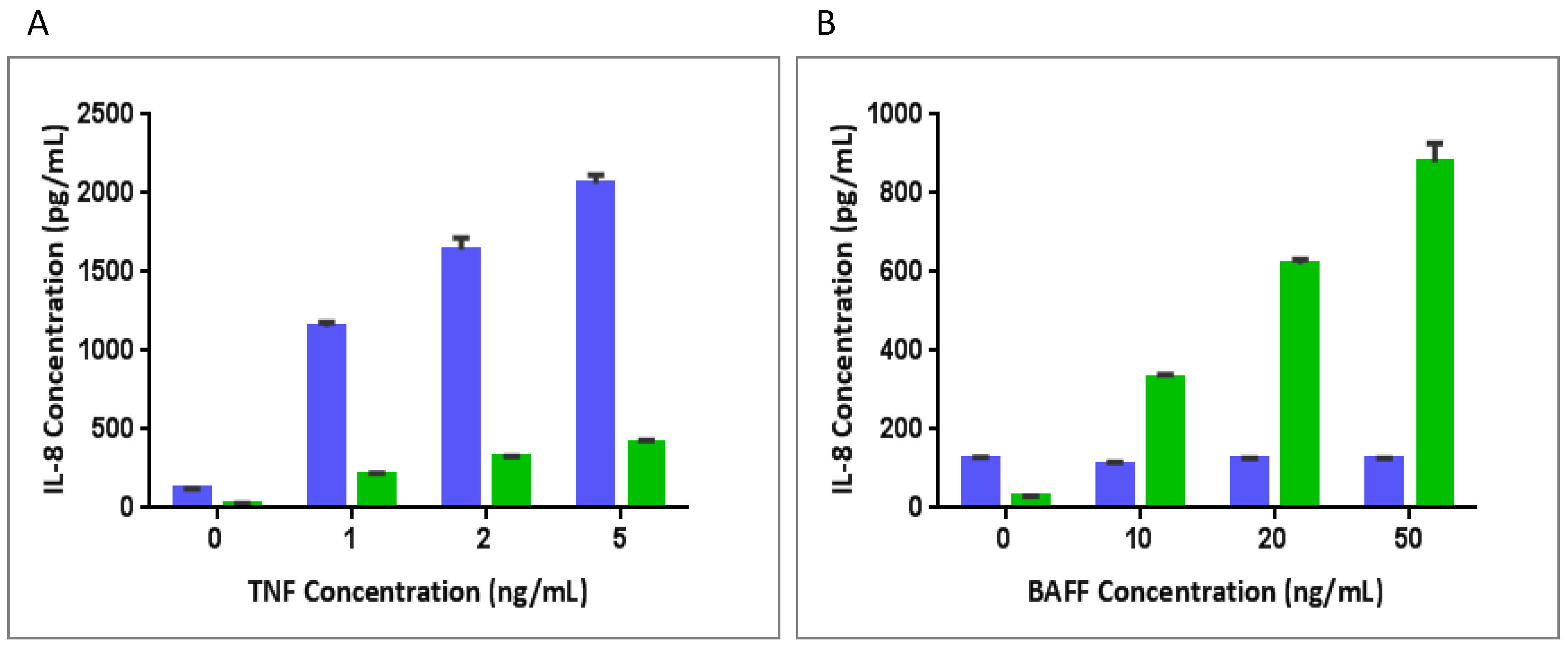
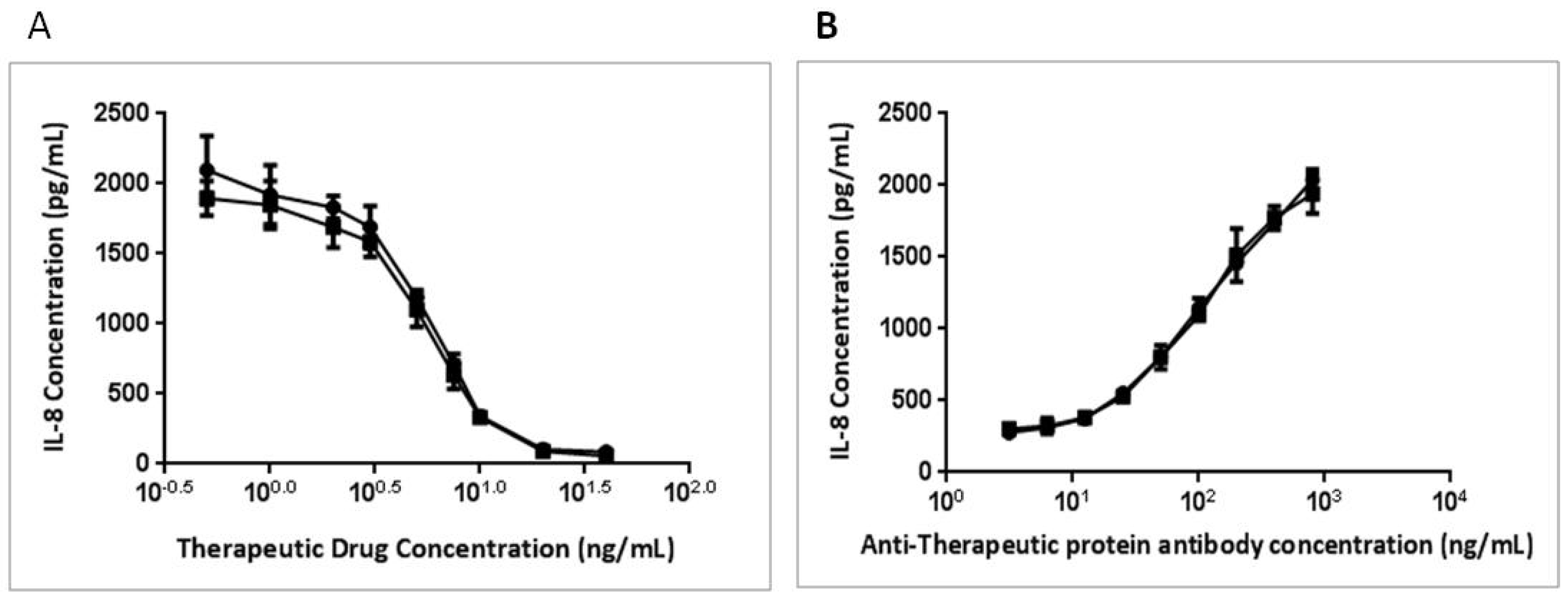
3.4. BAFFR-TNFR1-Mediated BAFF Bioassay Detects BAFF and Neutralizing Antibodies against Anti-BAFF Therapeutic Protein in Serum Samples


4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Mackay, F.; Browning, J.L. BAFF: A fundamental survival factor for B-cells. Nat. Rev. Immunol. 2002, 2, 465–475. [Google Scholar] [CrossRef]
- Gordon, N.C.; Pan, B.; Hymowitz, S.G.; Yin, J.; Kelley, R.F.; Cochran, A.G.; Yan, M.; Dixit, V.M.; Fairbrother, W.J.; Starovasnik, M.A. BAFF/BLyS receptor 3 comprises a minimal TNF receptor-like module that encodes a highly focused ligand-binding site. Biochemistry 2003, 42, 5977–5983. [Google Scholar] [CrossRef]
- Cheema, G.S.; Roschke, V.; Hilbert, D.M.; Stohl, W. Elevated serum B lymphocyte stimulator levels in patients with systemic immune-based rheumatic diseases. Arthritis Rheum. 2001, 44, 1313–1319. [Google Scholar] [CrossRef]
- Matsushita, T.; Hasegawa, M.; Yanaba, K.; Kodera, M.; Takehara, K.; Sato, S. Elevated serum BAFF levels in patients with systemic sclerosis: Enhanced BAFF signaling in systemic sclerosis B lymphocytes. Arthritis Rheum. 2006, 54, 192–201. [Google Scholar] [CrossRef]
- Zhang, J.; Roschke, V.; Baker, K.P.; Wang, Z.; Alarcón, G.S.; Fessler, B.J.; Bastian, H.; Kimberly, R.P.; Zhou, T. Cutting edge: A role for B lymphocyte stimulator in systemic lupus erythematosus. J. Immunol. 2001, 166, 6–10. [Google Scholar]
- Stohl, W. A therapeutic role for BLyS antagonists. Lupus 2004, 13, 317–322. [Google Scholar] [CrossRef]
- Wallace, D.J.; Stohl, W.; Furie, R.A. A phase II, randomized, double-blind, placebo-controlled, dose-ranging study of belimumab in patients with active systemic lupus erythematosus. Arthritis Rheum. 2009, 61, 1168–1178. [Google Scholar] [CrossRef]
- Tak, P.P.; Thurlings, R.M.; Rossier, C. Atacicept in patients with rheumatoid arthritis: Results of a multicenter, phase Ib, double-blind, placebo-controlled, dose-escalating, single- and repeated-dose study. Arthritis Rheum. 2008, 58, 61–72. [Google Scholar] [CrossRef]
- Kreuzaler, M.; Rauch, M.; Salzer, U.; Birmelin, J.; Rizzi, M.; Grimbacher, B.; Plebani, A.; Lougaris, V.; Quinti, I.; Thon, V.; et al. Soluble BAFF levels inversely correlate with peripheral B-cell numbers and the expression of BAFF receptors. J. Immunol. 2012, 188, 497–503. [Google Scholar] [CrossRef]
- Molica, S.; Digiesi, G.; Battaglia, C. Baff serum level predicts time to first treatment in early chronic lymphocytic leukemia. Eur. J. Haematol. 2010, 85, 314–320. [Google Scholar] [CrossRef]
- Lahiri, A.; Pochard, P.; Le Pottier, L. The complexity of the BAFF TNF-family members: implications for autoimmunity. J. Autoimmun. 2012, 39, 189–198. [Google Scholar] [CrossRef]
- Day, E.S.; Cachero, T.G.; Qian, F. Selectivity of BAFF/BLyS and APRIL for binding to the TNF family receptors BAFFR/BR3 and BCMA. Biochemistry 2005, 44, 1919–1931. [Google Scholar] [CrossRef]
- Morrison, M.D.; Reiley, W.; Zhang, M.; Sun, S.C. An atypical tumor necrosis factor (TNF) receptor-associated factor-binding motif of B-cell-activating factor belonging to the TNF family (BAFF) receptor mediates induction of the noncanonical NF-kappaB signaling pathway. J. Biol. Chem. 2005, 280, 10018–10024. [Google Scholar] [CrossRef]
- Yang, M.; Hase, H.; Legarda-Addison, D.; Varughese, L.; Seed, B.; Ting, A.T. B-cell maturation antigen, the receptor for a proliferation-inducing ligand and B-cell-activating factor of the TNF family, induces antigen presentation in B-cells. J. Immunol. 2005, 175, 2814–2824. [Google Scholar]
- McClements, M.; Williams, S.; Ball, C.; Bristow, A.; Wadhwa, M.; Meager, A. A novel bioassay for B-cell activating factor (BAFF) based on expression of a BAFF-receptor ectodomain-tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) receptor-2 endodomain fusion receptor in human rhabdomyosarcoma cells. J. Immunol. Methods 2008, 337, 63–70. [Google Scholar] [CrossRef]
- Thompson, J.S.; Bixler, S.A.; Qian, F. BAFF-R, a newly identified TNF receptor that specifically interacts with BAFF. Science 2001, 293, 2108–2111. [Google Scholar] [CrossRef]
- Miller, D.J.; Hayes, C.E. Phenotypic and genetic characterization of a unique B lymphocyte deficiency in strain A/WySnJ mice. Eur. J. Immunol. 1991, 21, 1123–1130. [Google Scholar] [CrossRef]
- Schiemann, B.; Gommerman, J.L.; Vora, K. An essential role for BAFF in the normal development of B-cells through a BCMA-independent pathway. Science 2001, 293, 2111–2114. [Google Scholar] [CrossRef]
- Aggarwal, B.B. Signaling pathways of the TNF superfamily: A double-edged sword. Nat. Rev. Immunol. 2003, 3, 745–756. [Google Scholar] [CrossRef]
- Chen, Z.J. Ubiquitin signalling in the NF-kappaB pathway. Nat. Cell. Biol. 2005, 7, 758–765. [Google Scholar] [CrossRef]
- Kim, S.H.; Reznikov, L.L.; Stuyt, R.J. Functional reconstitution and regulation of IL-18 activity by the IL-18R beta chain. J. Immunol. 2001, 166, 148–154. [Google Scholar]
- Banner, D.W.; D'Arcy, A.; Janes, W. Crystal structure of the soluble human 55 kd TNF receptor-human TNF beta complex: Implications for TNF receptor activation. Cell 1993, 73, 431–445. [Google Scholar] [CrossRef]
- Fantl, W.J.; Johnson, D.E.; Williams, L.T. Signalling by receptor tyrosine kinases. Annu. Rev. Biochem. 1993, 62, 453–481. [Google Scholar] [CrossRef]
- Adam, D.; Kessler, U.; Krönke, M. Cross-linking of the p55 tumor necrosis factor receptor cytoplasmic domain by a dimeric ligand induces nuclear factor-kappa B and mediates cell death. J. Biol. Chem. 1995, 270, 17482–17487. [Google Scholar]
- Bazzoni, F.; Alejos, E.; Beutler, B. Chimeric tumor necrosis factor receptors with constitutive signaling activity. Proc. Natl. Acad. Sci. USA 1995, 92, 5376–5380. [Google Scholar] [CrossRef]
- Boldin, M.P.; Mett, I.L.; Varfolomeev, E.E. Self-association of the “death domains” of the p55 tumor necrosis factor (TNF) receptor and Fas/APO1 prompts signaling for TNF and Fas/APO1 effects. J. Biol. Chem. 1995, 270, 387–391. [Google Scholar] [CrossRef]
- Gavin, A.L.; Aït-Azzouzene, D.; Ware, CF.; Nemazee, D. DeltaBAFF, an alternate splice isoform that regulates receptor binding and biopresentation of the B-cell survival cytokine, BAFF. J. Biol. Chem. 2003, 278, 38220–38228. [Google Scholar] [CrossRef]
- Casadevall, N.; Nataf, J.; Viron, B.; Kolta, A.; Kiladjian, J.J.; Martin-Dupont, P.; Michaud, P.; Papo, T.; Ugo, V.; Teyssandier, I.; et al. Pure red-cell aplasia and antierythropoietin antibodies in patients treated with recombinant erythropoietin. N. Engl. J. Med. 2002, 346, 469–475. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hu, J.; Yu, Y.; Han, H.; Civoli, F.; Zhuang, Y.; Thomas, J.; Swanson, S.; Jing, S.; Gupta, S. Development of a Novel BAFF Responsive Cell Line Suitable for Detecting Bioactive BAFF and Neutralizing Antibodies against BAFF-Pathway Inhibiting Therapeutics. Cells 2014, 3, 79-91. https://doi.org/10.3390/cells3010079
Hu J, Yu Y, Han H, Civoli F, Zhuang Y, Thomas J, Swanson S, Jing S, Gupta S. Development of a Novel BAFF Responsive Cell Line Suitable for Detecting Bioactive BAFF and Neutralizing Antibodies against BAFF-Pathway Inhibiting Therapeutics. Cells. 2014; 3(1):79-91. https://doi.org/10.3390/cells3010079
Chicago/Turabian StyleHu, Jenny, Yanbin Yu, Hong Han, Francesca Civoli, Yao Zhuang, John Thomas, Steve Swanson, Shuqian Jing, and Shalini Gupta. 2014. "Development of a Novel BAFF Responsive Cell Line Suitable for Detecting Bioactive BAFF and Neutralizing Antibodies against BAFF-Pathway Inhibiting Therapeutics" Cells 3, no. 1: 79-91. https://doi.org/10.3390/cells3010079
APA StyleHu, J., Yu, Y., Han, H., Civoli, F., Zhuang, Y., Thomas, J., Swanson, S., Jing, S., & Gupta, S. (2014). Development of a Novel BAFF Responsive Cell Line Suitable for Detecting Bioactive BAFF and Neutralizing Antibodies against BAFF-Pathway Inhibiting Therapeutics. Cells, 3(1), 79-91. https://doi.org/10.3390/cells3010079