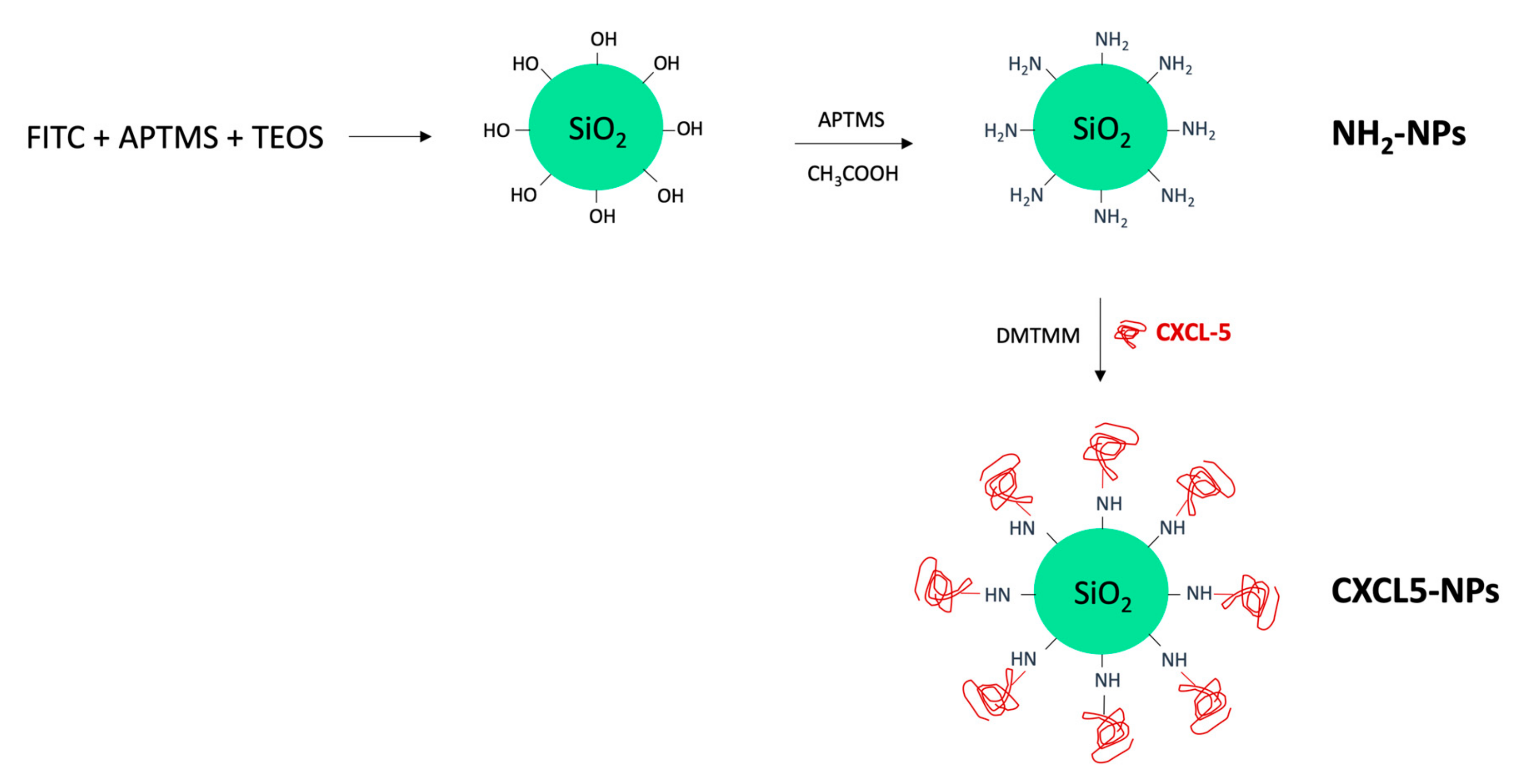
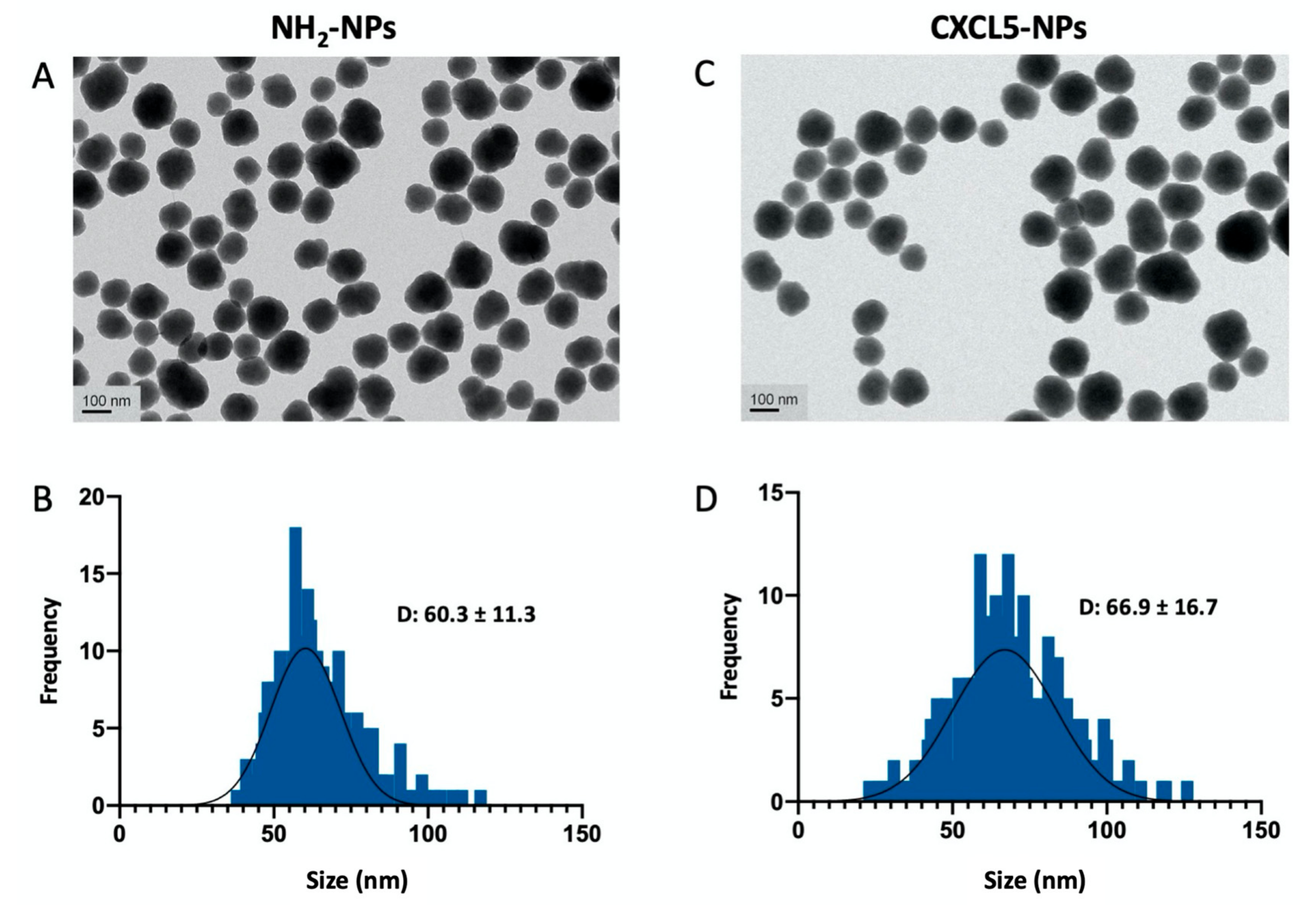
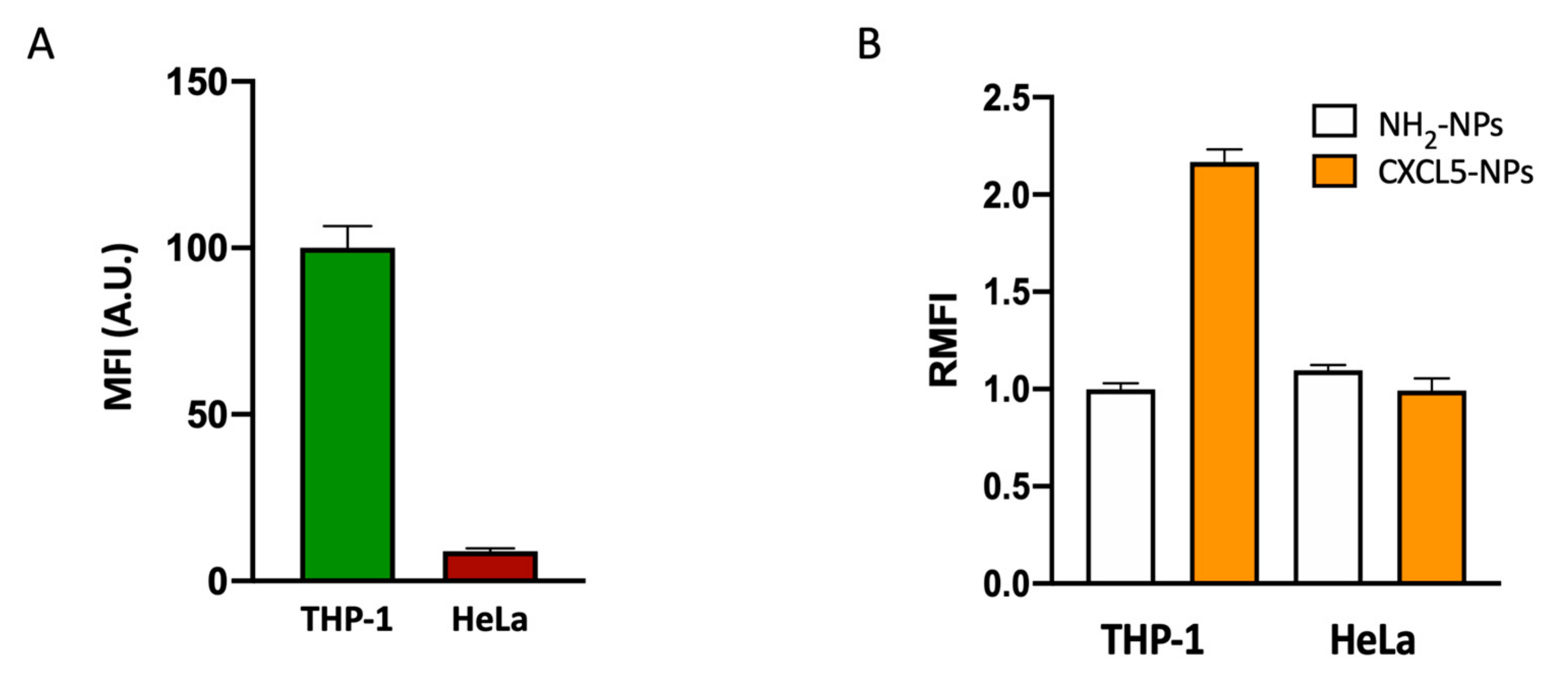
CXCL5 Modified Nanoparticle Surface Improves CXCR2+ Cell Selective Internalization

 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
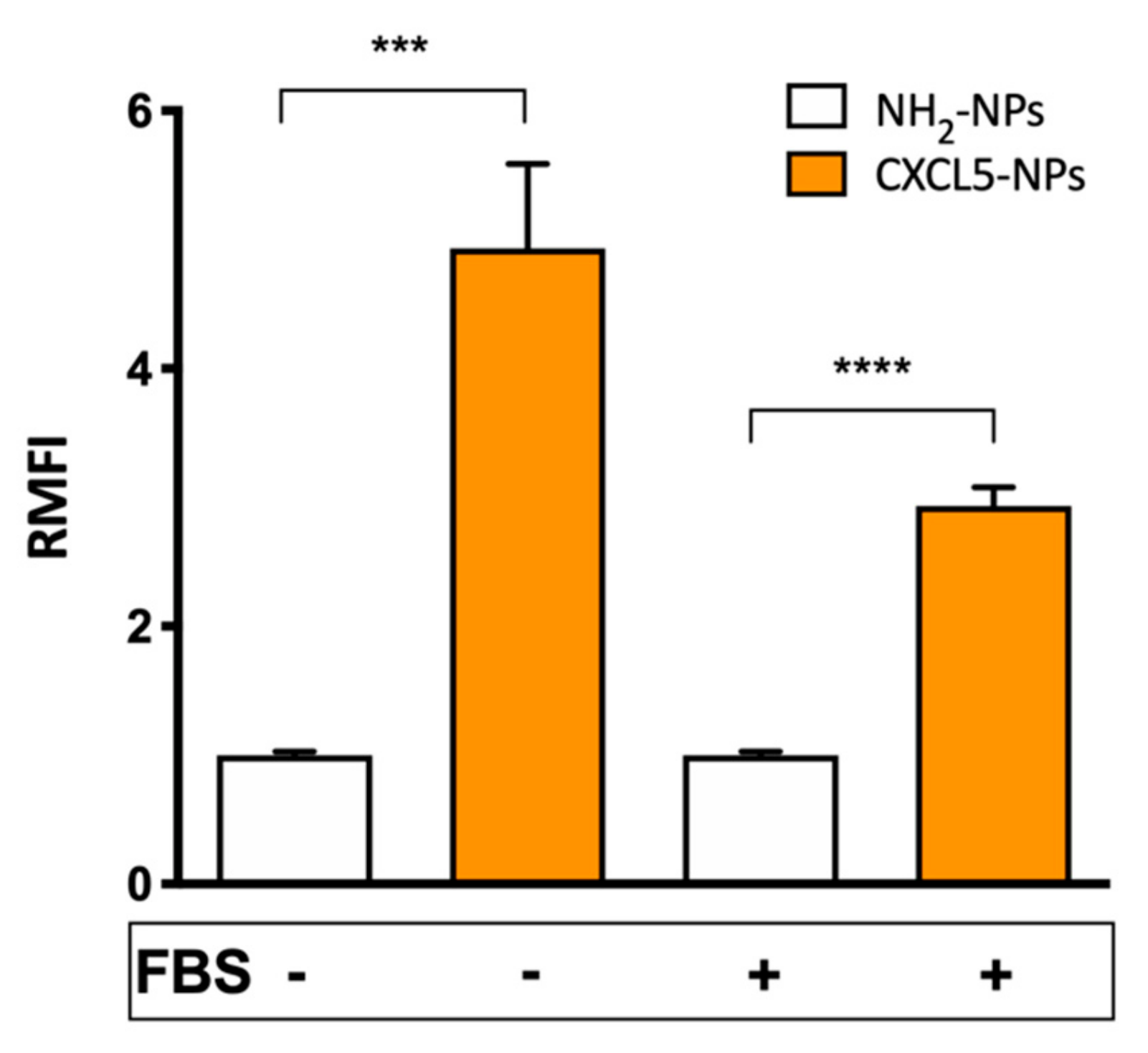{kind=link}
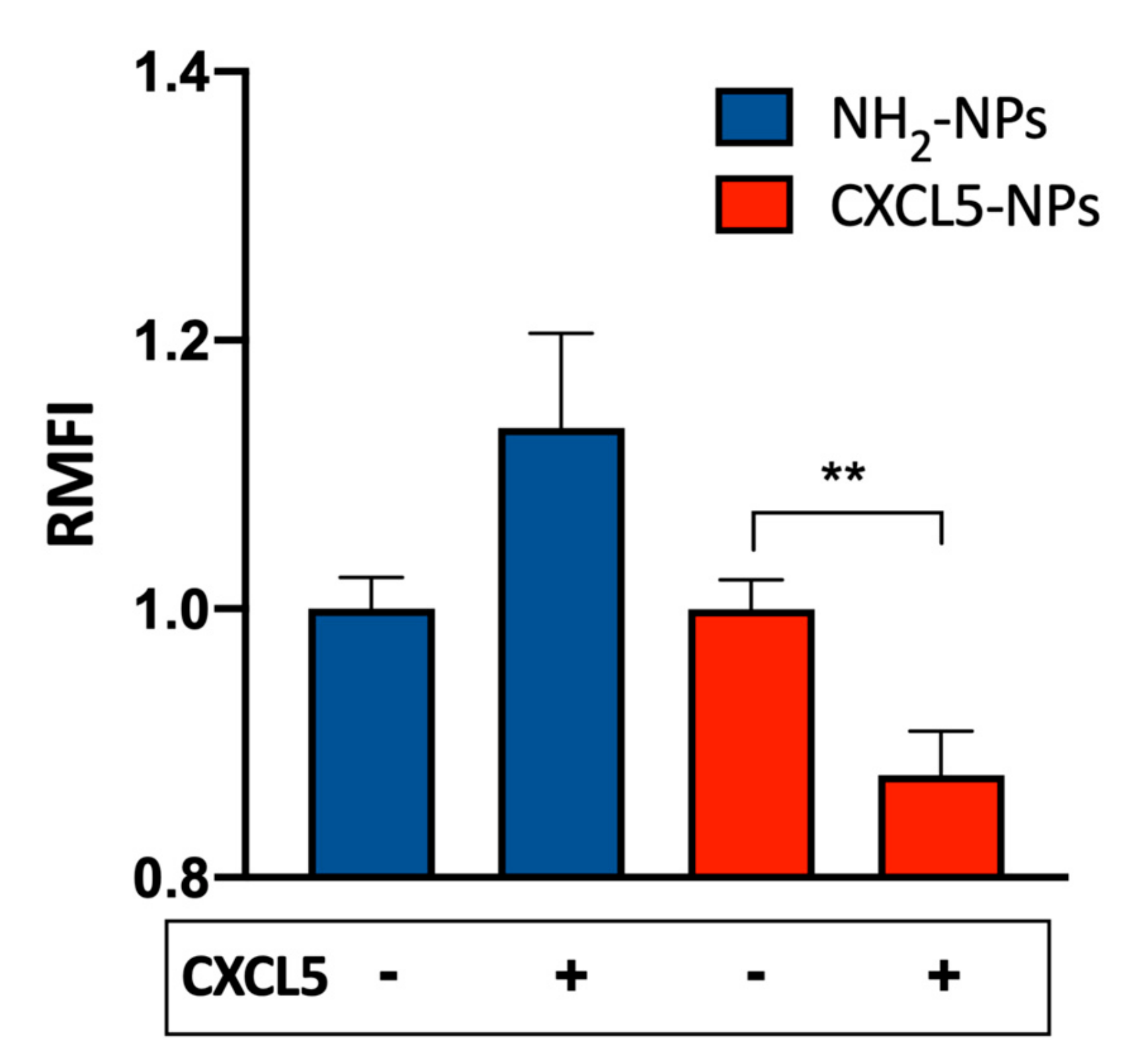{kind=link}
{kind=link}
Abstract
Share and Cite
Cagliani, R.; Gatto, F.; Cibecchini, G.; Marotta, R.; Catalano, F.; Sanchez-Moreno, P.; Pompa, P.P.; Bardi, G. CXCL5 Modified Nanoparticle Surface Improves CXCR2+ Cell Selective Internalization. Cells 2020, 9, 56. https://doi.org/10.3390/cells9010056
Cagliani R, Gatto F, Cibecchini G, Marotta R, Catalano F, Sanchez-Moreno P, Pompa PP, Bardi G. CXCL5 Modified Nanoparticle Surface Improves CXCR2+ Cell Selective Internalization. Cells. 2020; 9(1):56. https://doi.org/10.3390/cells9010056
Chicago/Turabian StyleCagliani, Roberta, Francesca Gatto, Giulia Cibecchini, Roberto Marotta, Federico Catalano, Paola Sanchez-Moreno, Pier Paolo Pompa, and Giuseppe Bardi. 2020. "CXCL5 Modified Nanoparticle Surface Improves CXCR2+ Cell Selective Internalization" Cells 9, no. 1: 56. https://doi.org/10.3390/cells9010056
APA StyleCagliani, R., Gatto, F., Cibecchini, G., Marotta, R., Catalano, F., Sanchez-Moreno, P., Pompa, P. P., & Bardi, G. (2020). CXCL5 Modified Nanoparticle Surface Improves CXCR2+ Cell Selective Internalization. Cells, 9(1), 56. https://doi.org/10.3390/cells9010056