Animal Female Meiosis: The Challenges of Eliminating Centrosomes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
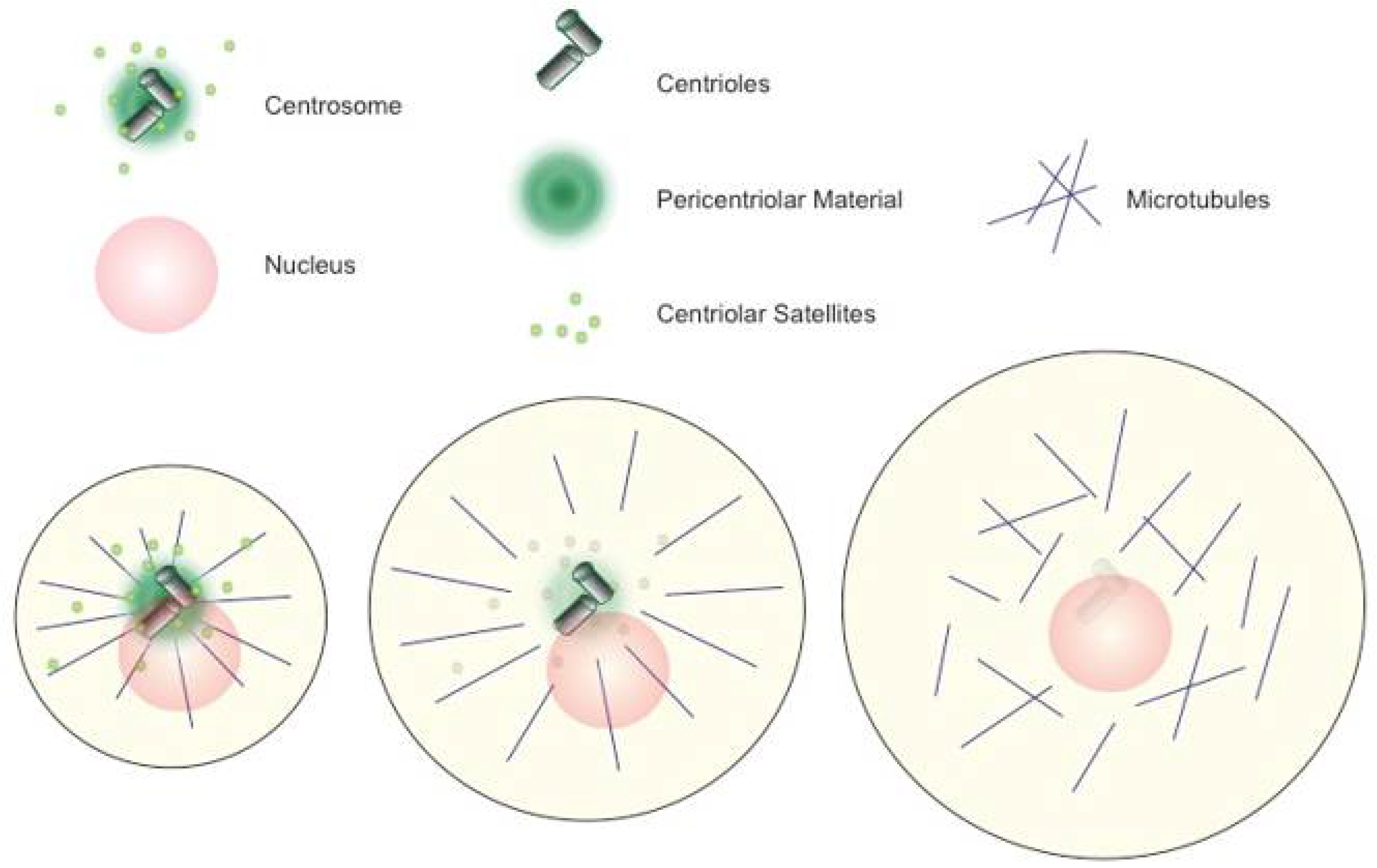
:1. Defining the Biological Problem of Centrosome Elimination in Animal Oocytes
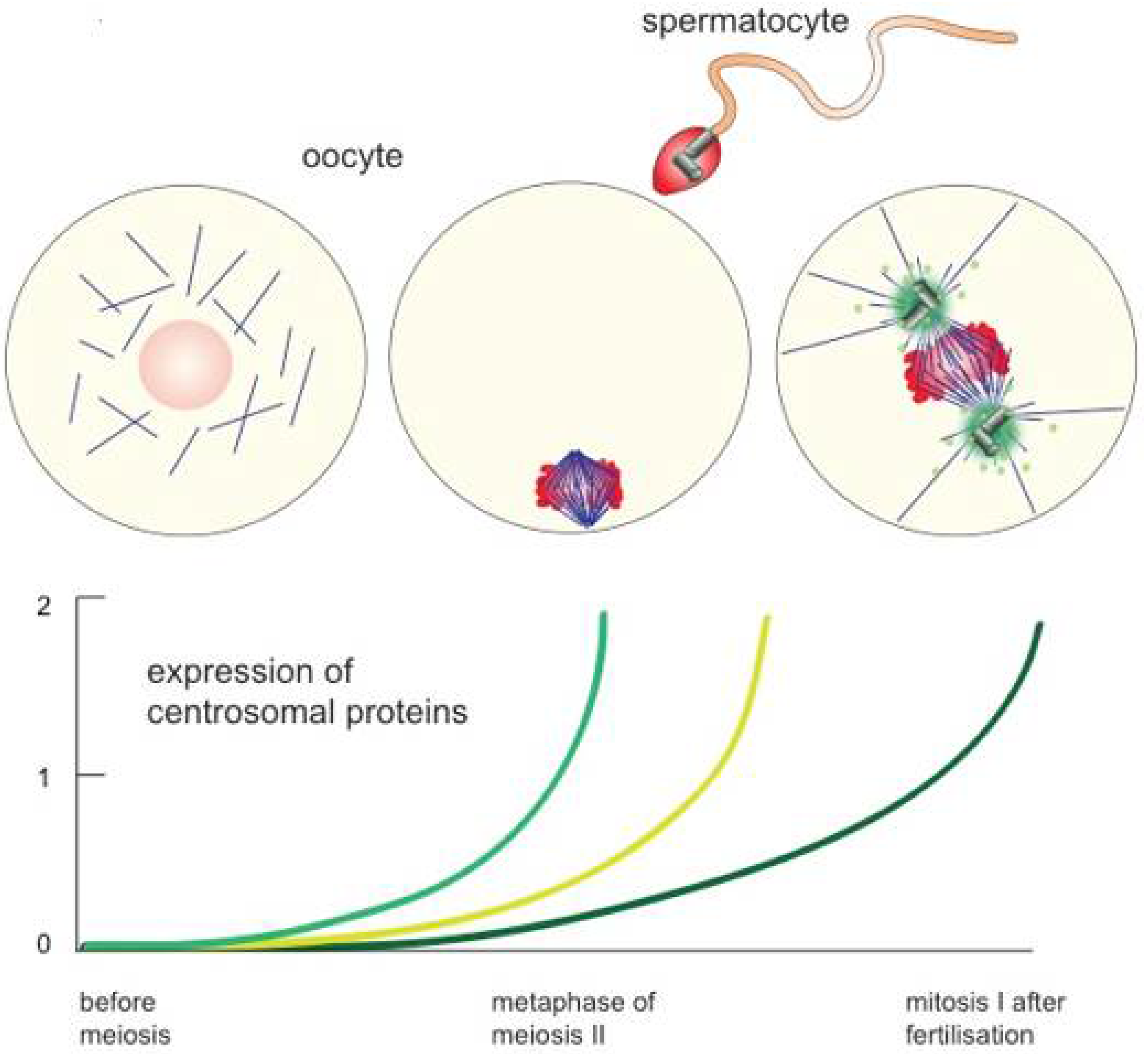
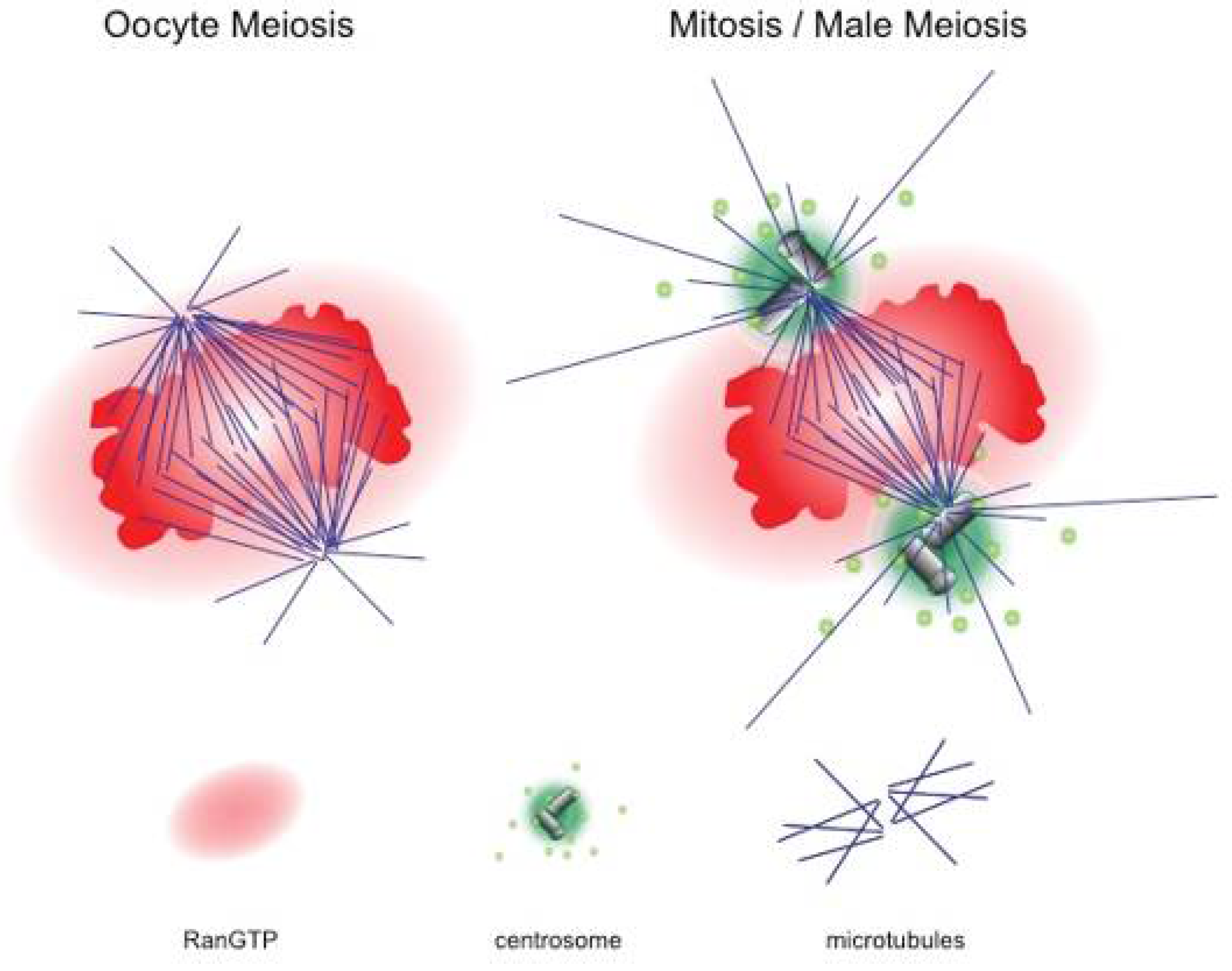
2. Female Meiotic Spindle Formation: Self Assembly of Bipolar Structures
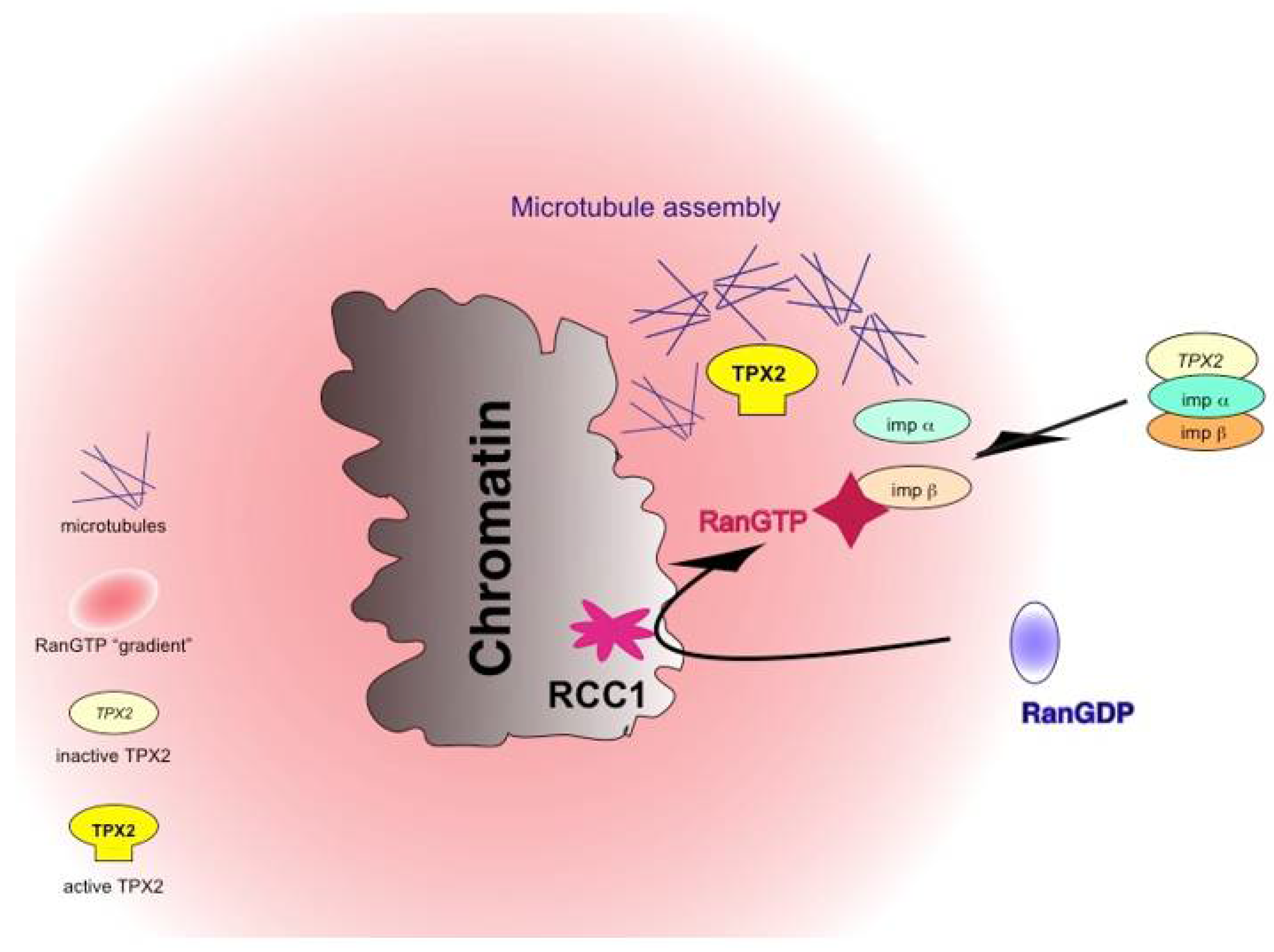
3. The Role of RanGTP in Meiotic Spindle Formation
4. The Mechanism of Centrosome-Independent Microtubule Nucleation
5. The Mechanism of Spindle Organization without Centrosomes
6. Communication between Chromatin and Newly Formed Centrosomes
7. Self-Assembly of Centrioles in Oocytes
8. Concluding Remarks
Funding
Conflicts of Interest
References
- Severson, A.F.; von Dassow, G.; Bowerman, B. Oocyte Meiotic Spindle Assembly and Function. Curr. Top. Dev. Biol. 2016, 116, 65–98. [Google Scholar] [CrossRef] [PubMed]
- Schatten, G. The centrosome and its mode of inheritance: The reduction of the centrosome during Gametogenesis and its restoration during fertilization. Dev. Biol. 1994, 165, 299–335. [Google Scholar] [CrossRef] [PubMed]
- Clift, D.; Schuh, M. Restarting life: Fertilization and the transition from meiosis to mitosis. Nat. Rev. Mol. Cell Biol. 2013, 14, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Loncarek, J.; Bettencourt-Dias, M. Building the right centriole for each cell type. J. Cell Biol. 2018, 217, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Luders, J. The amorphous pericentriolar cloud takes shape. Nat. Cell Biol. 2012, 14, 1126–1128. [Google Scholar] [CrossRef] [PubMed]
- Kollman, J.M.; Merdes, A.; Mourey, L.; Agard, D.A. Microtubule nucleation by gamma-tubulin complexes. Nat. Rev. Mol. Cell Biol. 2011, 12, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.C.; Neuner, A.; Schiebel, E. Targeting of gamma-tubulin complexes to microtubule organizing centers: Conservation and divergence. Trends Cell Biol. 2015, 25, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, J.B.; Wueseke, O.; Hyman, A.A. Pericentriolar material structure and dynamics. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef] [PubMed]
- Fry, A.M.; Sampson, J.; Shak, C.; Shackleton, S. Recent advances in pericentriolar material organization: Ordered layers and scaffolding gels. F1000Research 2017, 6, 1622. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, J.B.; Ferreira Gomes, B.; Widlund, P.O.; Mahamid, J.; Honigmann, A.; Hyman, A.A. The Centrosome Is a Selective Condensate that Nucleates Microtubules by Concentrating Tubulin. Cell 2017, 169, 1066–1077. [Google Scholar] [CrossRef] [PubMed]
- Petry, S. Mechanisms of Mitotic Spindle Assembly. Annu. Rev. Biochem. 2016, 85, 659–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prosser, S.L.; Pelletier, L. Mitotic spindle assembly in animal cells: A fine balancing act. Nat. Rev. Mol. Cell Biol. 2017, 18, 187–201. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, J.R. Mitosis. Cold Spring Harbor Perspect. Biol. 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Nigg, E.A.; Cajanek, L.; Arquint, C. The centrosome duplication cycle in health and disease. FEBS Lett. 2014, 588, 2366–2372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sluder, G. One to only two: A short history of the centrosome and its duplication. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef] [PubMed]
- Firat-Karalar, E.N.; Stearns, T. The centriole duplication cycle. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef]
- Hinchcliffe, E.H. Centrosomes and the art of mitotic spindle maintenance. Int. Rev. Cell Mol. Biol. 2014, 313, 179–217. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Hagan, I.M.; Glover, D.M. The centrosome and its duplication cycle. Cold Spring Harbor Perspect. Biol. 2015, 7, a015800. [Google Scholar] [CrossRef] [PubMed]
- Paz, J.; Luders, J. Microtubule-Organizing Centers: Towards a Minimal Parts List. Trends Cell Biol. 2018, 28, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Nigg, E.A.; Holland, A.J. Once and only once: Mechanisms of centriole duplication and their deregulation in disease. Nat. Rev. Mol. Cell Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Bärenz, F.; Mayilo, D.; Gruss, O.J. Centriolar Satellites: Busy Orbits around the Centrosome. Eur. J. Cell Biol. 2011, 90, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Tollenaere, M.A.; Mailand, N.; Bekker-Jensen, S. Centriolar satellites: Key mediators of centrosome functions. Cell. Mol. Life Sci. 2015, 72, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Hori, A.; Toda, T. Regulation of centriolar satellite integrity and its physiology. Cell. Mol. Life Sci. 2017, 74, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Szollosi, D.; Calarco, P.; Donahue, R.P. Absence of centrioles in the first and second meiotic spindles of mouse oocytes. J. Cell Sci. 1972, 11, 521–541. [Google Scholar] [PubMed]
- Sluder, G.; Miller, F.J.; Lewis, K.; Davison, E.D.; Rieder, C.L. Centrosome inheritance in starfish zygotes: Selective loss of the maternal centrosome after fertilization. Dev. Biol. 1989, 131, 567–579. [Google Scholar] [CrossRef]
- Nakashima, S.; Kato, K.H. Centriole behavior during meiosis in oocytes of the sea urchin Hemicentrotus pulcherrimus. Dev. Growth Differ. 2001, 43, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Shirato, Y.; Tamura, M.; Yoneda, M.; Nemoto, S. Centrosome destined to decay in starfish oocytes. Development 2006, 133, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Sathananthan, A.H.; Selvaraj, K.; Girijashankar, M.L.; Ganesh, V.; Selvaraj, P.; Trounson, A.O. From oogonia to mature oocytes: Inactivation of the maternal centrosome in humans. Microsc. Res. Tech. 2006, 69, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Pimenta-Marques, A.; Bento, I.; Lopes, C.A.; Duarte, P.; Jana, S.C.; Bettencourt-Dias, M. A mechanism for the elimination of the female gamete centrosome in Drosophila melanogaster. Science 2016, 353, aaf4866. [Google Scholar] [CrossRef] [PubMed]
- Washitani-Nemoto, S.; Saitoh, C.; Nemoto, S. Artificial parthenogenesis in starfish eggs: Behavior of nuclei and chromosomes resulting in tetraploidy of parthenogenotes produced by the suppression of polar body extrusion. Dev. Biol. 1994, 163, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Manandhar, G.; Schatten, H.; Sutovsky, P. Centrosome reduction during gametogenesis and its significance. Biol. Reprod. 2005, 72, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Roy, R. Cell cycle regulators control centrosome elimination during oogenesis in Caenorhabditis elegans. J. Cell Biol. 2006, 174, 751–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borrego-Pinto, J.; Somogyi, K.; Karreman, M.A.; Konig, J.; Muller-Reichert, T.; Bettencourt-Dias, M.; Gonczy, P.; Schwab, Y.; Lenart, P. Distinct mechanisms eliminate mother and daughter centrioles in meiosis of starfish oocytes. J. Cell Biol. 2016, 212, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Schoborg, T.A.; Rusan, N.M. Taking Centrioles to the Elimination Round. Dev. Cell 2016, 38, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Moorhouse, K.S.; Burgess, D.R. How to be at the right place at the right time: The importance of spindle positioning in embryos. Mol. Reprod. Dev. 2014, 81, 884–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaigne, A.; Terret, M.E.; Verlhac, M.H. Asymmetries and Symmetries in the Mouse Oocyte and Zygote. Results Probl. Cell Differ. 2017, 61, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Hasley, A.; Chavez, S.; Danilchik, M.; Wuhr, M.; Pelegri, F. Vertebrate Embryonic Cleavage Pattern Determination. Adv. Exp. Med. Biol. 2017, 953, 117–171. [Google Scholar] [CrossRef] [PubMed]
- Cavazza, T.; Peset, I.; Vernos, I. From meiosis to mitosis—The sperm centrosome defines the kinetics of spindle assembly after fertilization in Xenopus. J. Cell Sci. 2016, 129, 2538–2547. [Google Scholar] [CrossRef] [PubMed]
- Inoue, D.; Wittbrodt, J.; Gruss, O.J. Loss and Rebirth of the Animal Microtubule Organizing Center: How Maternal Expression of Centrosomal Proteins Cooperates with the Sperm Centriole in Zygotic Centrosome Reformation. Bioessays 2018, 40, e1700135. [Google Scholar] [CrossRef] [PubMed]
- Avidor-Reiss, T.; Gopalakrishnan, J. Building a centriole. Curr. Opin. Cell Biol. 2013, 25, 72–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verlhac, M.H.; Terret, M.E. Oocyte Maturation and Development. F1000Research 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Hunt, P.A. Meiosis in mammals: Recombination, non-disjunction and the environment. Biochem. Soc. Trans. 2006, 34, 574–577. [Google Scholar] [CrossRef] [PubMed]
- Pacchierotti, F.; Adler, I.D.; Eichenlaub-Ritter, U.; Mailhes, J.B. Gender effects on the incidence of aneuploidy in mammalian germ cells. Environ. Res. 2007, 104, 46–69. [Google Scholar] [CrossRef] [PubMed]
- Webster, A.; Schuh, M. Mechanisms of Aneuploidy in Human Eggs. Trends Cell Biol. 2017, 27, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Greaney, J.; Wei, Z.; Homer, H. Regulation of chromosome segregation in oocytes and the cellular basis for female meiotic errors. Hum. Reprod. Update 2017. [Google Scholar] [CrossRef] [PubMed]
- Karsenti, E.; Newport, J.; Hubble, R.; Kirschner, M. Interconversion of metaphase and interphase microtubule arrays, as studied by the injection of centrosomes and nuclei into Xenopus eggs. J. Cell Biol. 1984, 98, 1730–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karsenti, E.; Newport, J.; Kirschner, M. The respective roles of centrosomes and chromatin in the conversion of microtubule arrays from interphase to metaphase. J. Cell Biol. 1984, 99, 47s–54s. [Google Scholar] [CrossRef] [PubMed]
- Karsenti, E.; Vernos, I. The mitotic spindle: A self-made machine. Science 2001, 294, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Hallen, M.A.; Endow, S.A. Anastral spindle assembly: A mathematical model. Biophys. J. 2009, 97, 2191–2201. [Google Scholar] [CrossRef] [PubMed]
- Masui, Y.; Markert, C.L. Cytoplasmic control of nuclear behavior during meiotic maturation of frog oocytes. J. Exp. Zool. 1971, 177, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Murray, A. Xenopus Laevis: Practical Uses in Cell and Molecular Biology; Kay, B.K., Peng, H.B., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1991; Volume 36, pp. 581–605. [Google Scholar]
- Sawin, K.E.; Mitchison, T.J. Mitotic spindle assembly by two different pathways in vitro. J. Cell Biol. 1991, 112, 925–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heald, R.; Tournebize, R.; Blank, T.; Sandaltzopoulos, R.; Becker, P.; Hyman, A.; Karsenti, E. Self-organization of microtubules into bipolar spindles around artificial chromosomes in Xenopus egg extracts. Nature 1996, 382, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Heald, R.; Tournebize, R.; Habermann, A.; Karsenti, E.; Hyman, A. Spindle assembly in Xenopus egg extracts: Respective roles of centrosomes and microtubule self-organization. J. Cell Biol. 1997, 138, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Cavazza, T.; Vernos, I. The RanGTP Pathway: From Nucleo-Cytoplasmic Transport to Spindle Assembly and Beyond. Front. Cell Dev. Biol. 2015, 3, 82. [Google Scholar] [CrossRef] [PubMed]
- Carazo-Salas, R.E.; Guarguaglini, G.; Gruss, O.J.; Segref, A.; Karsenti, E.; Mattaj, I.W. Generation of GTP-bound Ran by RCC1 is required for chromatin-induced mitotic spindle formation. Nature 1999, 400, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Gruss, O.J.; Wittmann, M.; Yokoyama, H.; Pepperkok, R.; Kufer, T.; Silljé, H.; Karsenti, E.; Mattaj, I.W.; Vernos, I. Chromosome-induced microtubule assembly mediated by TPX2 is required for spindle formation in HeLa cells. Nat. Cell Biol. 2002, 4, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Wilde, A.; Zheng, Y. Stimulation of microtubule aster formation and spindle assembly by the small GTPase Ran. Science 1999, 284, 1359–1362. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Heidebrecht, H.; Rott, A.; Schlegelberger, B.; Parwaresch, R. Assignment of human proliferation associated p100 gene (C20orf1) to human chromosome band 20q11.2 by in situ hybridization. Cytogenet. Cell Genet. 1999, 84, 182–183. [Google Scholar] [CrossRef] [PubMed]
- Kalab, P.; Pu, R.T.; Dasso, M. The ran GTPase regulates mitotic spindle assembly. Curr. Biol. 1999, 9, 481–484. [Google Scholar] [CrossRef]
- Carazo-Salas, R.E.; Gruss, O.J.; Mattaj, I.W.; Karsenti, E. Ran-GTP coordinates regulation of microtubule nucleation and dynamics during mitotic-spindle assembly. Nat. Cell Biol. 2001, 3, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Wilde, A.; Lizarraga, S.B.; Zhang, L.; Wiese, C.; Gliksman, N.R.; Walczak, C.E.; Zheng, Y. Ran stimulates spindle assembly by altering microtubule dynamics and the balance of motor activities. Nat. Cell Biol. 2001, 3, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Ohba, T.; Nakamura, M.; Nishitani, H.; Nishimoto, T. Self-organization of microtubule asters induced in Xenopus egg extracts by GTP-bound Ran. Science 1999, 284, 1356–1358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Hughes, M.; Clarke, P.R. Ran-GTP stabilises microtubule asters and inhibits nuclear assembly in Xenopus egg extracts. J. Cell Sci. 1999, 112, 2453–2461. [Google Scholar] [PubMed]
- Gruss, O.J.; Carazo-Salas, R.E.; Schatz, C.A.; Guarguaglini, G.; Kast, J.; Wilm, M.; Le Bot, N.; Vernos, I.; Karsenti, E.; Mattaj, I.W. Ran Induces Spindle Assembly by Reversing the Inhibitory Effect of Importin alpha on TPX2 Activity. Cell 2001, 104, 83–93. [Google Scholar] [CrossRef]
- Nachury, M.V.; Maresca, T.J.; Salmon, W.C.; Waterman-Storer, C.M.; Heald, R.; Weis, K. Importin beta Is a Mitotic Target of the Small GTPase Ran in Spindle Assembly. Cell 2001, 104, 95–106. [Google Scholar] [CrossRef]
- Wiese, C.; Wilde, A.; Moore, M.S.; Adam, S.A.; Merdes, A.; Zheng, Y. Role of Importin b in Coupling Ran to Downstream Targets in Microtubule Assembly. Science 2001, 4, 4–8. [Google Scholar] [CrossRef]
- Hetzer, M.; Gruss, O.J.; Mattaj, I.W. The Ran GTPase as a marker for chromosome position in spindle formation and nuclear envelope assembly. Nat. Cell Biol. 2002, 4, E177–E184. [Google Scholar] [CrossRef] [PubMed]
- Giesecke, A.; Stewart, M. Novel binding of the mitotic regulator TPX2 (target protein for Xenopus kinesin-like protein 2) to importin-alpha. J. Biol. Chem. 2010, 285, 17628–17635. [Google Scholar] [CrossRef] [PubMed]
- Gruss, O.J. Ras Superfamily Small G Proteins: Biology and Mechanisms; Wittinghofer, A., Ed.; Springer: Basel, Switzerland, 2014; Volume 2, pp. 125–150. [Google Scholar]
- Okada, N.; Sato, M. Spatiotemporal Regulation of Nuclear Transport Machinery and Microtubule Organization. Cells 2015, 4, 406–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, D.J.; Travesa, A.; Nord, M.S.; Bernis, C. Nuclear transport factors: Global regulation of mitosis. Curr. Opin. Cell Biol. 2015, 35, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Meunier, S.; Shvedunova, M.; Van Nguyen, N.; Avila, L.; Vernos, I.; Akhtar, A. An epigenetic regulator emerges as microtubule minus-end binding and stabilizing factor in mitosis. Nat. Commun. 2015, 6, 7889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, M.Y.; Wiese, C.; Cao, K.; Martin, O.; Donovan, P.; Ruderman, J.; Pringent, C.; Zheng, Y. A Ran signalling pathway mediated by the mitotic kinase Aurora A in spindle assembly. Nat. Cell Biol. 2003, 5, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Kufer, T.A.; Sillje, H.H.; Korner, R.; Gruss, O.J.; Meraldi, P.; Nigg, E.A. Human TPX2 is required for targeting Aurora-A kinase to the spindle. J. Cell Biol. 2002, 158, 617–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eyers, P.A.; Erikson, E.; Chen, L.G.; Maller, J.L. A novel mechanism for activation of the protein kinase Aurora A. Curr. Biol. 2003, 13, 691–697. [Google Scholar] [CrossRef]
- Bayliss, R.; Sardon, T.; Vernos, I.; Conti, E. Structural Basis of Aurora-A Activation by TPX2 at the Mitotic Spindle. Mol. Cell 2003, 12, 851–862. [Google Scholar] [CrossRef]
- Bayliss, R.; Sardon, T.; Ebert, J.; Lindner, D.; Vernos, I.; Conti, E. Determinants for Aurora-A activation and Aurora-B discrimination by TPX2. Cell Cycle 2004, 3, 404–407. [Google Scholar] [CrossRef] [PubMed]
- Eyers, P.A.; Maller, J.L. Regulation of Xenopus Aurora A activation by TPX2. J. Biol. Chem. 2004, 279, 9008–9015. [Google Scholar] [CrossRef] [PubMed]
- Barr, A.R.; Gergely, F. Aurora-A: The maker and breaker of spindle poles. J. Cell Sci. 2007, 120, 2987–2996. [Google Scholar] [CrossRef] [PubMed]
- Scrofani, J.; Sardon, T.; Meunier, S.; Vernos, I. Microtubule nucleation in mitosis by a RanGTP-dependent protein complex. Curr. Biol. 2015, 25, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Roostalu, J.; Surrey, T.; Nogales, E. Structural insight into TPX2-stimulated microtubule assembly. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsia, K.C.; Wilson-Kubalek, E.M.; Dottore, A.; Hao, Q.; Tsai, K.L.; Forth, S.; Shimamoto, Y.; Milligan, R.A.; Kapoor, T.M. Reconstitution of the augmin complex provides insights into its architecture and function. Nat. Cell Biol. 2014, 16, 852–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petry, S.; Groen, A.C.; Ishihara, K.; Mitchison, T.J.; Vale, R.D. Branching microtubule nucleation in Xenopus egg extracts mediated by augmin and TPX2. Cell 2013, 152, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Farache, D.; Emorine, L.; Haren, L.; Merdes, A. Assembly and regulation of gamma-tubulin complexes. Open Biol. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Huertas, C.; Luders, J. The augmin connection in the geometry of microtubule networks. Curr. Biol. 2015, 25, R294–R299. [Google Scholar] [CrossRef] [PubMed]
- Clausen, T.; Ribbeck, K. Self-organization of anastral spindles by synergy of dynamic instability, autocatalytic microtubule production, and a spatial signaling gradient. PLoS ONE 2007, 2, e244. [Google Scholar] [CrossRef] [PubMed]
- Petry, S.; Pugieux, C.; Nedelec, F.J.; Vale, R.D. Augmin promotes meiotic spindle formation and bipolarity in Xenopus egg extracts. Proc. Natl. Acad. Sci. USA 2011, 108, 14473–14478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bärenz, F.; Inoue, D.; Yokoyama, H.; Tegha-Dunghu, J.; Freiss, S.; Draeger, S.; Mayilo, D.; Cado, I.; Merker, S.; Klinger, M.; et al. The centriolar satellite protein SSX2IP promotes centrosome maturation. J. Cell Biol. 2013, 202, 81–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfaro-Aco, R.; Thawani, A.; Petry, S. Structural analysis of the role of TPX2 in branching microtubule nucleation. J. Cell Biol. 2017, 216, 983–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumont, J.; Petri, S.; Pellegrin, F.; Terret, M.E.; Bohnsack, M.T.; Rassinier, P.; Georget, V.; Kalab, P.; Gruss, O.J.; Verlhac, M.H. A centriole- and RanGTP-independent spindle assembly pathway in meiosis I of vertebrate oocytes. J. Cell Biol. 2007, 176, 295–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesario, J.; McKim, K.S. RanGTP is required for meiotic spindle organization and the initiation of embryonic development in Drosophila. J. Cell Sci. 2011, 124, 3797–3810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holubcova, Z.; Blayney, M.; Elder, K.; Schuh, M. Human oocytes. Error-prone chromosome-mediated spindle assembly favors chromosome segregation defects in human oocytes. Science 2015, 348, 1143–1147. [Google Scholar] [CrossRef] [PubMed]
- Sampath, S.C.; Ohi, R.; Leismann, O.; Salic, A.; Pozniakovski, A.; Funabiki, H. The chromosomal passenger complex is required for chromatin-induced microtubule stabilization and spindle assembly. Cell 2004, 118, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.E.; Sampath, S.C.; Maniar, T.A.; Woo, E.M.; Chait, B.T.; Funabiki, H. Chromosomal enrichment and activation of the aurora B pathway are coupled to spatially regulate spindle assembly. Dev. Cell 2007, 12, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Tseng, B.S.; Tan, L.; Kapoor, T.M.; Funabiki, H. Dual detection of chromosomes and microtubules by the chromosomal passenger complex drives spindle assembly. Dev. Cell 2010, 18, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Radford, S.J.; Jang, J.K.; McKim, K.S. The chromosomal passenger complex is required for meiotic acentrosomal spindle assembly and chromosome biorientation. Genetics 2012, 192, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Shah, S.J.; Fan, B.; Paik, D.; DiSanto, D.J.; Hinman, A.M.; Cesario, J.M.; Battaglia, R.A.; Demos, N.; McKim, K.S. Spindle Assembly and Chromosome Segregation Requires Central Spindle Proteins in Drosophila Oocytes. Genetics 2016, 202, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Blower, M.D.; Nachury, M.; Heald, R.; Weis, K. A Rae1-containing ribonucleoprotein complex is required for mitotic spindle assembly. Cell 2005, 121, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Merdes, A.; Cleveland, D.W. Pathways of spindle pole formation: Different mechanisms; conserved components. J. Cell Biol. 1997, 138, 953–956. [Google Scholar] [CrossRef] [PubMed]
- Walczak, C.E.; Vernos, I.; Mitchison, T.J.; Karsenti, E.; Heald, R. A model for the proposed roles of different microtubule-based motor proteins in establishing spindle bipolarity. Curr. Biol. 1998, 8, 903–913. [Google Scholar] [CrossRef]
- Kapoor, T.M.; Mayer, T.U.; Coughlin, M.L.; Mitchison, T.J. Probing spindle assembly mechanisms with monastrol, a small molecule inhibitor of the mitotic kinesin, Eg5. J. Cell Biol. 2000, 150, 975–988. [Google Scholar] [CrossRef] [PubMed]
- Mailhes, J.B.; Mastromatteo, C.; Fuseler, J.W. Transient exposure to the Eg5 kinesin inhibitor monastrol leads to syntelic orientation of chromosomes and aneuploidy in mouse oocytes. Mutat Res. 2004, 559, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Schuh, M.; Ellenberg, J. Self-organization of MTOCs replaces centrosome function during acentrosomal spindle assembly in live mouse oocytes. Cell 2007, 130, 484–498. [Google Scholar] [CrossRef] [PubMed]
- Koffa, M.D.; Casanova, C.M.; Santarella, R.; Kocher, T.; Wilm, M.; Mattaj, I.W. HURP is part of a Ran-dependent complex involved in spindle formation. Curr. Biol. 2006, 16, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Sillje, H.H.; Nagel, S.; Korner, R.; Nigg, E.A. HURP is a Ran-importin beta-regulated protein that stabilizes kinetochore microtubules in the vicinity of chromosomes. Curr. Biol. 2006, 16, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Breuer, M.; Kolano, A.; Kwon, M.; Li, C.C.; Tsai, T.F.; Pellman, D.; Brunet, S.; Verlhac, M.H. HURP permits MTOC sorting for robust meiotic spindle bipolarity, similar to extra centrosome clustering in cancer cells. J. Cell Biol. 2010, 191, 1251–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luksza, M.; Queguigner, I.; Verlhac, M.H.; Brunet, S. Rebuilding MTOCs upon centriole loss during mouse oogenesis. Dev. Biol. 2013, 382, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Clift, D.; Schuh, M. A three-step MTOC fragmentation mechanism facilitates bipolar spindle assembly in mouse oocytes. Nat. Commun. 2015, 6, 7217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, H.; Rybina, S.; Santarella-Mellwig, R.; Mattaj, I.W.; Karsenti, E. ISWI is a RanGTP-dependent MAP required for chromosome segregation. J. Cell Biol. 2009, 187, 813–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Dawe, R.K. Mechanisms of plant spindle formation. Chromosom. Res. 2011, 19, 335–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.W.; Barker, A.R.; Wakefield, J.G. The Ran Pathway in Drosophila melanogaster Mitosis. Front. Cell Dev. Biol. 2015, 3, 74. [Google Scholar] [CrossRef] [PubMed]
- Bennabi, I.; Terret, M.E.; Verlhac, M.H. Meiotic spindle assembly and chromosome segregation in oocytes. J. Cell Biol. 2016, 215, 611–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heald, R.; Gibeaux, R. Subcellular scaling: Does size matter for cell division? Curr. Opin. Cell Biol. 2018, 52, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Kalab, P.; Solc, P.; Motlik, J. The role of RanGTP gradient in vertebrate oocyte maturation. Results Probl. Cell Differ. 2011, 53, 235–267. [Google Scholar] [CrossRef] [PubMed]
- Weaver, L.N.; Walczak, C.E. Spatial gradients controlling spindle assembly. Biochem. Soc. Trans. 2015, 43, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.S.; Arnaoutov, A.; Dasso, M. RanBP1 governs spindle assembly by defining mitotic Ran-GTP production. Dev. Cell 2014, 31, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, H.; Gruss, O.J.; Rybina, S.; Caudron, M.; Schelder, M.; Wilm, M.; Mattaj, I.W.; Karsenti, E. Cdk11 is a RanGTP-dependent microtubule stabilization factor that regulates spindle assembly rate. J. Cell Biol. 2008, 180, 867–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavazza, T.; Malgaretti, P.; Vernos, I. The sequential activation of the mitotic microtubule assembly pathways favors bipolar spindle formation. Mol. Biol. Cell. 2016, 27, 2935–2945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arquint, C.; Gabryjonczyk, A.M.; Nigg, E.A. Centrosomes as signalling centres. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, H.; Gruss, O.J. New mitotic regulators released from chromatin. Front. Oncol. 2013, 3, 308. [Google Scholar] [CrossRef] [PubMed]
- Beck, M.; Hurt, E. The nuclear pore complex: Understanding its function through structural insight. Nat. Rev. Mol. Cell Biol. 2017, 18, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Imamoto, N.; Funakoshi, T. Nuclear pore dynamics during the cell cycle. Curr. Opin. Cell Biol. 2012, 24, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Franz, C.; Walczak, R.; Yavuz, S.; Santarella, R.; Gentzel, M.; Askjaer, P.; Galy, V.; Hetzer, M.; Mattaj, I.W.; Antonin, W. MEL-28/ELYS is required for the recruitment of nucleoporins to chromatin and postmitotic nuclear pore complex assembly. EMBO Rep. 2007, 8, 165–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, H.; Koch, B.; Walczak, R.; Ciray-Duygu, F.; Gonzalez-Sanchez, J.C.; Devos, D.P.; Mattaj, I.W.; Gruss, O.J. The nucleoporin MEL-28 promotes RanGTP-dependent gamma-tubulin recruitment and microtubule nucleation in mitotic spindle formation. Nat. Commun. 2014, 5, 3270. [Google Scholar] [CrossRef] [PubMed]
- Eckerdt, F.; Yamamoto, T.M.; Lewellyn, A.L.; Maller, J.L. Identification of a polo-like kinase 4-dependent pathway for de novo centriole formation. Curr. Biol. 2011, 21, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Kallenbach, R.; Mazia, D. Origin and maturation of centrioles in association with the nuclear envelope in hypertonic stressed sea urchin eggs. Eur. J. Cell Biol. 1982, 28, 68–76. [Google Scholar] [PubMed]
- Kallenbach, R.J. The induction of de novo centrioles in sea urchin eggs: A possible common mechanism for centriolar activation among parthenogenetic procedures. Eur. J. Cell Biol. 1983, 30, 159–166. [Google Scholar] [PubMed]
- Courtois, A.; Schuh, M.; Ellenberg, J.; Hiiragi, T. The transition from meiotic to mitotic spindle assembly is gradual during early mammalian development. J. Cell Biol. 2012, 198, 357–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, C.M.; Lenart, P. Bulk cytoplasmic actin and its functions in meiosis and mitosis. Curr. Biol. 2011, 21, R825–R830. [Google Scholar] [CrossRef] [PubMed]
- Yi, K.; Li, R. Actin cytoskeleton in cell polarity and asymmetric division during mouse oocyte maturation. Cytoskeleton (Hoboken) 2012, 69, 727–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Albertini, D.F. The road to maturation: Somatic cell interaction and self-organization of the mammalian oocyte. Nat. Rev. Mol. Cell Biol. 2013, 14, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Almonacid, M.; Terret, M.E.; Verlhac, M.H. Actin-based spindle positioning: New insights from female gametes. J. Cell Sci. 2014, 127, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Carabatsos, M.J.; Combelles, C.M.; Messinger, S.M.; Albertini, D.F. Sorting and reorganization of centrosomes during oocyte maturation in the mouse. Microsc. Res. Tech. 2000, 49, 435–444. [Google Scholar] [CrossRef]
- Luo, Y.B.; Kim, N.H. PLK4 is essential for meiotic resumption in mouse oocytes. Biol. Reprod. 2015, 92, 101. [Google Scholar] [CrossRef] [PubMed]
- Bury, L.; Coelho, P.A.; Simeone, A.; Ferries, S.; Eyers, C.E.; Eyers, P.A.; Zernicka-Goetz, M.; Glover, D.M. Plk4 and Aurora A cooperate in the initiation of acentriolar spindle assembly in mammalian oocytes. J. Cell Biol. 2017, 216, 3571–3590. [Google Scholar] [CrossRef] [PubMed]
- Coelho, P.A.; Bury, L.; Sharif, B.; Riparbelli, M.G.; Fu, J.; Callaini, G.; Glover, D.M.; Zernicka-Goetz, M. Spindle formation in the mouse embryo requires Plk4 in the absence of centrioles. Dev. Cell 2013, 27, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.W.; Jo, Y.J.; Jung, S.M.; Wang, H.Y.; Kim, N.H.; Namgoong, S. Distinct roles of Cep192 and Cep152 in acentriolar MTOCs and spindle formation during mouse oocyte maturation. FASEB J. 2017. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gruss, O.J. Animal Female Meiosis: The Challenges of Eliminating Centrosomes. Cells 2018, 7, 73. https://doi.org/10.3390/cells7070073
Gruss OJ. Animal Female Meiosis: The Challenges of Eliminating Centrosomes. Cells. 2018; 7(7):73. https://doi.org/10.3390/cells7070073
Chicago/Turabian StyleGruss, Oliver J. 2018. "Animal Female Meiosis: The Challenges of Eliminating Centrosomes" Cells 7, no. 7: 73. https://doi.org/10.3390/cells7070073
APA StyleGruss, O. J. (2018). Animal Female Meiosis: The Challenges of Eliminating Centrosomes. Cells, 7(7), 73. https://doi.org/10.3390/cells7070073