Exercise Exacerbates the Transcriptional Profile of Hypoxia, Oxidative Stress and Inflammation in Rats with Adjuvant-Induced Arthritis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Study Groups
2.2. Arthritis Induction
2.3. Familiarization
2.4. Physical Capacity Test
2.5. Exercise Program
2.6. Histological Analysis
2.7. DNA Microarray and Bioinformatics Analysis
2.8. Statistical Analysis
3. Results
3.1. Maximal Physical Capacity
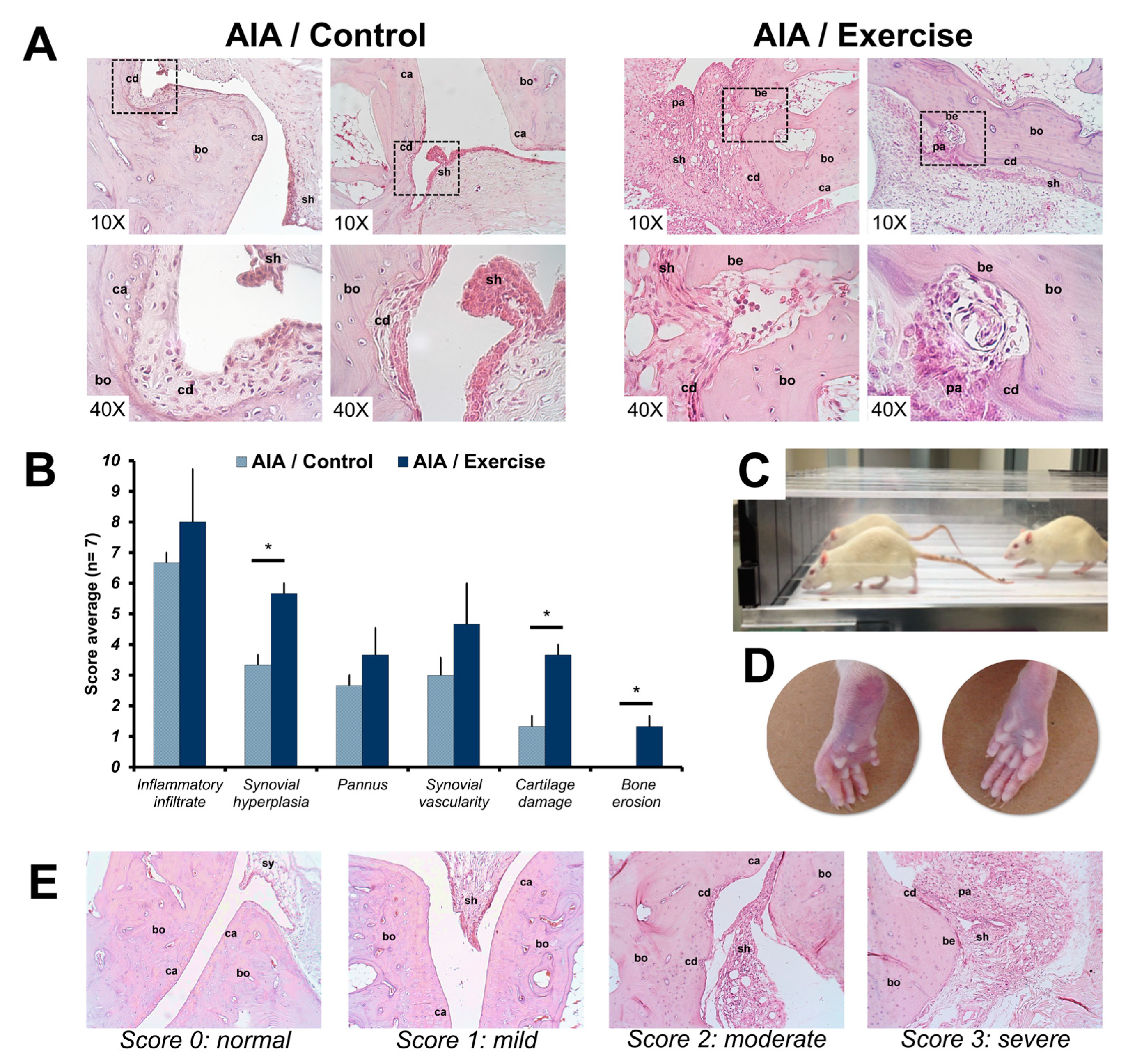
3.2. Histological Analysis
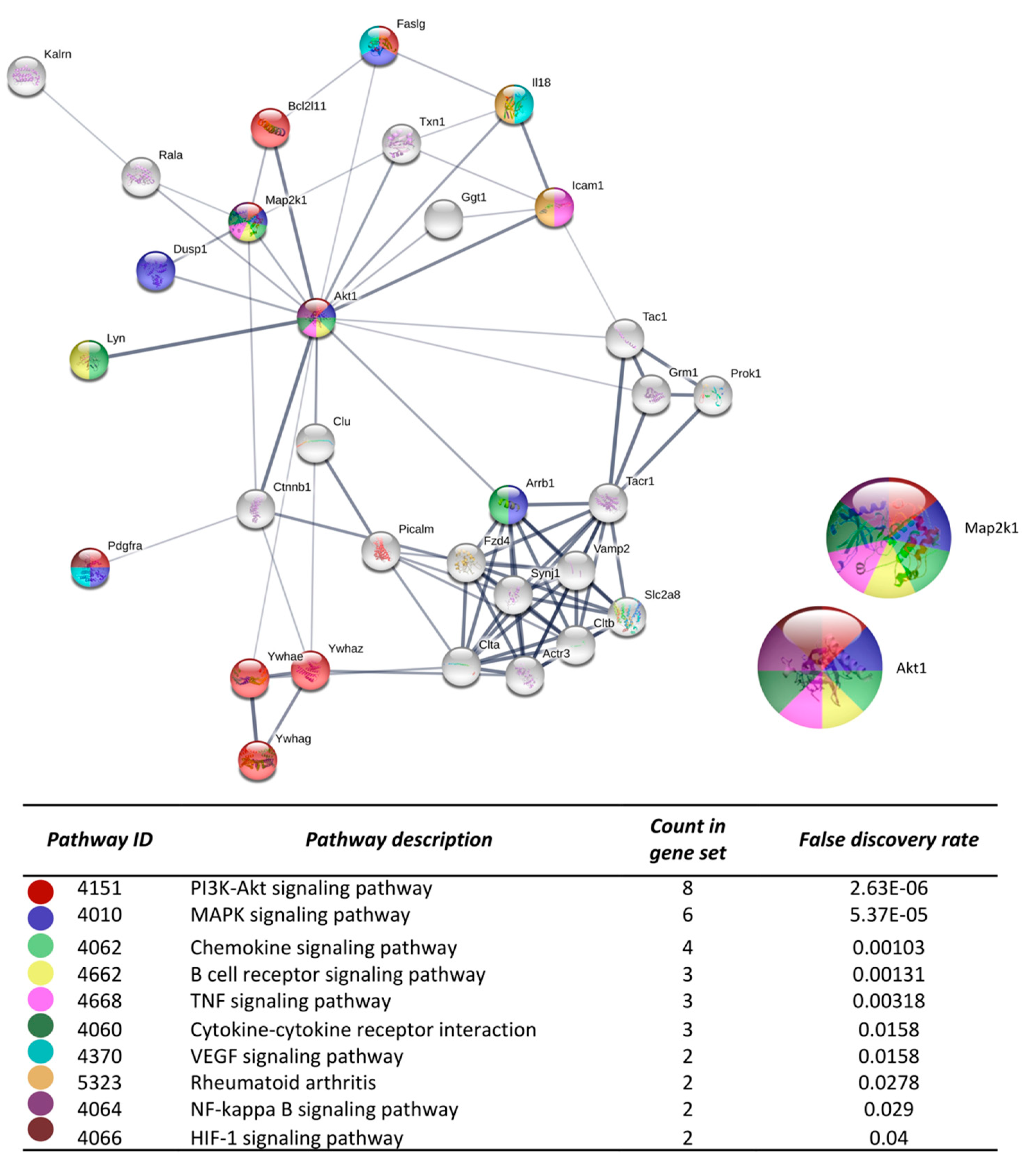
3.3. Microarray and Bioinformatic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. List of Genes Up-Regulated by PE Effect
| Gene Id | Gene | Z-Score |
| AA818786 | No data | 1.770223 |
| AA963507 | No data | 1.500387 |
| AB000928 | zona pellucida glycoprotein 1(Zp1) | 3.330347 |
| AB003753 | No data | 2.348897 |
| AB012139 | bone morphogenetic protein 1(Bmp1) | 1.717532 |
| AB022014 | proteasome 26S subunit, non-ATPase 10(Psmd10) | 1.800734 |
| AB024333 | barrier to autointegration factor 1(Banf1) | 2.409521 |
| AB027143 | septin 5(Sept5) | 2.332071 |
| AB037248 | ATPase H+ transporting V0 subunit e1(Atp6v0e1) | 1.790973 |
| AB041723 | apoptosis inducing factor, mitochondria associated 1(Aifm1) | 2.20147 |
| AB043959 | ubiquitin C-terminal hydrolase L3(Uchl3) | 2.083474 |
| AB070355 | kinesin family member 1B(Kif1b) | 2.560659 |
| AB073318 | brain and acute leukemia, cytoplasmic(Baalc) | 1.825273 |
| AF000139 | cytochrome P450, family 27, subfamily b, polypeptide 1(Cyp27b1) | 3.142163 |
| AF016183 | vomeronasal 2 receptor, pseudogene 45(Vom2r-ps45) | 2.284991 |
| AF026554 | solute carrier family 5 member 6(Slc5a6) | 1.552609 |
| AF030243 | interleukin 3 receptor subunit alpha(Il3ra) | 1.890042 |
| AF041374 | phosphatidylinositol binding clathrin assembly protein(Picalm) | 2.019124 |
| AF053989 | vomeronasal 2 receptor 44(Vom2r44) | 1.534172 |
| AF059678 | replication factor C subunit 1(Rfc1) | 1.961506 |
| AF063103 | adhesion G protein-coupled receptor L3(Adgrl3) | 1.510015 |
| AF063890 | protein tyrosine kinase 2 beta(Ptk2b) | 2.392998 |
| AF065161 | cytokine inducible SH2-containing protein(Cish) | 2.074343 |
| AF102262 | beta-1,4-galactosyltransferase 1(B4galt1) | 1.819501 |
| AF106657 | ubiquitin specific peptidase 15(Usp15) | 2.061666 |
| AF115768 | defensin alpha 24(Defa24) | 2.430658 |
| AF121265 | catenin beta 1(Ctnnb1) | 1.995608 |
| AF121893 | single stranded DNA binding protein 3(Ssbp3) | 1.660423 |
| AF145050 | eukaryotic translation elongation factor 1 delta(Eef1d) | 1.902054 |
| AF146738 | centrosomal protein 19(Cep19) | 1.815362 |
| AF155196 | No data | 1.575313 |
| AF169390 | phosphodiesterase 6H(Pde6h) | 1.774068 |
| AF169636 | leukocyte immunoglobulin-like receptor, subfamily B (with TM and ITIM domains), member 3-like(Lilrb3l) | 3.098234 |
| AF173834 | calpain 3(Capn3) | 1.680331 |
| AF176023 | PR/SET domain 4(Prdm4) | 2.242755 |
| AF178689 | carbohydrate sulfotransferase 3(Chst3) | 1.70415 |
| AF187323 | cathepsin Q(Ctsq) | 2.106614 |
| AF190256 | phosphate cytidylyltransferase 1, choline, beta(Pcyt1b) | 2.363304 |
| AF201901 | interferon gamma receptor 1(Ifngr1) | 1.542342 |
| AF208125 | acyl-CoA synthetase bubblegum family member 1(Acsbg1) | 2.403616 |
| AF214647 | N-acylsphingosine amidohydrolase 1(Asah1) | 1.539499 |
| AF218575 | nibrin(Nbn) | 1.875441 |
| AF228043 | nuclear receptor coactivator 6(Ncoa6) | 1.721794 |
| AF228917 | zinc finger, DHHC-type containing 2(Zdhhc2) | 1.609594 |
| AF237778 | calcium/calmodulin-dependent protein kinase II alpha(Camk2a) | 1.638852 |
| AF273025 | solute carrier family 38, member 3(Slc38a3) | 2.373408 |
| AF276940 | ectonucleoside triphosphate diphosphohydrolase 2(Entpd2) | 2.0331 |
| AF281304 | potassium channel, two pore domain subfamily K, member 6(Kcnk6) | 2.029054 |
| AF288611 | RUN domain containing 3A(Rundc3a) | 2.840557 |
| AF303035 | PARP1 binding protein(Parpbp) | 2.933101 |
| AF308818 | KH-type splicing regulatory protein(Khsrp) | 1.694262 |
| AF311055 | thioredoxin 1(Txn1) | 2.848762 |
| AF333325 | HPS1, biogenesis of lysosomal organelles complex 3 subunit 1(Hps1) | 1.833169 |
| AF333986 | oxidation resistance 1(Oxr1) | 2.393664 |
| AF336113 | actin-binding Rho activating protein(Abra) | 2.216764 |
| AF345444 | potassium voltage-gated channel interacting protein 4(Kcnip4) | 2.743564 |
| AF368269 | cytochrome P450, family 2, subfamily t, polypeptide 1(Cyp2t1) | 1.620044 |
| AF393750 | BPI fold containing family A, member 1(Bpifa1) | 2.365618 |
| AF398465 | dihydropyrimidinase-like 3(Dpysl3) | 1.872371 |
| AF419333 | gamma-aminobutyric acid type A receptor theta subunit(Gabrq) | 1.792286 |
| AI059116 | No data | 1.626247 |
| AI102932 | No data | 1.930485 |
| AI104638 | No data | 2.595373 |
| AI105022 | No data | 1.646591 |
| AI113337 | No data | 1.994172 |
| AI230498 | No data | 1.568518 |
| AI230682 | No data | 2.036962 |
| AI231775 | No data | 1.560267 |
| AI385377 | No data | 3.220256 |
| AI406694 | No data | 2.313044 |
| AI500802 | No data | 2.504556 |
| AI535093 | No data | 1.900459 |
| AI599423 | No data | 1.514733 |
| AI602844 | No data | 1.652924 |
| AI717432 | No data | 1.677302 |
| AJ012482 | phosphatidylinositol-4,5-bisphosphate 3-kinase, catalytic subunit beta(Pik3cb) | 2.039374 |
| AT005481 | No data | 1.678314 |
| AW141757 | No data | 1.778821 |
| AW520335 | No data | 2.449793 |
| AW533345 | No data | 1.75804 |
| AW914284 | No data | 1.704328 |
| AW915815 | No data | 1.91521 |
| AW916410 | No data | 1.858779 |
| AW918157 | No data | 1.600146 |
| AW918457 | No data | 1.504258 |
| AW918850 | No data | 1.940086 |
| AW921253 | No data | 1.848394 |
| AY026068 | ras homolog family member A(Rhoa) | 2.033377 |
| AY030278 | Wnt inhibitory factor 1(Wif1) | 1.764736 |
| AY034383 | dynein light chain LC8-type 2(Dynll2) | 1.527297 |
| AY035403 | kinesin family member 6(Kif6) | 1.9322 |
| AY122322 | protein phosphatase 1, regulatory (inhibitor) subunit 14D(Ppp1r14d) | 2.374861 |
| BE100543 | No data | 2.356497 |
| BE113282 | No data | 2.311542 |
| BE113420 | No data | 1.660364 |
| BE118455 | No data | 1.925824 |
| BF390636 | No data | 1.846075 |
| BF405195 | No data | 1.559474 |
| BF406553 | No data | 1.625267 |
| BF411273 | No data | 1.537562 |
| BF418537 | No data | 1.791187 |
| BF420605 | No data | 1.918056 |
| BF522262 | No data | 2.816838 |
| BF522647 | No data | 1.689307 |
| BF525282 | No data | 2.164385 |
| BF543281 | No data | 1.772391 |
| BF543321 | No data | 1.81299 |
| BF546465 | No data | 2.021733 |
| BF553424 | No data | 1.799168 |
| BF561374 | No data | 1.753152 |
| BF563877 | No data | 2.224055 |
| BF564018 | No data | 1.608414 |
| BF565044 | No data | 1.715644 |
| BF565565 | No data | 1.534809 |
| BG374035 | No data | 1.848573 |
| BG664137 | No data | 1.654493 |
| BG665039 | No data | 1.996679 |
| BG666176 | No data | 2.409512 |
| BG668317 | No data | 1.503399 |
| BG668660 | No data | 1.902199 |
| BG671436 | No data | 2.520046 |
| BG671873 | No data | 2.019116 |
| BG673258 | No data | 2.563218 |
| BI282584 | No data | 2.135856 |
| BM383757 | No data | 1.584869 |
| BQ207374 | No data | 1.532423 |
| BQ211469 | No data | 1.62545 |
| BU671396 | No data | 1.554288 |
| BU671701 | No data | 2.802387 |
| D00729 | enoyl-CoA delta isomerase 1(Eci1) | 2.814065 |
| D14480 | calpain 8(Capn8) | 1.674437 |
| D21095 | chemokine (C-X-C motif) ligand 3(Cxcl3) | 1.92518 |
| D30040 | AKT serine/threonine kinase 1(Akt1) | 1.680116 |
| D38261 | protein phosphatase 2, regulatory subunit B, gamma(Ppp2r2c) | 1.79365 |
| D85435 | protein kinase C, delta binding protein(Prkcdbp) | 1.78163 |
| D87950 | ubiquilin 1(Ubqln1) | 1.763773 |
| J00744 | No data | 1.516659 |
| L04760 | tripartite motif-containing 23(Trim23) | 2.018875 |
| L07315 | dipeptidase 1 (renal)(Dpep1) | 1.601127 |
| L13606 | myosin heavy chain 2(Myh2) | 1.787821 |
| L15618 | casein kinase 2 alpha 1(Csnk2a1) | 1.7187 |
| L28801 | general transcription factor IIIC subunit 1(Gtf3c1) | 1.516161 |
| L29427 | No data | 2.009211 |
| L35921 | G protein subunit gamma 8(Gng8) | 1.713815 |
| M13011 | No data | 1.614206 |
| M15883 | clathrin, light chain B(Cltb) | 1.806758 |
| M21759 | filaggrin(Flg) | 1.813956 |
| M22030 | No data | 1.780357 |
| M32062 | Fc fragment of IgG, low affinity IIa, receptor(Fcgr2a) | 1.60304 |
| M33821 | gamma-glutamyltransferase 1(Ggt1) | 2.263792 |
| M91450 | No data | 1.564184 |
| M96626 | ATPase plasma membrane Ca2+ transporting 3(Atp2b3) | 2.172804 |
| M97255 | trefoil factor 2(Tff2) | 1.647276 |
| NM_012519 | calcium/calmodulin-dependent protein kinase II delta(Camk2d) | 1.505653 |
| NM_012520 | catalase(Cat) | 2.274713 |
| NM_012526 | chromogranin B(Chgb) | 2.018038 |
| NM_012528 | cholinergic receptor nicotinic beta 1 subunit(Chrnb1) | 1.680012 |
| NM_012531 | catechol-O-methyltransferase(Comt) | 2.231328 |
| NM_012543 | D-box binding PAR bZIP transcription factor(Dbp) | 1.532421 |
| NM_012549 | endothelin 2(Edn2) | 1.830311 |
| NM_012565 | glucokinase(Gck) | 3.003472 |
| NM_012572 | glutamate ionotropic receptor kainate type subunit 4(Grik4) | 1.960676 |
| NM_012594 | lactalbumin, alpha(Lalba) | 2.35724 |
| NM_012596 | leptin receptor(Lepr) | 1.572461 |
| NM_012600 | malic enzyme 1(Me1) | 1.705429 |
| NM_012612 | natriuretic peptide A(Nppa) | 1.752513 |
| NM_012619 | phenylalanine hydroxylase(Pah) | 1.861168 |
| NM_012653 | solute carrier family 9 member A2(Slc9a2) | 1.680892 |
| NM_012663 | vesicle-associated membrane protein 2(Vamp2) | 1.751666 |
| NM_012667 | tachykinin receptor 1(Tacr1) | 2.182398 |
| NM_012695 | sulfotransferase family 2A, dehydroepiandrosterone (DHEA)-preferring, member 6(Sult2a6) | 1.605457 |
| NM_012730 | cytochrome P450, family 2, subfamily d, polypeptide 2(Cyp2d2) | 1.829295 |
| NM_012734 | hexokinase 1(Hk1) | 1.709348 |
| NM_012788 | discs large MAGUK scaffold protein 1(Dlg1) | 1.700899 |
| NM_012850 | growth hormone releasing hormone receptor(Ghrhr) | 1.557048 |
| NM_012876 | ribosomal protein S29(Rps29) | 2.638145 |
| NM_012904 | annexin A1(Anxa1) | 1.502437 |
| NM_012908 | Fas ligand(Faslg) | 2.142956 |
| NM_012910 | arrestin, beta 1(Arrb1) | 1.672619 |
| NM_012941 | cytochrome P450, family 51(Cyp51) | 1.657199 |
| NM_012963 | high mobility group box 1(Hmgb1) | 1.666837 |
| NM_012978 | luteinizing hormone/choriogonadotropin receptor(Lhcgr) | 3.050579 |
| NM_012980 | matrix metallopeptidase 11(Mmp11) | 2.630092 |
| NM_012993 | nardilysin convertase(Nrdc) | 2.298098 |
| NM_013005 | phosphoinositide-3-kinase regulatory subunit 1(Pik3r1) | 2.802073 |
| NM_013008 | POU class 1 homeobox 1(Pou1f1) | 1.552156 |
| NM_013030 | solute carrier family 34 member 1(Slc34a1) | 1.569844 |
| NM_013183 | meprin A subunit beta(Mep1b) | 1.901411 |
| NM_013224 | ribosomal protein S26(Rps26) | 1.620632 |
| NM_013226 | ribosomal protein L32(Rpl32) | 2.284438 |
| NM_013413 | relaxin 1(Rln1) | 1.971464 |
| NM_017015 | glucuronidase, beta(Gusb) | 1.8737 |
| NM_017042 | protein phosphatase 3 catalytic subunit beta(Ppp3cb) | 1.989406 |
| NM_017137 | chloride channel, voltage-sensitive 2(Clcn2) | 1.867684 |
| NM_017144 | troponin I3, cardiac type(Tnni3) | 1.571609 |
| NM_017169 | peroxiredoxin 2(Prdx2) | 2.126629 |
| NM_017193 | aminoadipate aminotransferase(Aadat) | 2.633207 |
| NM_017251 | gap junction protein, beta 1(Gjb1) | 1.77218 |
| NM_017256 | transforming growth factor beta receptor 3(Tgfbr3) | 1.864647 |
| NM_017264 | proteasome activator subunit 1(Psme1) | 1.819425 |
| NM_017279 | proteasome subunit alpha 2(Psma2) | 1.725385 |
| NM_017348 | solute carrier family 6 member 8(Slc6a8) | 1.628447 |
| NM_019137 | early growth response 4(Egr4) | 1.894886 |
| NM_019151 | myostatin(Mstn) | 2.351448 |
| NM_019161 | cadherin 22(Cdh22) | 2.206278 |
| NM_019165 | interleukin 18(Il18) | 1.52836 |
| NM_019170 | carbonyl reductase 1(Cbr1) | 1.714897 |
| NM_019179 | thymidylate synthetase(Tyms) | 1.837275 |
| NM_019198 | fibroblast growth factor 17(Fgf17) | 2.35561 |
| NM_019216 | growth differentiation factor 15(Gdf15) | 1.772315 |
| NM_019266 | sodium voltage-gated channel alpha subunit 8(Scn8a) | 2.082371 |
| NM_019272 | ssemaphorin 4F(Sema4f) | 1.685348 |
| NM_019277 | exocyst complex component 6(Exoc6) | 1.957294 |
| NM_019291 | carbonic anhydrase 2(Car2) | 2.185707 |
| NM_019323 | mast cell protease 9(Mcpt9) | 1.647125 |
| NM_019340 | regulator of G-protein signaling 3(Rgs3) | 1.639602 |
| NM_019353 | thyroid peroxidase(Tpo) | 1.757899 |
| NM_019376 | tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, gamma(Ywhag) | 1.753748 |
| NM_019904 | galectin 1(Lgals1) | 1.747295 |
| NM_020082 | ribonuclease A family member 4(Rnase4) | 1.5499 |
| NM_021581 | prolyl 3-hydroxylase family member 4(P3h4) | 1.874377 |
| NM_021585 | mast cell immunoglobulin-like receptor 1(Milr1) | 1.878356 |
| NM_021598 | mast cell protease 8(Mcpt8) | 2.72646 |
| NM_021671 | transmembrane protein 33(Tmem33) | 1.613433 |
| NM_021684 | adenylate cyclase 10 (soluble)(Adcy10) | 1.83554 |
| NM_021697 | potassium voltage-gated channel modifier subfamily V member 1(Kcnv1) | 1.756715 |
| NM_021701 | protein phosphatase 3, regulatory subunit B, beta(Ppp3r2) | 1.764523 |
| NM_021748 | N-ethylmaleimide sensitive factor, vesicle fusing ATPase(Nsf) | 1.720353 |
| NM_021772 | cyclin-dependent kinase-like 3(Cdkl3) | 1.589505 |
| NM_021849 | RFNG O-fucosylpeptide 3-beta-N-acetylglucosaminyltransferase(Rfng) | 1.75421 |
| NM_021859 | megakaryocyte-associated tyrosine kinase(Matk) | 1.846828 |
| NM_021997 | CAP-GLY domain containing linker protein 2(Clip2) | 1.829356 |
| NM_022203 | No data | 1.914261 |
| NM_022210 | MYC associated factor X(Max) | 1.814325 |
| NM_022254 | G protein-coupled receptor 85(Gpr85) | 1.800299 |
| NM_022282 | discs large MAGUK scaffold protein 2(Dlg2) | 1.768177 |
| NM_022284 | guanylate cyclase activator 2B(Guca2b) | 2.039829 |
| NM_022296 | xylosyltransferase 2(Xylt2) | 2.198775 |
| NM_022297 | dimethylarginine dimethylaminohydrolase 1(Ddah1) | 1.685947 |
| NM_022509 | survival of motor neuron 1, telomeric(Smn1) | 1.906114 |
| NM_022538 | phospholipid phosphatase 1(Plpp1) | 1.909921 |
| NM_022600 | adenylate cyclase 5(Adcy5) | 1.520435 |
| NM_022613 | No data | 1.929265 |
| NM_022623 | frizzled class receptor 4(Fzd4) | 2.230565 |
| NM_022625 | tropic 1808(Tpc1808) | 1.653187 |
| NM_022638 | transient receptor potential cation channel, subfamily C, member 2, pseudogene(Trpc2) | 1.663797 |
| NM_022639 | cholinergic receptor nicotinic alpha 10 subunit(Chrna10) | 1.732239 |
| NM_022686 | histone cluster 1, H4b(Hist1h4b) | 2.369333 |
| NM_022704 | mannose-binding lectin (protein C) 2(Mbl2) | 1.568571 |
| NM_022850 | dipeptidyl peptidase like 6(Dpp6) | 2.262092 |
| NM_022859 | cysteine-rich secretory protein 1(Crisp1) | 1.609319 |
| NM_022929 | potassium voltage-gated channel interacting protein 1(Kcnip1) | 1.663328 |
| NM_022947 | ClpB homolog, mitochondrial AAA ATPase chaperonin(Clpb) | 1.811601 |
| NM_022954 | FAT atypical cadherin 2(Fat2) | 2.727616 |
| NM_023102 | casein kinase 1, gamma 2(Csnk1g2) | 1.702925 |
| NM_023963 | caudal type homeo box 2(Cdx2) | 1.683684 |
| NM_023969 | lysophosphatidic acid receptor 3(Lpar3) | 1.63442 |
| NM_023973 | indoleamine 2,3-dioxygenase 1(Ido1) | 1.519611 |
| NM_023981 | colony stimulating factor 1(Csf1) | 1.538487 |
| NM_023994 | taste receptor, type 2, member 118(Tas2r118) | 1.695958 |
| NM_023995 | taste receptor, type 2, member 107(Tas2r107) | 2.24126 |
| NM_024127 | growth arrest and DNA-damage-inducible, alpha(Gadd45a) | 2.152067 |
| NM_024381 | glycerol kinase(Gk) | 1.811678 |
| NM_024487 | GrpE-like 1, mitochondrial(Grpel1) | 1.943116 |
| NM_024489 | zinc finger and BTB domain containing 10(Zbtb10) | 2.056393 |
| NM_030826 | glutathione peroxidase 1(Gpx1) | 1.652839 |
| NM_030837 | kidney specific organic anion transporter(Slc21a4) | 2.998452 |
| NM_030843 | syntaxin binding protein 5(Stxbp5) | 2.170744 |
| NM_030860 | myocyte enhancer factor 2D(Mef2d) | 2.048593 |
| NM_030868 | nephroblastoma overexpressed(Nov) | 1.600397 |
| NM_030875 | sodium voltage-gated channel alpha subunit 1(Scn1a) | 1.622929 |
| NM_031000 | aldo-keto reductase family 1 member A1(Akr1a1) | 1.567229 |
| NM_031021 | casein kinase 2 beta(Csnk2b) | 1.917293 |
| NM_031036 | G protein subunit alpha q(Gnaq) | 1.918023 |
| NM_031059 | msh homeobox 1(Msx1) | 1.99165 |
| NM_031065 | ribosomal protein L10A(Rpl10a) | 2.300643 |
| NM_031085 | protein kinase C, eta(Prkch) | 2.372214 |
| NM_031093 | RAS like proto-oncogene A(Rala) | 2.39697 |
| NM_031120 | signal sequence receptor, gamma(Ssr3) | 1.674383 |
| NM_031129 | transcription elongation factor B subunit 2(Tceb2) | 2.331149 |
| NM_031130 | nuclear receptor subfamily 2, group F, member 1(Nr2f1) | 1.84239 |
| NM_031133 | thrombopoietin(Thpo) | 1.757085 |
| NM_031136 | thymosin beta 4, X-linked(Tmsb4x) | 2.204979 |
| NM_031142 | double C2 domain beta(Doc2b) | 2.438734 |
| NM_031240 | cysteine-rich secretory protein 2(Crisp2) | 2.969829 |
| NM_031352 | drebrin-like(Dbnl) | 1.689691 |
| NM_031360 | sphingomyelin phosphodiesterase 2(Smpd2) | 1.509688 |
| NM_031537 | RoBo-1(LOC24906) | 2.382064 |
| NM_031570 | ribosomal protein S7(Rps7) | 1.720451 |
| NM_031575 | AKT serine/threonine kinase 3(Akt3) | 1.827906 |
| NM_031601 | calcium voltage-gated channel subunit alpha1 G(Cacna1g) | 2.076481 |
| NM_031603 | tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, epsilon(Ywhae) | 1.516651 |
| NM_031604 | ATPase H+ transporting V0 subunit a1(Atp6v0a1) | 2.734505 |
| NM_031621 | SH2B adaptor protein 3(Sh2b3) | 1.87789 |
| NM_031624 | immunoglobulin (CD79A) binding protein 1(Igbp1) | 1.647906 |
| NM_031627 | nuclear receptor subfamily 1, group H, member 3(Nr1h3) | 1.513038 |
| NM_031643 | mitogen activated protein kinase kinase 1(Map2k1) | 2.033606 |
| NM_031728 | synaptosomal-associated protein 91(Snap91) | 1.989374 |
| NM_031729 | protein phosphatase 5, catalytic subunit(Ppp5c) | 1.7209 |
| NM_031762 | cyclin-dependent kinase inhibitor 1B(Cdkn1b) | 1.807938 |
| NM_031781 | amyloid beta precursor protein binding family A member 3(Apba3) | 1.770669 |
| NM_031789 | nuclear factor, erythroid 2-like 2(Nfe2l2) | 2.362231 |
| NM_031821 | polo-like kinase 2(Plk2) | 2.022837 |
| NM_031823 | wolframin ER transmembrane glycoprotein(Wfs1) | 2.469074 |
| NM_031832 | galectin 3(Lgals3) | 1.597594 |
| NM_031836 | vascular endothelial growth factor A(Vegfa) | 1.578976 |
| NM_031974 | clathrin, light chain A(Clta) | 1.664435 |
| NM_032079 | DnaJ heat shock protein family (Hsp40) member A2(Dnaja2) | 1.580883 |
| NM_032082 | hydroxyacid oxidase 2(Hao2) | 1.813424 |
| NM_033376 | potassium two pore domain channel subfamily K member 3(Kcnk3) | 1.540317 |
| NM_053323 | delta(4)-desaturase, sphingolipid 1(Degs1) | 1.581652 |
| NM_053716 | fructose-bisphosphatase 2(Fbp2) | 1.901898 |
| NM_057098 | transcription elongation factor A2(Tcea2) | 2.570171 |
| NM_057099 | proteasome subunit beta 6(Psmb6) | 1.767274 |
| NM_078622 | phosphate cytidylyltransferase 1, choline, alpha(Pcyt1a) | 1.676003 |
| NM_080900 | actin filament associated protein 1(Afap1) | 1.565316 |
| NM_130420 | tripartite motif-containing 9(Trim9) | 2.025582 |
| NM_130421 | lymphocyte cytosolic protein 2(Lcp2) | 1.912839 |
| NM_130753 | fibroblast growth factor 19(Fgf19) | 1.524216 |
| NM_133309 | calpain 8(Capn8) | 2.126877 |
| NM_133321 | potassium voltage-gated channel subfamily J member 15(Kcnj15) | 2.331074 |
| NM_133383 | serine carboxypeptidase 1(Scpep1) | 1.565457 |
| NM_133392 | serine/threonine kinase 17b(Stk17b) | 2.265261 |
| NM_133406 | 1-acylglycerol-3-phosphate O-acyltransferase 4(Agpat4) | 1.599979 |
| NM_133612 | No data | 1.653442 |
| NM_134376 | calsyntenin 3(Clstn3) | 1.500037 |
| NM_134462 | ATPase secretory pathway Ca2+ transporting 2(Atp2c2) | 2.49921 |
| NM_134465 | cytokine receptor-like factor 2(Crlf2) | 2.519908 |
| NM_138710 | DAB2 interacting protein(Dab2ip) | 1.61244 |
| NM_139193 | prolactin releasing hormone receptor(Prlhr) | 1.542231 |
| NM_144741 | resistin(Retn) | 1.849312 |
| NM_144742 | ATPase phospholipid transporting 11A(Atp11a) | 2.002968 |
| NM_147207 | ischemia related factor vof-16(Vof16) | 1.623283 |
| U03390 | receptor for activated C kinase 1(Rack1) | 1.944424 |
| U05593 | Cd80 molecule(Cd80) | 2.654472 |
| U06434 | C-C motif chemokine ligand 4(Ccl4) | 1.819756 |
| U14746 | von Hippel-Lindau tumor suppressor(Vhl) | 1.543271 |
| U17604 | reticulon 1(Rtn1) | 1.825625 |
| U18771 | RAB26, member RAS oncogene family(Rab26) | 1.820437 |
| U39208 | cytochrome P450, family 4, subfamily f, polypeptide 6(Cyp4f6) | 1.645854 |
| U48828 | No data | 1.727898 |
| U51583 | zinc finger E-box binding homeobox 1(Zeb1) | 1.51051 |
| U53475 | RAB8B, member RAS oncogene family(Rab8b) | 1.913337 |
| U57063 | granzyme F(Gzmf) | 1.980644 |
| U57391 | SH2B adaptor protein 1(Sh2b1) | 2.074809 |
| U72353 | lamin B1(Lmnb1) | 2.074554 |
| U92564 | zinc finger protein 423(Zfp423) | 2.090788 |
| U93851 | cyclic nucleotide gated channel alpha 1(Cnga1) | 1.506566 |
| U94856 | paraoxonase 1(Pon1) | 1.546413 |
| X51992 | gamma-aminobutyric acid type A receptor alpha 5 subunit(Gabra5) | 1.648049 |
| X52952 | Moloney sarcoma oncogene(Mos) | 1.892727 |
| X56190 | No data | 1.501611 |
| X63281 | No data | 2.264276 |
| X69029 | No data | 1.553785 |
| X80671 | olfactory receptor 1271(Olr1271) | 1.74883 |
| X81193 | cysteine and glycine rich protein 3(Csrp3) | 1.681766 |
| X89603 | metallothionein 3(Mt3) | 2.362925 |
| X89962 | cold shock domain containing C2(Csdc2) | 2.244 |
| X96790 | glutamate metabotropic receptor 7(Grm7) | 1.589366 |
| X98490 | replication protein A2(Rpa2) | 1.518939 |
| Z75029 | heat shock 70kD protein 1B (mapped)(Hspa1b) | 1.844737 |
Appendix B. List of Genes Down-Regulated by PE Effect
| Gene Id | Gene | Z-Score |
| AA944170 | S/D | −1.506199 |
| AA944489 | No data | −1.530233 |
| AA996993 | No data | −2.983268 |
| AB011529 | cadherin, EGF LAG seven-pass G-type receptor 2(Celsr2) | −1.74635 |
| AB011679 | tubulin, beta 5 class I(Tubb5) | −2.325114 |
| AB020504 | No data | −2.211124 |
| AB020757 | chymotrypsin-like(Ctrl) | −2.059928 |
| AB025017 | No data | −2.29894 |
| AB033418 | No data | −2.907947 |
| AB047540 | isocitrate dehydrogenase 3 (NAD+) beta(Idh3B) | −1.980831 |
| AB067445 | integrin alpha 2(Itga2) | −1.977614 |
| AF000944 | general transcription factor IIA, 2(Gtf2a2) | −2.614055 |
| AF006664 | NK2 homeobox 5(Nkx2-5) | −1.571779 |
| AF012714 | multiple inositol-polyphosphate phosphatase 1(Minpp1) | −1.517925 |
| AF015953 | aryl hydrocarbon receptor nuclear translocator-like(Arntl) | −1.67005 |
| AF016387 | retinoid X receptor gamma(Rxrg) | −1.847981 |
| AF021936 | CDC42 binding protein kinase beta(Cdc42bpb) | −2.024961 |
| AF026505 | sorbin and SH3 domain containing 2(Sorbs2) | −1.84203 |
| AF032120 | GIPC PDZ domain containing family, member 1(Gipc1) | −1.832343 |
| AF037071 | nitric oxide synthase 1 adaptor protein(Nos1ap) | −2.242322 |
| AF039584 | CD55 molecule, decay accelerating factor for complement(Cd55) | −1.707524 |
| AF053093 | No data | −1.976489 |
| AF053097 | No data | −1.603483 |
| AF084544 | versican(Vcan) | −1.711944 |
| AF092090 | polyamine modulated factor 1 binding protein 1(Pmfbp1) | −1.629489 |
| AF123651 | spermatogenesis associated 2(Spata2) | −1.791841 |
| AF151710 | No data | −1.874231 |
| AF170284 | gap junction protein, beta 6(Gjb6) | −2.501274 |
| AF193757 | N-terminal EF-hand calcium binding protein 2(Necab2) | −1.515331 |
| AF200359 | UDP-glucose glycoprotein glucosyltransferase 1(Uggt1) | −2.250208 |
| AF243515 | BCL2/adenovirus E1B interacting protein 3(Bnip3) | −1.627503 |
| AF247450 | hyperpolarization-activated cyclic nucleotide-gated potassium channel 1(Hcn1) | −1.51455 |
| AF277901 | zinc finger protein 483(Zfp483) | −1.798414 |
| AF302047 | CXADR-like membrane protein(Clmp) | −2.140061 |
| AF304429 | mitochondrial pyruvate carrier 1(Mpc1) | −2.257792 |
| AF307852 | ARP3 actin related protein 3 homolog(Actr3) | −1.95466 |
| AF323615 | phospholipase C, epsilon 1(Plce1) | −4.175461 |
| AF329856 | phosphodiesterase 1C(Pde1c) | −1.504374 |
| AF347935 | interleukin 11(Il11) | −1.674463 |
| AF361476 | Cbp/p300-interacting transactivator, with Glu/Asp-rich carboxy-terminal domain, 2(Cited2) | −1.710103 |
| AF367467 | selenoprotein S(Selenos) | −2.382484 |
| AF379608 | doublesex and mab-3 related transcription factor 1(Dmrt1) | −1.683129 |
| AF400662 | calcium voltage-gated channel auxiliary subunit alpha2delta 1(Cacna2d1) | −1.543195 |
| AF537333 | aurora kinase A(Aurka) | −1.661243 |
| AI059699 | No data | −1.842174 |
| AI136886 | No data | −1.650746 |
| AI229387 | No data | −1.511607 |
| AI598392 | No data | −1.725522 |
| AJ006855 | synaptojanin 1(Synj1) | −1.727807 |
| AJ131196 | No data | −1.769069 |
| AJ250280 | potassium voltage-gated channel subfamily H member 5(Kcnh5) | −1.520944 |
| AJ303374 | ATP binding cassette subfamily G member 1(Abcg1) | −1.609867 |
| AJ409332 | TIMP metallopeptidase inhibitor 2(Timp2) | −2.3002 |
| AW141281 | No data | −2.051067 |
| AW141994 | No data | −1.692865 |
| AW434257 | No data | −1.789087 |
| AW915590 | No data | −1.557293 |
| AW916146 | No data | −1.959788 |
| AW916635 | No data | −1.746735 |
| AW917632 | No data | −1.549931 |
| AW918748 | No data | −2.166549 |
| AW918775 | No data | −1.649553 |
| AW919008 | No data | −1.692303 |
| AY011335 | No data | −1.711045 |
| AY014898 | inositol polyphosphate multikinase(Ipmk) | −2.342213 |
| AY026512 | dynein light chain roadblock-type 1(Dynlrb1) | −2.119183 |
| BF284301 | No data | −2.121369 |
| BF287132 | No data | −2.068833 |
| BF392368 | No data | −2.212956 |
| BF395339 | No data | −1.928218 |
| BF399588 | No data | −2.70933 |
| BF414052 | No data | −2.155561 |
| BF416262 | No data | −1.546722 |
| BF524417 | No data | −2.049708 |
| BF549771 | No data | −1.690628 |
| BF555199 | No data | −1.523037 |
| BF559446 | No data | −2.697068 |
| BF560218 | No data | −2.252126 |
| BF566173 | No data | −2.261156 |
| BF567456 | No data | −1.898923 |
| BG663098 | No data | −1.5361 |
| BG664103 | No data | −2.448298 |
| BG666505 | No data | −1.572982 |
| BG667467 | No data | −2.508352 |
| BG671325 | No data | −1.655659 |
| BI278738 | No data | −2.62235 |
| BI295378 | No data | −2.278025 |
| BI296499 | No data | −2.01441 |
| BM386847 | No data | −2.077829 |
| BQ194714 | No data | −1.88522 |
| BQ205274 | NADH:ubiquinone oxidoreductase subunit A12(Ndufa12) | −2.38962 |
| BQ208291 | No data | −1.636124 |
| BU670896 | No data | −3.513474 |
| BU671010 | No data | −2.556612 |
| BU671095 | No data | −2.578096 |
| BU671151 | No data | −1.744558 |
| D00920 | seminal vesicle secretory protein 3A(Svs3a) | −2.000729 |
| D14048 | heterogeneous nuclear ribonucleoprotein U(Hnrnpu) | −2.261341 |
| D16465 | adenylate cyclase activating polypeptide 1 receptor type 1(Adcyap1r1) | −1.966142 |
| D16479 | hydroxyacyl-CoA dehydrogenase/3-ketoacyl-CoA thiolase/enoyl-CoA hydratase (trifunctional protein), beta subunit(Hadhb) | −1.944151 |
| D26178 | intestinal cell kinase(Ick) | −2.540983 |
| D63772 | solute carrier family 1 member 1(Slc1a1) | −1.665296 |
| J02585 | stearoyl-CoA desaturase(Scd) | −2.299181 |
| J02868 | cytochrome P450, family 2, subfamily d, polypeptide 3(Cyp2d3) | −1.688779 |
| J04628 | 3-hydroxyisobutyrate dehydrogenase(Hibadh) | −1.717486 |
| L02121 | cyclin-dependent kinase 5(Cdk5) | −2.181096 |
| L02315 | calcium voltage-gated channel auxiliary subunit beta 4(Cacnb4) | −1.528497 |
| L10362 | synaptic vesicle glycoprotein 2b(Sv2b) | −1.569646 |
| L12407 | No data | −1.867835 |
| L22654 | gamma-2a immunoglobulin heavy chain(IgG-2a) | −1.709121 |
| L27487 | calcitonin receptor-like(Calcrl) | −1.970351 |
| M11794 | No data | −1.536746 |
| M15202 | No data | −1.947036 |
| M15327 | alcohol dehydrogenase 1 (class I)(Adh1) | −2.079242 |
| M15481 | insulin-like growth factor 1(Igf1) | −2.298139 |
| M16409 | cholinergic receptor, muscarinic 4(Chrm4) | −1.743017 |
| M18841 | No data | −1.998377 |
| M29295 | small nuclear ribonucleoprotein polypeptides B and B1(Snrpb) | −1.675114 |
| M30692 | No data | −2.027642 |
| M31322 | amyloid beta precursor like protein 2(Aplp2) | −1.814887 |
| M57705 | thyroid peroxidase(Tpo) | −1.513088 |
| M61142 | thimet oligopeptidase 1(Thop1) | −1.696997 |
| M63837 | platelet derived growth factor receptor alpha(Pdgfra) | −1.822324 |
| M64381 | olfactory receptor 1082(Olr1082) | −1.761637 |
| M76733 | glutathione peroxidase 6(Gpx6) | −1.971279 |
| M83143 | ST6 beta-galactoside alpha-2,6-sialyltransferase 1(St6gal1) | −1.602127 |
| M83209 | BPI fold containing family A, member 2(Bpifa2) | −2.386439 |
| M83210 | BPI fold containing family A, member 2F(Bpifa2f) | −1.615422 |
| M83745 | proprotein convertase subtilisin/kexin type 1(Pcsk1) | −1.961893 |
| M94043 | RAB38, member RAS oncogene family(Rab38) | −1.660598 |
| NM_012490 | acrosin(Acr) | −1.761651 |
| NM_012502 | androgen receptor(Ar) | −2.367953 |
| NM_012542 | cytochrome P450, family 2, subfamily a, polypeptide 3(Cyp2a3) | −1.563632 |
| NM_012589 | interleukin 6(Il6) | −2.279895 |
| NM_012595 | lactate dehydrogenase B(Ldhb) | −1.56024 |
| NM_012621 | 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 1(Pfkfb1) | −2.130949 |
| NM_012635 | protease, serine 1(Prss1) | −1.938556 |
| NM_012657 | serine (or cysteine) proteinase inhibitor, clade A, member 3C(Serpina3c) | −1.73983 |
| NM_012679 | No data | −1.660105 |
| NM_012681 | transthyretin(Ttr) | −1.632171 |
| NM_012682 | uncoupling protein 1(Ucp1) | −1.956669 |
| NM_012703 | thyroid hormone responsive(Thrsp) | −2.132157 |
| NM_012706 | gastrin releasing peptide receptor(Grpr) | −2.32707 |
| NM_012740 | tyrosine hydroxylase(Th) | −1.687967 |
| NM_012777 | apolipoprotein D(Apod) | −2.293886 |
| NM_012805 | retinoid X receptor alpha(Rxra) | −1.788029 |
| NM_012825 | aquaporin 4(Aqp4) | −1.738027 |
| NM_012826 | alpha-2-glycoprotein 1, zinc-binding(Azgp1) | −1.917567 |
| NM_012841 | DCC netrin 1 receptor(Dcc) | −2.060894 |
| NM_012843 | epithelial membrane protein 1(Emp1) | −1.524748 |
| NM_012922 | caspase 3(Casp3) | −1.730399 |
| NM_012940 | cytochrome P450, family 1, subfamily b, polypeptide 1(Cyp1b1) | −1.825438 |
| NM_012949 | enolase 3(Eno3) | −2.250452 |
| NM_012951 | fibroblast growth factor 10(Fgf10) | −1.581501 |
| NM_012967 | intercellular adhesion molecule 1(Icam1) | −1.527629 |
| NM_012971 | potassium voltage-gated channel subfamily A member 4(Kcna4) | −2.126831 |
| NM_013023 | S-antigen visual arrestin(Sag) | −1.921779 |
| NM_013054 | No data | −1.571264 |
| NM_013095 | SMAD family member 3(Smad3) | −1.508985 |
| NM_013159 | insulin degrading enzyme(Ide) | −1.663445 |
| NM_013161 | pancreatic lipase(Pnlip) | −2.292426 |
| NM_013167 | uncoupling protein 3(Ucp3) | −2.046956 |
| NM_013168 | hydroxymethylbilane synthase(Hmbs) | −4.18491 |
| NM_013192 | potassium voltage-gated channel subfamily J member 6(Kcnj6) | −1.544981 |
| NM_016998 | carboxypeptidase A1(Cpa1) | −1.72664 |
| NM_017006 | glucose-6-phosphate dehydrogenase(G6pd) | −2.105296 |
| NM_017011 | glutamate receptor, metabotropic 1(Grm1) | −1.945417 |
| NM_017014 | glutathione S-transferase mu 1(Gstm1) | −2.433956 |
| NM_017019 | interleukin 1 alpha(Il1a) | −1.572188 |
| NM_017069 | gamma-aminobutyric acid type A receptor alpha3 subunit(Gabra3) | −3.176886 |
| NM_017075 | acetyl-CoA acetyltransferase 1(Acat1) | −1.693916 |
| NM_017084 | glycine N-methyltransferase(Gnmt) | −1.587023 |
| NM_017085 | cytochrome P450, family 19, subfamily a, polypeptide 1(Cyp19a1) | −3.202593 |
| NM_017093 | AKT serine/threonine kinase 2(Akt2) | −1.708931 |
| NM_017101 | peptidylprolyl isomerase A (cyclophilin A)(Ppia) | −1.962416 |
| NM_017106 | chloride voltage-gated channel 5(Clcn5) | −2.303484 |
| NM_017120 | casein beta(Csn2) | −1.753558 |
| NM_017156 | cytochrome P450, family 2, subfamily b, polypeptide 12(Cyp2b12) | −1.675282 |
| NM_017158 | cytochrome P450, family 2, subfamily c, polypeptide 7(Cyp2c7) | −1.9999 |
| NM_017177 | choline kinase beta(Chkb) | −1.598296 |
| NM_017179 | UNC homeobox(Uncx) | −1.985105 |
| NM_017190 | myelin-associated glycoprotein(Mag) | −1.957495 |
| NM_017197 | CUGBP, Elav-like family member 2(Celf2) | −2.358172 |
| NM_017228 | atrophin 1(Atn1) | −3.12621 |
| NM_017265 | hydroxy-delta-5-steroid dehydrogenase, 3 beta- and steroid delta-isomerase 6(Hsd3b6) | −1.512438 |
| NM_017285 | proteasome subunit beta 3(Psmb3) | −2.192897 |
| NM_017291 | gamma-aminobutyric acid type A receptor rho 1 subunit(Gabrr1) | −3.620853 |
| NM_017292 | gamma-aminobutyric acid type A receptor rho 2 subunit(Gabrr2) | −1.576548 |
| NM_017326 | calmodulin 2(Calm2) | −2.141575 |
| NM_017337 | phosphodiesterase 3A(Pde3a) | −1.72263 |
| NM_017339 | ISL LIM homeobox 1(Isl1) | −2.983281 |
| NM_017343 | myosin light chain 12B(Myl12b) | −1.748003 |
| NM_019182 | ring finger protein 4(Rnf4) | −1.852677 |
| NM_019190 | CD46 molecule(Cd46) | −1.945158 |
| NM_019211 | RAS guanyl releasing protein 1(Rasgrp1) | −1.760087 |
| NM_019241 | gap junction protein, beta 5(Gjb5) | −1.748274 |
| NM_019248 | neurotrophic receptor tyrosine kinase 3(Ntrk3) | −1.880549 |
| NM_019255 | calcium voltage-gated channel auxiliary subunit gamma 1(Cacng1) | −1.569839 |
| NM_019261 | killer cell lectin-like receptor subfamily C, member 2(Klrc2) | −2.129231 |
| NM_019270 | potassium voltage-gated channel subfamily A member 3(Kcna3) | −2.943964 |
| NM_019278 | regulated endocrine-specific protein 18(Resp18) | −2.015828 |
| NM_019285 | adenylate cyclase 4(Adcy4) | −1.51557 |
| NM_019299 | clathrin heavy chain(Cltc) | −1.670579 |
| NM_019348 | somatostatin receptor 2(Sstr2) | −1.921952 |
| NM_019354 | uncoupling protein 2(Ucp2) | −2.271891 |
| NM_019363 | aldehyde oxidase 1(Aox1) | −1.50887 |
| NM_019620 | zinc finger protein 386 (Kruppel-like)(Zfp386) | −2.6169 |
| NM_019621 | discs large MAGUK scaffold protein 4(Dlg4) | −1.768061 |
| NM_019907 | CXXC repeat containing interactor of PDZ3 domain(Cript) | −1.965978 |
| NM_020074 | serglycin(Srgn) | −1.792037 |
| NM_020076 | 3-hydroxyanthranilate 3,4-dioxygenase(Haao) | −1.647168 |
| NM_020096 | interferon-induced protein with tetratricopeptide repeats 1(Ifit1) | −1.569029 |
| NM_020301 | ADAM metallopeptidase domain 7(Adam7) | −1.648235 |
| NM_020542 | C-C motif chemokine receptor 1(Ccr1) | −1.713195 |
| NM_021580 | prolactin family 8, subfamily a, member 4(Prl8a4) | −2.141944 |
| NM_021659 | synaptotagmin 7(Syt7) | −1.795637 |
| NM_021672 | growth differentiation factor 9(Gdf9) | −1.566303 |
| NM_021676 | SH3 and multiple ankyrin repeat domains 3(Shank3) | −2.237337 |
| NM_021703 | A-kinase anchoring protein 14(Akap14) | −2.557232 |
| NM_021742 | nuclear receptor subfamily 5, group A, member 2(Nr5a2) | −1.603742 |
| NM_021769 | sulfotransferase family 1D, member 1(Sult1d1) | −1.883171 |
| NM_022178 | myosin VA(Myo5a) | −2.420753 |
| NM_022196 | leukemia inhibitory factor(Lif) | −1.875411 |
| NM_022202 | glutamate metabotropic receptor 8(Grm8) | −1.511956 |
| NM_022204 | exocyst complex component 5(Exoc5) | −1.572194 |
| NM_022214 | C-X-C motif chemokine ligand 6(Cxcl6) | −2.525867 |
| NM_022241 | prostaglandin D2 receptor(Ptgdrl) | −1.557255 |
| NM_022261 | B-box and SPRY domain containing(Bspry) | −1.56046 |
| NM_022278 | glutaredoxin(Glrx) | −2.208115 |
| NM_022283 | No data | −2.543935 |
| NM_022386 | MAF bZIP transcription factor G(Mafg) | −2.383068 |
| NM_022388 | FXYD domain-containing ion transport regulator 4(Fxyd4) | −2.897865 |
| NM_022394 | scaffold attachment factor B(Safb) | −2.659302 |
| NM_022502 | palmitoyl-protein thioesterase 1(Ppt1) | −1.90058 |
| NM_022503 | cytochrome c oxidase subunit VIIa polypeptide 2(Cox7a2) | −1.523241 |
| NM_022504 | ribosomal protein L36(Rpl36) | −1.906632 |
| NM_022506 | ribosomal protein L31(Rpl31) | −2.087805 |
| NM_022530 | prolactin family 7, subfamily a, member 3(Prl7a3) | −1.576727 |
| NM_022595 | PDGFA associated protein 1(Pdap1) | −2.240738 |
| NM_022597 | cathepsin B(Ctsb) | −2.37573 |
| NM_022612 | BCL2 like 11(Bcl2l11) | −1.597404 |
| NM_022634 | leukocyte specific transcript 1(Lst1) | −1.756651 |
| NM_022694 | staphylococcal nuclease and tudor domain containing 1(Snd1) | −2.119255 |
| NM_022711 | steroid 5 alpha-reductase 2(Srd5a2) | −1.557239 |
| NM_022797 | glutamate ionotropic receptor NMDA type subunit 2D(Grin2d) | −2.63892 |
| NM_022863 | iron responsive element binding protein 2(Ireb2) | −1.923661 |
| NM_022867 | microtubule-associated protein 1 light chain 3 beta(Map1lc3b) | −1.780024 |
| NM_022949 | ribosomal protein L14(Rpl14) | −1.533009 |
| NM_023095 | mannosyl (alpha-1,6-)-glycoprotein beta-1,6-N-acetyl-glucosaminyltransferase(Mgat5) | −2.000325 |
| NM_023100 | neuromedin U receptor 1(Nmur1) | −1.627031 |
| NM_023974 | synaptoporin(Synpr) | −1.803122 |
| NM_023998 | taste receptor, type 2, member 13(Tas2r13) | −1.675877 |
| NM_024156 | annexin A6(Anxa6) | −1.687137 |
| NM_024346 | stathmin 3(Stmn3) | −1.575717 |
| NM_024360 | hes family bHLH transcription factor 1(Hes1) | −2.146919 |
| NM_024385 | hematopoietically expressed homeobox(Hhex) | −1.692521 |
| NM_024388 | nuclear receptor subfamily 4, group A, member 1(Nr4a1) | −2.520533 |
| NM_024397 | abl-interactor 1(Abi1) | −1.624808 |
| NM_024404 | heterogeneous nuclear ribonucleoprotein D(Hnrnpd) | −3.042042 |
| NM_024483 | adrenoceptor alpha 1D(Adra1d) | −1.736564 |
| NM_024484 | 5′-aminolevulinate synthase 1(Alas1) | −1.714727 |
| NM_030857 | LYN proto-oncogene, Src family tyrosine kinase(Lyn) | −2.18526 |
| NM_031008 | adaptor-related protein complex 2, alpha 2 subunit(Ap2a2) | −1.552738 |
| NM_031055 | matrix metallopeptidase 9(Mmp9) | −2.852522 |
| NM_031057 | aldehyde dehydrogenase 6 family, member A1(Aldh6a1) | −1.519431 |
| NM_031088 | prostaglandin E receptor 2(Ptger2) | −2.221709 |
| NM_031127 | sulfite oxidase(Suox) | −2.397507 |
| NM_031236 | fucosyltransferase 1(Fut1) | −1.985466 |
| NM_031327 | cysteine-rich, angiogenic inducer, 61(Cyr61) | −1.638485 |
| NM_031346 | polypyrimidine tract binding protein 3(Ptbp3) | −1.590357 |
| NM_031531 | serine (or cysteine) peptidase inhibitor, clade A, member 3N(Serpina3n) | −2.119708 |
| NM_031549 | transgelin(Tagln) | −2.37869 |
| NM_031563 | Y box binding protein 1(Ybx1) | −1.584841 |
| NM_031590 | WNT1 inducible signaling pathway protein 2(Wisp2) | −1.877269 |
| NM_031678 | period circadian clock 2(Per2) | −1.587107 |
| NM_031706 | ribosomal protein S8(Rps8) | −2.051886 |
| NM_031719 | chloride nucleotide-sensitive channel 1A(Clns1a) | −1.549741 |
| NM_031720 | deiodinase, iodothyronine, type II(Dio2) | −1.958028 |
| NM_031786 | tripartite motif-containing 3(Trim3) | −1.668096 |
| NM_031792 | sperm associated antigen 4(Spag4) | −1.634627 |
| NM_031800 | death effector domain-containing(Dedd) | −1.641957 |
| NM_031808 | calpain 6(Capn6) | −1.820196 |
| NM_031810 | defensin beta 1(Defb1) | −2.054222 |
| NM_031828 | potassium calcium-activated channel subfamily M alpha 1(Kcnma1) | −1.606975 |
| NM_032056 | transporter 2, ATP binding cassette subfamily B member(Tap2) | −2.753854 |
| NM_032062 | kalirin, RhoGEF kinase(Kalrn) | −1.810885 |
| NM_033021 | SEC31 homolog A, COPII coat complex component(Sec31a) | −1.750599 |
| NM_033095 | crystallin, gamma D(Crygd) | −1.569465 |
| NM_053649 | kringle containing transmembrane protein 1(Kremen1) | −1.821446 |
| NM_053674 | phytanoyl-CoA 2-hydroxylase(Phyh) | −1.905711 |
| NM_053701 | calcium voltage-gated channel subunit alpha1 F(Cacna1f) | −1.62128 |
| NM_080689 | dynamin 1(Dnm1) | −1.617324 |
| NM_080783 | UDP-galactose-4-epimerase(Gale) | −2.373817 |
| NM_130405 | KH RNA binding domain containing, signal transduction associated 1(Khdrbs1) | −1.779596 |
| NM_131914 | caveolin 2(Cav2) | −1.636241 |
| NM_133403 | E1A binding protein p300(Ep300) | −2.567324 |
| NM_133594 | small ubiquitin-like modifier 2(Sumo2) | −1.569797 |
| NM_134375 | NLR family, pyrin domain containing 6(Nlrp6) | −2.192127 |
| NM_134387 | dicarbonyl and L-xylulose reductase(Dcxr) | −2.210838 |
| NM_134432 | angiotensinogen(Agt) | −1.737263 |
| NM_134457 | siah E3 ubiquitin protein ligase 2(Siah2) | −1.625673 |
| NM_138851 | prokineticin 1(Prok1) | −1.918202 |
| NM_138890 | EH-domain containing 3(Ehd3) | −1.507669 |
| NM_138976 | mitofusin 1(Mfn1) | −1.671108 |
| NM_139082 | BMP and activin membrane-bound inhibitor(Bambi) | −2.450906 |
| NM_139097 | sodium voltage-gated channel beta subunit 3(Scn3b) | −3.059727 |
| NM_139258 | Bcl2 modifying factor(Bmf) | −2.168055 |
| NM_139336 | UDP-glucuronate decarboxylase 1(Uxs1) | −1.54412 |
| NM_145673 | MAF bZIP transcription factor K(Mafk) | −1.536959 |
| NM_147137 | cystatin SC(LOC257643) | −3.032693 |
| NM_152849 | homeobox and leucine zipper encoding(Homez) | −1.725448 |
| S75437 | No data | −1.965821 |
| U03407 | proline rich, lacrimal 1(Prol1) | −2.285635 |
| U03417 | olfactomedin 1(Olfm1) | −1.797439 |
| U03630 | store-operated calcium entry-associated regulatory factor(Saraf) | −2.016451 |
| U20286 | torsin 1A interacting protein 1(Tor1aip1) | −1.727547 |
| U21954 | Eph receptor A7(Epha7) | −1.630146 |
| U22663 | No data | −1.993711 |
| U24489 | tenascin XA, pseudogene 1(Tnxa-ps1) | −2.172308 |
| U28356 | protein tyrosine phosphatase, non-receptor type 7(Ptpn7) | −2.374227 |
| U35371 | contactin 4(Cntn4) | −1.667741 |
| U40652 | protein tyrosine phosphatase, receptor type, N(Ptprn) | −1.867797 |
| U41663 | neuroligin 3(Nlgn3) | −1.591787 |
| U41803 | mitofusin 2(Mfn2) | −1.574964 |
| U41853 | hypoxia up-regulated 1(Hyou1) | −1.657766 |
| U44129 | lectin, mannose-binding, 1(Lman1) | −2.451062 |
| U49694 | acyl-CoA thioesterase 7(Acot7) | −1.924287 |
| U50948 | No data | −1.881066 |
| U53420 | No data | −4.016245 |
| U56241 | MAF bZIP transcription factor B(Mafb) | −2.421294 |
| U56936 | killer cell lectin-like receptor subfamily B member 1B(Klrb1b) | −1.830188 |
| U57362 | collagen type XII alpha 1 chain(Col12a1) | −2.0687 |
| U62316 | solute carrier family 16 member 7(Slc16a7) | −2.847701 |
| U69702 | activin A receptor type 1C(Acvr1c) | −1.705311 |
| U76206 | purinergic receptor P2Y14(P2ry14) | −2.320753 |
| U78304 | cartilage acidic protein 1(Crtac1) | −1.778874 |
| U81037 | neuronal cell adhesion molecule(Nrcam) | −1.502983 |
| U92010 | similar to RIKEN cDNA D230025D16Rik(RGD621098) | −1.975117 |
| V01222 | albumin(Alb) | −2.265804 |
| X01153 | No data | −1.843423 |
| X05034 | No data | −1.718179 |
| X13309 | WAP four-disulfide core domain 18(Wfdc18) | −2.535182 |
| X53003 | acetyl-CoA carboxylase alpha(Acaca) | −1.756607 |
| X54549 | transcription factor 3(Tcf3) | −2.43995 |
| X56328 | No data | −2.636174 |
| X60822 | methionine adenosyltransferase 1A(Mat1a) | −1.586548 |
| X64411 | ubiquitin protein ligase E3 component n-recognin 5(Ubr5) | −2.174631 |
| X68101 | dedicator of cytokinesis 9(Dock9) | −4.49773 |
| X71463 | No data | −1.940264 |
| X73579 | Fc fragment of IgE receptor II(Fcer2) | −1.640805 |
| X74815 | myosin IE(Myo1e) | −1.973621 |
| X78167 | ribosomal protein L15(Rpl15) | −2.28316 |
| X78604 | ADP-ribosylation factor like GTPase 5A(Arl5a) | −1.523338 |
| X84004 | dual specificity phosphatase 1(Dusp1) | −2.325959 |
| Y12178 | ceruloplasmin(Cp) | −1.709408 |
| Y17319 | No data | −1.766901 |
| Y17325 | NSA2 ribosome biogenesis homolog(Nsa2) | −1.702431 |
| Y17328 | crystallin, mu(Crym) | −1.788589 |
References
- Potempa, J.; Mydel, P.; Koziel, J. The case for periodontitis in the pathogenesis of rheumatoid arthritis. Nat. Rev. Rheumatol. 2017, 13, 606–620. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Schett, G. The pathogenesis of rheumatoid arthritis. N. Engl. J. Med. 2011, 365, 2205–2219. [Google Scholar] [CrossRef] [PubMed]
- Firestein, G.S.; McInnes, I.B. Immunopathogenesis of Rheumatoid Arthritis. Immunity 2017, 46, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Rausch, A.K.; Osthoff, K.; Niedermann, J.; Braun, J.; Adams, N.; Brodin, H.; Dagfinrud, T.; Duruoz, B.A.; Esbensen, K.P.; Günther, E.; et al. 2018 EULAR recommendations for physical activity in people with inflammatory arthritis and osteoarthritis. Ann. Rheum. Dis. 2018, 77, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Smolen, J.S.; Landewé, R.; Bijlsma, J.; Burmester, G.; Chatzidionysiou, K.; Dougados, M.; Nam, J.; Ramiro, S.; Voshaar, M.; van Vollenhoven, R.; et al. EULAR recommendations for the management of rheumatoid arthritis with synthetic and biological disease-modifying antirheumatic drugs: 2016 update. Ann. Rheum. Dis. 2017, 76, 960–977. [Google Scholar] [CrossRef] [PubMed]
- Anvar, N.; Matlabi, H.; Safaiyan, A.; Allahverdipour, H.; Kolahi, S. Effectiveness of self-management program on arthritis symptoms among older women: A randomized controlled trial study. Health Care Women Int. 2018, 39, 1326–1339. [Google Scholar] [CrossRef] [PubMed]
- Agca, R.; Heslinga, S.C.; Rollefstad, S.; Heslinga, M.; McInnes, I.B.; Peters, M.J.L.; Kvien, T.K.; Dougados, M.; Radner, H.; Atzeni, F.; et al. EULAR recommendations for cardiovascular disease risk management in patients with rheumatoid arthritis and other forms of inflammatory joint disorders: 2015/2016 update. Ann. Rheum. Dis. 2017, 76, 17–28. [Google Scholar] [CrossRef]
- Law, R.J.; Saynor, Z.L.; Gabbitas, J.; Jones, J.; Kraus, A.; Breslin, A.; Maddison, P.J.; Thom, J.M. The Effects of Aerobic and Resistance Exercise on Markers of Large Joint Health in Stable Rheumatoid Arthritis Patients: A Pilot Study. Musculoskeletal Care 2015, 13, 222–235. [Google Scholar] [CrossRef]
- Bergstra, S.A.; Murgia, A.; Te Velde, A.F.; Caljouw, S.R. A systematic review into the effectiveness of hand exercise therapy in the treatment of rheumatoid arthritis. Clin. Rheumatol. 2014, 33, 1539–1548. [Google Scholar] [CrossRef]
- van Rensburg, D.C.J.; Ker, J.A.; Grant, C.C.; Fletcher, L. Effect of exercise on cardiac autonomic function in females with rheumatoid arthritis. Clin. Rheumatol. 2012, 31, 1155–1162. [Google Scholar] [CrossRef]
- Stavropoulos-Kalinoglou, A.; Metsios, G.S.; van Zanten, J.J.J.C.S.V.; Nightingale, P.; Kitas, G.D.; Koutedakis, Y. Individualised aerobic and resistance exercise training improves cardiorespiratory fitness and reduces cardiovascular risk in patients with rheumatoid arthritis. Ann. Rheum. Dis. 2013, 72, 1819–1825. [Google Scholar] [CrossRef] [PubMed]
- Waite-Jones, J.M.; Hale, C.A.; Lee, H.Y. Psychosocial effects of Tai Chi exercise on people with rheumatoid arthritis. J. Clin. Nurs. 2013, 22, 3053–3061. [Google Scholar] [CrossRef]
- Perandini, L.A.; de Sá-Pinto, A.L.; Roschel, H.; Benatti, F.B.; Lima, F.R.; Bonfá, E.; Gualano, B. Exercise as a therapeutic tool to counteract inflammation and clinical symptoms in autoimmune rheumatic diseases. Autoimmun. Rev. 2012, 12, 218–224. [Google Scholar] [CrossRef]
- Sharif, S.; Thomas, J.M.; Donley, D.A.; Gilleland, D.L.; Bonner, D.E.; McCrory, J.L.; Hornsby, W.G.; Zhao, H.; Lively, M.W.; Hornsby, J.A.A.; et al. Resistance exercise reduces skeletal muscle cachexia and improves muscle function in rheumatoid arthritis. Case Rep. Med. 2011, 2011, 205691. [Google Scholar] [CrossRef] [PubMed]
- Feldthusen, C.; Dean, E.; Forsblad-d’Elia, H.; Mannerkorpi, K. Effects of Person-Centered Physical Therapy on Fatigue-Related Variables in Persons With Rheumatoid Arthritis: A Randomized Controlled Trial. Arch. Phys. Med. Rehabil. 2016, 97, 26–36. [Google Scholar] [CrossRef]
- Cambré, I.; Gaublomme, D.; Schryvers, N.; Lambrecht, S.; Lories, R.; Venken, K.; Elewaut, D. Running promotes chronicity of arthritis by local modulation of complement activators and impairing T regulatory feedback loops. Ann. Rheum. Dis. 2019, 6, 787–795. [Google Scholar]
- Cambré, I.; Gaublomme, D.; Burssens, A.; Jacques, P.; Schryvers, N.; De Muynck, A.; Meuris, L.; Lambrecht, S.; Carter, S.; de Bleser, P.; et al. Mechanical strain determines the site-specific localization of inflammation and tissue damage in arthritis. Nat. Commun. 2018, 9, 4613. [Google Scholar] [CrossRef]
- Bevaart, L.; Vervoordeldonk, M.J.; Tak, P.P. Evaluation of therapeutic targets in animal models of arthritis: How does it relate to rheumatoid arthritis? Arthritis Rheum. 2010, 62, 2192–2205. [Google Scholar] [CrossRef]
- Bordy, R.; Verhoeven, F.; Tournier-Nappey, M.; Wendling, D.; Demougeot, C.; Totoson, P. Methotrexate did not improve endothelial function in rheumatoid arthritis: A study in rats with adjuvant-induced arthritis. Clin. Exp. Rheumatol. 2018, 1, 81–88. [Google Scholar]
- Abdel-Maged, A.E.S.; Gad, A.M.; Abdel-Aziz, A.K.; Aboulwafa, M.M.; Azab, S.S. Comparative study of anti-VEGF Ranibizumab and Interleukin-6 receptor antagonist Tocilizumab in Adjuvant-induced Arthritis. Toxicol. Appl. Pharmacol. 2018, 356, 65–75. [Google Scholar] [CrossRef]
- Vidal, B.; Cascão, R.; Finnilä, M.A.J.; Lopes, I.P.; da Glória, V.G.; Saarakkala, S.; Zioupos, P.; Canhão, H.; Fonseca, J.E. Effects of tofacitinib in early arthritis-induced bone loss in an adjuvant-induced arthritis rat model. Rheumatology 2018, 57, 1461–1471. [Google Scholar] [CrossRef] [PubMed]
- Kaliyappan, K.; Palanisamy, M.; Govindarajan, R.; Duraiyan, J. Microarray and its applications. J. Pharm. Bioallied Sci. 2012, 4, 310. [Google Scholar] [CrossRef] [PubMed]
- Snekhalatha, U.; Anburajan, M.; Venkatraman, B.; Menaka, M. Evaluation of complete Freund’s adjuvant-induced arthritis in a Wistar rat model. Comparison of thermography and histopathology. Z. Für Rheumatol. 2013, 72, 375–382. [Google Scholar] [CrossRef]
- González-Chávez, S.A.; Pacheco-Tena, C.; Macías-Vázquez, C.E.; Luévano-Flores, E. Assessment of different decalcifying protocols on Osteopontin and Osteocalcin immunostaining in whole bone specimens of arthritis rat model by confocal immunofluorescence. Int. J. Clin. Exp. Pathol. 2013, 6, 1972–1983. [Google Scholar] [PubMed]
- Huang, D.W.; Sherman, B.T.; Tan, Q.; Kir, J.; Liu, D.; Bryant, D.; Guo, Y.; Stephens, R.; Baseler, M.W.; Lane, H.C.; et al. DAVID Bioinformatics Resources: Expanded annotation database and novel algorithms to better extract biology from large gene lists. Nucleic Acids Res. 2007, 35, W169–W175. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Isserlin, R.; Merico, D.; Voisin, V.; Bader, G.D. Enrichment Map—A Cytoscape app to visualize and explore OMICs pathway enrichment results. F1000 Res. 2014, 3, 141. [Google Scholar] [CrossRef]
- Bader, G.D.; Hogue, C.W.V. An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinform. 2003, 4, 2. [Google Scholar] [CrossRef]
- Geenen, R.; Overman, C.L.; Christensen, R.; Åsenlöf, P.; Capela, S.; Huisinga, K.L.; Husebø, M.E.P.; Köke, A.J.A.; Paskins, Z.; Pitsillidou, I.A.; et al. EULAR recommendations for the health professional’s approach to pain management in inflammatory arthritis and osteoarthritis. Ann. Rheum. Dis. 2018, 77, 797–807. [Google Scholar] [CrossRef]
- Peters, C.L.; Morris, C.J.; Mapp, P.I.; Blake, D.R.; Lewis, C.E.; Winrow, V.R. The transcription factors hypoxia-inducible factor 1alpha and Ets-1 colocalize in the hypoxic synovium of inflamed joints in adjuvant-induced arthritis. Arthritis Rheum. 2004, 50, 291–296. [Google Scholar] [CrossRef]
- Jeon, C.H.; Ahn, J.K.; Chai, J.Y.; Kim, H.J.; Bae, E.K.; Park, S.H.; Cho, E.Y.; Cha, H.S.; Ahn, K.S.; Koh, E.M. Hypoxia appears at pre-arthritic stage and shows co-localization with early synovial inflammation in collagen induced arthritis. Clin. Exp. Rheumatol. 2008, 26, 646–648. [Google Scholar] [PubMed]
- Quiñonez-Flores, C.M.; González-Chávez, S.A.; Pacheco-Tena, C. Hypoxia and its implications in rheumatoid arthritis. J. Biomed. Sci. 2016, 23, 1. [Google Scholar] [CrossRef] [PubMed]
- Quiñonez-Flores, C.M.; González-Chávez, S.A.; Del Río Nájera, D.; Pacheco-Tena, C. Oxidative Stress Relevance in the Pathogenesis of the Rheumatoid Arthritis: A Systematic Review. BioMed Res. Int. 2016, 2016, 6097417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fearon, U.; Canavan, M.; Biniecka, M.; Veale Hypoxia, D.J. mitochondrial dysfunction and synovial invasiveness in rheumatoid arthritis. Nat. Rev. Rheumatol. 2016, 12, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.E.; Harlow, L.A.; Haines, G.K.; Amento, E.P.; Unemori, E.N.; Wong, W.L.; Pope, R.M.; Ferrara, N. Vascular endothelial growth factor. A cytokine modulating endothelial function in rheumatoid arthritis. J. Immunol. Baltim. Md 1950 1994, 152, 4149–4156. [Google Scholar]
- Asahara, T.; Chen, D.; Takahashi, T.; Fujikawa, K.; Kearney, M.; Magner, M.; Yancopoulos, G.D.; Isner, J.M. Tie2 receptor ligands, angiopoietin-1 and angiopoietin-2, modulate VEGF-induced postnatal neovascularization. Circ. Res. 1998, 83, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Holash, J.; Maisonpierre, P.C.; Compton, D.; Boland, P.; Alexander, C.R.; Zagzag, D.; Yancopoulos, G.D.; Wiegand, S.J. Vessel cooption, regression, and growth in tumors mediated by angiopoietins and VEGF. Science 1999, 284, 1994–1998. [Google Scholar] [CrossRef] [Green Version]
- Paleolog, E.M.; Young, S.; Stark, A.C.; McCloskey, R.V.; Feldmann, M.; Maini, R.N. Modulation of angiogenic vascular endothelial growth factor by tumor necrosis factor alpha and interleukin-1 in rheumatoid arthritis. Arthritis Rheum. 1998, 41, 1258–1265. [Google Scholar] [CrossRef]
- Etherington, P.J.; Winlove, P.; Taylor, P.; Paleolog, E.; Miotla, J.M. VEGF release is associated with reduced oxygen tensions in experimental inflammatory arthritis. Clin. Exp. Rheumatol. 2002, 20, 799–805. [Google Scholar]
- Chang, X.; Wei, C. Glycolysis and rheumatoid arthritis. Int. J. Rheum. Dis. 2011, 14, 217–222. [Google Scholar] [CrossRef]
- Song, G.; Lu, Q.; Fan, H.; Zhang, X.; Ge, L.; Tian, R.; Wang, S.; Feng, T.; Pan, J.; Feng, J.; et al. Inhibition of hexokinases holds potential as treatment strategy for rheumatoid arthritis. Arthritis Res. Ther. 2019, 21, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hitchon, C.A.; El-Gabalawy, H.S. Oxidation in rheumatoid arthritis. Arthritis Res. Ther. 2004, 6, 265–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mountz, J.D.; Zhang, H.G.; Wang, Y.; Xie, J.F.; Liang, X.; Hsu, H.C.; Curiel, D.T. AKT regulates TNF-alpha-mediated apoptosis of rheumatoid arthritis synovial fibrblasts. Arthritis Res. 2001, 3, 8. [Google Scholar] [CrossRef]
- Clark, A.R.; Dean, J.L. The p38 MAPK Pathway in Rheumatoid Arthritis: A Sideways Look. Open Rheumatol. J. 2001, 6, 209–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, G.; Cooles, F.A.; Isaacs, J.D.; Hilkens, C.M. Emerging immunotherapies for rheumatoid arthritis. Hum. Vaccines Immunother. 2014, 10, 822–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGonagle, D.; Gibbon, W.; O’Connor, P.; Green, M.; Pease, C.; Emery, P. Characteristic magnetic resonance imaging entheseal changes of knee synovitis in spondylarthropathy. Arthritis Rheum. 1998, 41, 694–700. [Google Scholar] [CrossRef]
- Blazek, A.D.; Nam, J.; Gupta, R.; Pradhan, M.; Perera, P.; Weisleder, N.L.; Hewett, T.E.; Chaudhari, A.M.; Lee, B.S.; Leblebicioglu, B.; et al. Exercise-driven metabolic pathways in healthy cartilage. Osteoarthr. Cartil. 2016, 24, 1210–1222. [Google Scholar] [CrossRef] [Green Version]
- Shay, A.K.; Bliven, M.L.; Scampoli, D.N.; Otterness, I.G.; Milici, A.J. Effects of exercise on synovium and cartilage from normal and inflamed knees. Rheumatol. Int. 1995, 14, 183–189. [Google Scholar] [CrossRef]
- Gomes, R.P.; Bressan, E.; da Silva, T.M.; da Silva Gevaerd, M.; Tonussi, C.R.; Domenech, S.C. Effects of one minute and ten minutes of walking activity in rats with arthritis induced by complete Freund’s adjuvant on pain and edema symptoms. Rev. Bras. Reumatol. 2014, 54, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Gomes, R.P.; Bressan, E.; da Silva, T.M.; Domenech, S.C.; Tonussi, C.R. Evidências de que um protocolo de atividade física pode reduzir a contagem de leucócitos sinoviais de ratos artríticos. Rev. Bras. Med. Esporte 2013, 19, 70–73. [Google Scholar] [CrossRef] [Green Version]
- Shimomura, S.; Inoue, H.; Arai, Y.; Nakagawa, S.; Fujii, Y.; Kishida, T.; Ichimaru, S.; Tsuchida, S.; Shirai, T.; Ikoma, K.; et al. Treadmill Running Ameliorates Destruction of Articular Cartilage and Subchondral Bone, Not Only Synovitis, in a Rheumatoid Arthritis Rat Model. Int. J. Mol. Sci. 2018, 19, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, D.Y.; Lee, S.R.; Kim, N.; Ko, K.S.; Rhee, B.D.; Han, J. Humanized animal exercise model for clinical implication. Pflugers Arch. 2014, 466, 1673–1687. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Effect on Regulation | Number of Genes | Genes | p-Value | |
|---|---|---|---|---|---|
| Biological Process | Response to hypoxia | Up | 12 | Camk2d, Cat, Cdkn1b, Il18, Lepr, Mt3, Nppa, Ptk2b, Rhoa, Tgfbr3, Vegfa, Vhl | 3.1 × 10−3 |
| Down | 17 | Alas1, Bnip3, Cited2, Ep300, Smad3, Casp3, Hmbs, Hyou1, Igf1, Itga2, Icam1, Il1a, Mmp9, Kcnma1, Th, Ucp2, Ucp3 | 7.5 × 10−6 | ||
| Oxidation-reduction process | Up | 19 | Akr1a1, Aifm1, Cbr1, Cyp2d2, Cyp2t1, Cyp27b1, Cyp4f6, Cyp51, Degs1, Gpx1, Hao2, Ido1, Kif1b, Me1, Oxr1, Prdx2, Pah, Txn1, Tpo | 1.2 × 10−2 | |
| Down | 26 | Haao, Hibadh, Ndufa12, Aldh6a1, Aox1, Cp, Crym, Cyp19a1, Cyp1b1, Cyp2a3, Cyp2b12, Cyp2p2c7, Cyp2d3, Dio2, Dcxr, G6pd, Glrx, Gpx6, Hsd3b6, Hadhb, Ldhb, Scd, Srd5a2, Suox, Tpo, Th | 4.9 × 10−5 | ||
| Hydrogen peroxide catabolic process | Up | 4 | Cat, Gpx1, Prdx2, Tpo | 2.5 × 10−3 | |
| Response to oxidative stress | Up | 8 | Akt1, Cat, Gpx1, Mt3, Map2k1, Oxr1, Prdx2, Tpo | 7.4 × 10−3 | |
| Cellular response to oxidative stress | Up | 5 | Ggt1, Mt3, Nfe2l2, Prdx2, Txn1 | 3.2 × 10−2 | |
| Cellular response to hydrogen peroxide | Up | 5 | Anxa1, Aifm1, Il18, Nfe2l2, Ppp5c | 3.5 × 10−2 | |
| Response to reactive oxygen species | Up | 3 | Cat, Gpx1, Ptk2b | 4.1 × 10−2 | |
| Inflammatory response | Up | 12 | Akt1, Ccl4, Anxa1, Cxcl3, Csf1, Cyp4f6, Crlf2, Hmgb1, Ido1, Il18, Mep1b, Nfe2l2 | 6.3 × 10−3 | |
| KEGG signaling pathway | HIF-1 signaling pathway | Up | 14 | Akt1, Akt3, Camk2a, Camk2d, Cdkn1b, Hk1, Ifngr1, Map2k1, Nppa, Pik3cb, Pik3r1, Tceb2, Vegfa, Vhl | 2.5 × 10−7 |
| VEGF signaling pathway | Up | 8 | Akt1, Akt3, Map2k1, Pik3cb, Pik3r1, Ppp3cb, Ppp3r2, Vegfa | 2.5 × 10−4 | |
| Rheumatoid Arthritis | Up | 6 | Atp6v0a1, Atp6v0e1, Cd80, Csf1, Il18, Vegfa | 4.2 × 10−2 | |
| T cell receptor signaling pathway | Up | 10 | Akt1, Akt3, Dlg1, Lcp2, Map2k1, Pik3cb, Pik3r1, Ppp3cb, Ppp3r2, Rhoa | 4.5 × 10−4 | |
| B cell receptor signaling pathway | Up | 7 | Akt1, Akt3, Map2k1, Pik3cb, Pik3r1, Ppp3cb, Ppp3r2 | 3.3 × 10−3 | |
| Chemokine signaling pathway | Up | 12 | Akt1, Akt3, Ccl4, Gng8, Adcy5, Arrb1, Cxcl3, Map2k1, Pik3cb, Pik3r1, Ptk2b, Rhoa | 1.3 × 10−3 | |
| PI3K-Akt signaling pathway | Up | 17 | Akt1, Akt3, Faslg, Gng8, Csf1, Cdkn1b, Fgd17, Fgf19, Il3ra, Lpar3, Map2k1, Pik3cb, Pik3r1, Ppp2r2c, Vegfa, Ywhae, Ywhag | 2.2 × 10−3 | |
| Jak-STAT signaling pathway | Up | 10 | Akt1, Akt3, Cish, Crlf2, Ifngr1, Il3ra, Lepr, Pik3cb, Pik3r1, Thpo | 2.2 × 10−3 | |
| TNF signaling pathway | Up | 7 | Akt1, Akt3, Cxcl3, Csf1, Map2k1, Pik3cb, Pik3rc1 | 2.8 × 10−2 | |
| Toll-like receptor signaling pathway | Up | 7 | Akt1, Akt3, Ccl4, Cd80, Map2k1, Pik3cb, Pik3r1 | 1.7 × 10−2 | |
| Wnt signaling pathway | Up | 10 | Wif1, Camk2a, Camk2d, Csnk2a1, Csnk2b, Ctnnb1, Fzd4, Ppp3cb, Ppp3r2, Rhoa | 3.0 × 10−3 | |
| Osteoclast differentiation | Up | 13 | Akt1, Akt3, Fcgr2a, Csfr1, Ifngr1,Lilrb3l, Lcp2, Mapk2k1, Pik3cb, Pik3r1, Ppp3cb, Ppp3r1, Ppp3r2 | 8.6 × 10−5 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Chávez, S.A.; Quiñonez-Flores, C.M.; Espino-Solís, G.P.; Vázquez-Contreras, J.Á.; Pacheco-Tena, C. Exercise Exacerbates the Transcriptional Profile of Hypoxia, Oxidative Stress and Inflammation in Rats with Adjuvant-Induced Arthritis. Cells 2019, 8, 1493. https://doi.org/10.3390/cells8121493
González-Chávez SA, Quiñonez-Flores CM, Espino-Solís GP, Vázquez-Contreras JÁ, Pacheco-Tena C. Exercise Exacerbates the Transcriptional Profile of Hypoxia, Oxidative Stress and Inflammation in Rats with Adjuvant-Induced Arthritis. Cells. 2019; 8(12):1493. https://doi.org/10.3390/cells8121493
Chicago/Turabian StyleGonzález-Chávez, Susana Aideé, Celia María Quiñonez-Flores, Gerardo Pavel Espino-Solís, José Ángel Vázquez-Contreras, and César Pacheco-Tena. 2019. "Exercise Exacerbates the Transcriptional Profile of Hypoxia, Oxidative Stress and Inflammation in Rats with Adjuvant-Induced Arthritis" Cells 8, no. 12: 1493. https://doi.org/10.3390/cells8121493
APA StyleGonzález-Chávez, S. A., Quiñonez-Flores, C. M., Espino-Solís, G. P., Vázquez-Contreras, J. Á., & Pacheco-Tena, C. (2019). Exercise Exacerbates the Transcriptional Profile of Hypoxia, Oxidative Stress and Inflammation in Rats with Adjuvant-Induced Arthritis. Cells, 8(12), 1493. https://doi.org/10.3390/cells8121493