1. Introduction
Aquaculture has become an agronomic activity with noticeable development around the world to respond to the increasing demand of aquatic products for human consumption [
1]. The majority of fish growth is invested in accreting muscle tissue, which is the main edible portion [
2,
3]. Muscle growth is a complex, dynamic process involving both the recruitment of new muscle fibers (hyperplasia) and the growth of existing fibers (hypertrophy) [
4], which is under the control of diverse regulatory factors such as insulin-like growth factors (IGFs), myogenic regulatory factors (MRFs), and myostatin (MSTN) [
5]. The IGF-I is a key regulatory hormone that controls growth in vertebrates [
6], which stimulates both proliferation and differentiation of myoblasts, as well as promoting myotube hypertrophy in vitro and in vivo [
7,
8,
9]. The MRFs are muscle-specific basic helix–loop–helix transcription factors. They include myoblast determination protein (MyoD), myogenic factor 5 (Myf5), myogenin (MyoG), and myogenic regulatory factor 4 (MRF4) [
10]. Myf5 and MyoD are mainly involved in muscle specification and trigger conversion of no muscle cells, such as fibroblasts, into muscle, whereas myogenin and MRF4 act later during myogenesis and allow myotube formation and maturation [
11,
12]. MSTN is a negative regulator of myogenesis, which inhibits myoblast cell proliferation and differentiation [
13]. Despite increased understanding of the regulation of skeletal muscle growth by some of these factors, its regulation by nutrients remains poorly documented in fish.
l-leucine (Leu) as a branched-chain amino acid is nutritionally essential for fish [
14]. Leu deficiency results in depressed growth rate, low feed efficiency, and poor protein retention, as reported for large yellow croaker
Pseudosciaena crocea [
15], rainbow trout
Oncorhynchus mykiss [
16], juvenile golden pompano
Trachinotus ovatus [
17], grass carp
Ctenopharyngodon idellus [
18], black carp
Mylopharyngodon piceus [
14], fingerling channel catfish
Ictalurus punctatus [
19], juvenile hybrid grouper
Epinephelus fuscoguttatus♀ ×
Epinephelus lanceolatus♂ [
20], juvenile blunt snout bream
Megalobrama amblycephala [
21],
Catla catla [
22], and stinging catfish
Heteropneustes fossilis [
23]. Nevertheless, these studies mainly focused on the effects of Leu on growth, protein retention, and immunity. Recently, studies in primary preterm rat satellite cells and porcine myoblasts showed Leu promoted proliferation and differentiation [
24,
25]. Averous et al. (2012) also reported Leu deficiency inhibited the differentiation of both C2C12 myoblasts and primary mice satellite cells through regulating Myf5 and MyoD expression [
26]. These data suggested that Leu might regulate muscle growth through affecting the process of cell proliferation and differentiation. However, the actual role of Leu in regulating muscle growth in fish still needs to be clarified.
The process involved in the increase in muscle growth is associated with accumulation of protein [
27,
28]. The protein deposition of muscle is the result of the balance of protein synthesis and degradation [
29,
30]. Previous studies have shown that nutrition can activate the IGF-I/PI3K/AKT signaling pathway and induce protein synthesis and accretion in rat and rainbow trout [
31,
32,
33]. The target of rapamycin (TOR) is a downstream component of the PI3K/AKT pathway, which plays a crucial role in protein synthesis of fish [
34]. The TOR regulates phosphorylation of its downstream effector ribosomal S6 kinase 1 (S6K1) and the eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1), ultimately promoting protein synthesis of fish [
35,
36,
37,
38]. Muscle protein degradation is primarily through the activation of the ubiquitin proteasome pathway (UPP), which can degrade most cell proteins and contribute to 75% protein degradation in muscle [
39,
40]. The AKT-dependent regulation of the forkhead box O3a (FOXO3a) protein has been shown to play a critical role in UPP pathway [
41,
42]. The AKT is known to phosphorylate FOXO3a, leading to the exclusion of phosphorylated FOXO3a proteins from the nucleus and the suppression of their transcriptional functions, which decreases muscle protein degradation in mammals [
29,
43,
44]. Muscle atrophy F-box (MAFBX) and muscle Ring finger 1 (MURF-1) are responsible for increased protein degradation through the UPP pathway, which can actually be considered the master genes for muscle atrophy and wasting [
39,
45,
46]. Leu could increase muscle protein deposition by regulating protein synthesis and protein degradation in mammals [
29,
47]. Dietary optimal Leu up-regulated liver TOR mRNA expression in juvenile hybrid grouper and juvenile blunt snout bream [
20,
21]. Supplementing media with Leu reduced protein degradation by regulating MAFBX32 expression in rainbow trout primary myocytes [
48]. Those data suggested that Leu might elevate protein deposition by regulating gene expressions related to protein synthesis and protein degradation in fish. However, the evaluation of the effects of Leu on PI3K/AKT/TOR and AKT/FOXO3a pathways in vivo and their contribution to somatic growth have not been previously studied.
Pelteobagrus vachelli♀ ×
Leiocassis longirostris♂ is a hybrid catfish that has been widely cultured in China in recent years. Our previous studies have determined optimal dietary tryptophan levels [
49]. Therefore, the objective of the present study was to investigate the effects of dietary Leu on growth performance and muscle growth, protein synthesis, and degradation-related signaling pathways of hybrid catfish. Furthermore, the dietary Leu requirement for hybrid catfish was evaluated.
4. Discussion
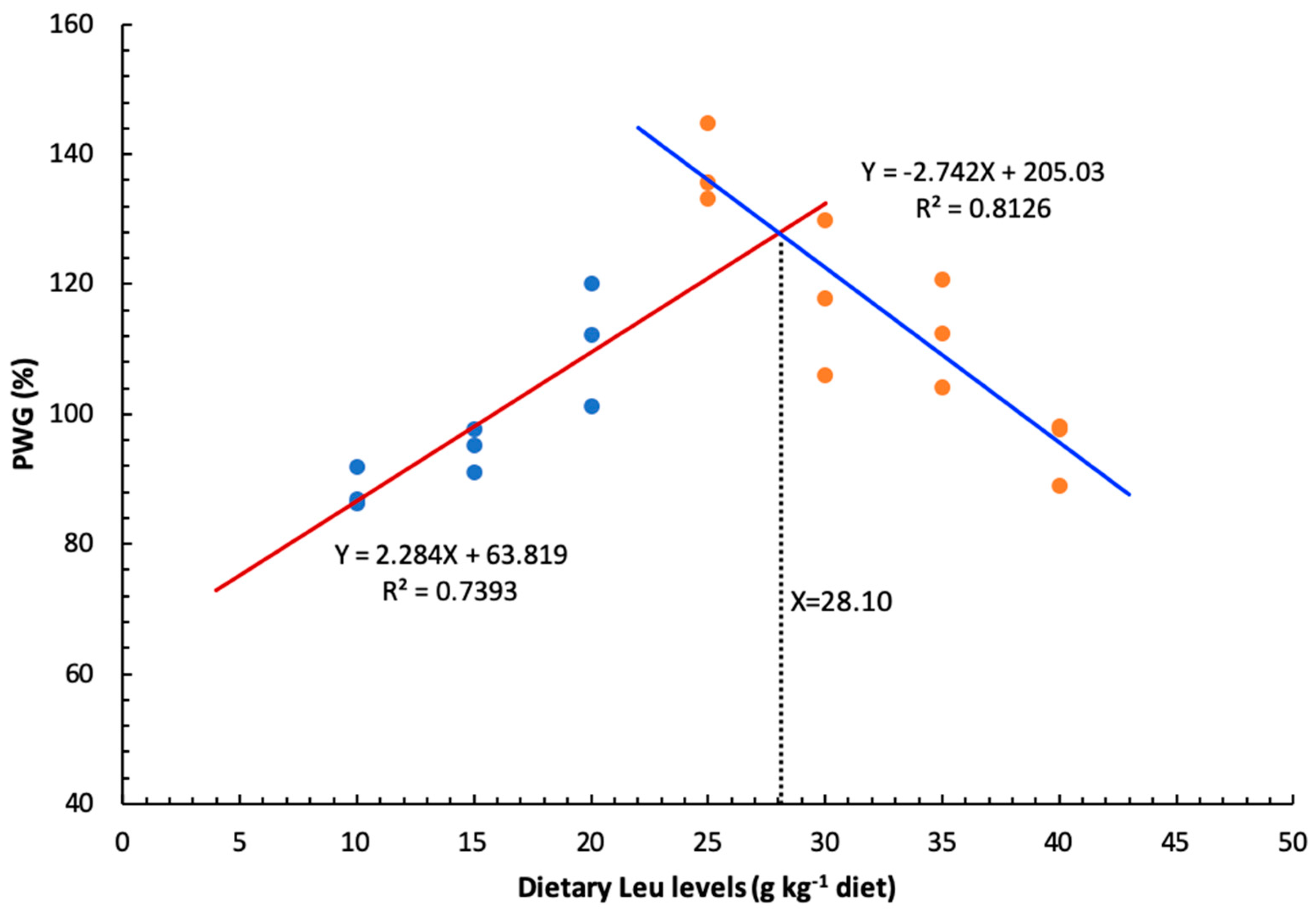
As an essential amino acid, dietary Leu level had a clear effect on growth and feed utilization of hybrid catfish. The optimal dietary Leu requirement for maximal PWG of hybrid catfish was estimated to be 28.10 g kg
−1 of the diet, corresponding to 73.04 g kg
−1 of dietary protein. This value (g kg
−1 of dietary protein) was similar to that of large yellow croaker (67.90 g kg
−1 of dietary protein) [
15], lower than those for rainbow trout (92.00 g kg
−1 of dietary protein) [
16] and juvenile golden pompano (80.20 g kg
−1 of dietary protein) [
17], but higher than those reported for grass carp (42.40 g kg
−1 of dietary protein) [
18], black carp (59.50 g kg
−1 of dietary protein) [
14], fingerling channel catfish (35.00 g kg
−1 of dietary protein) [
19], juvenile hybrid grouper (47.50 g kg
−1 of dietary protein) [
20], juvenile blunt snout bream (47.40 g kg
−1 of dietary protein) [
21],
Catla catla (47.90 g kg
−1 of dietary protein) [
22], and stinging catfish (43.40 g kg
−1 of dietary protein) [
23]. The wide variation (35.00–92.00) observed in the requirements for Leu among fish species may be due to the differences in dietary protein sources, size and age of fish, genetic and species differences, feeding practices, or growth environment. Furthermore, the hybrid catfish fed diets with Leu levels above 30 g kg
−1 diet resulted in a significant reduction in growth. Similarly, excess Leu in the diet also appeared to have adverse effects on growth of other fish species [
55,
56,
57], and this was attributed to the possible antagonism between Leu and other branched chain amino acid [
58]. A previous study also demonstrated that antagonism between branched chain amino acids generally arises in animals from an excess of Leu intake over isoleucine and valine because the requirements of branched chain amino acids were affected by each other [
55]. On the other hand, the supplementation of valine in excessive Leu diets could relieve the depression of growth and feed utilization in Lake trout
Salvelinus namaycush [
59].
Skeletal muscle is the main part of the trunk of fish, accounting for about 40%–60% of body weight. Most fish muscle is composed of white muscle [
60]. Fish muscle fiber is the basic unit of skeletal muscle, which represents the development of muscle [
61]. The changes in diameter and density of white muscle has a certain guiding role in understanding the growth and development process of fish muscles. The nutritional status has been shown to affect muscle mass in mammals and teleosts [
62,
63]. The present results for the first time show that dietary Leu significantly increased the muscle fiber diameter and density in fish, indicating that Leu has beneficial effects on muscle growth and development, which were in accordance with the result for C2C12 [
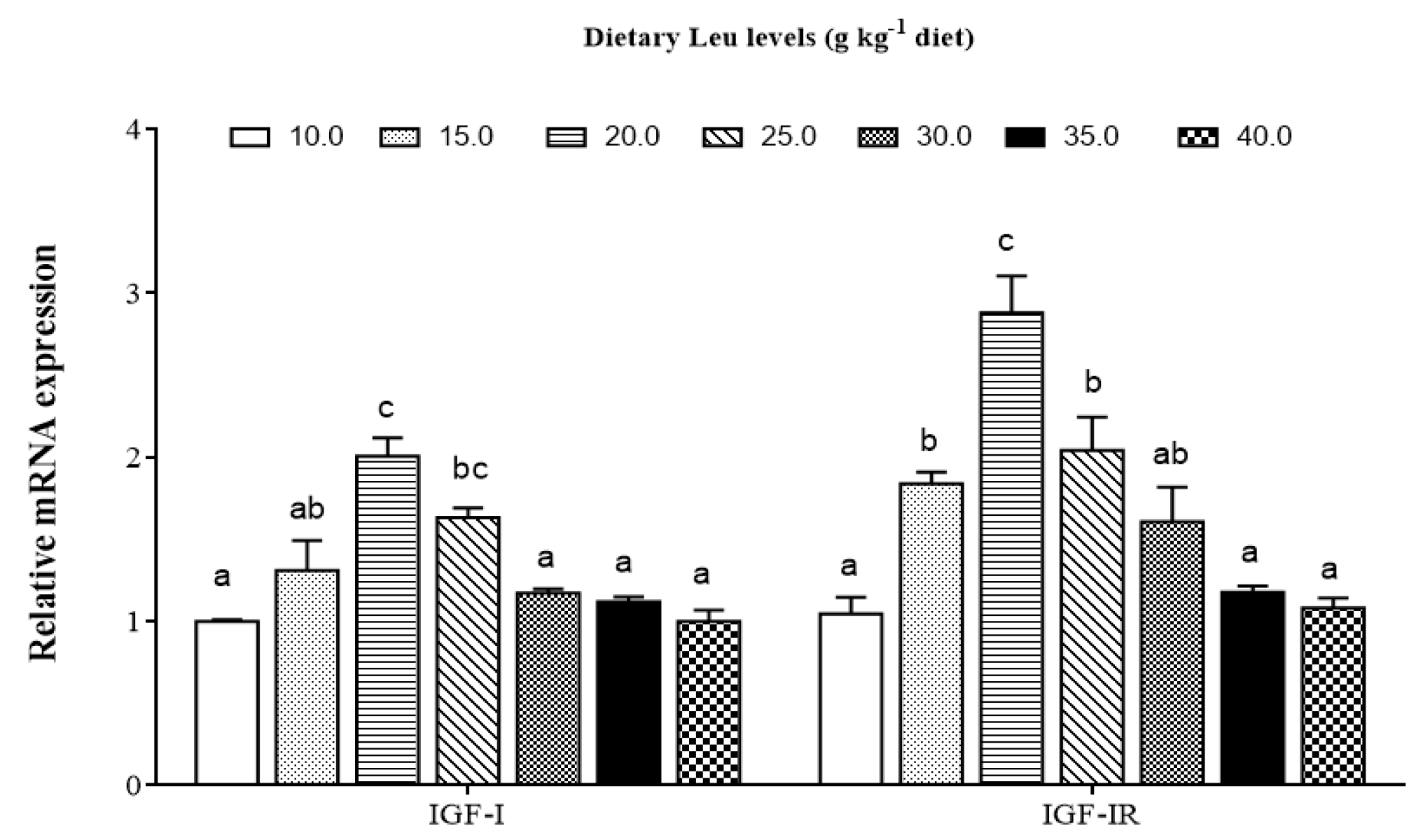
64]. The development of fish skeletal muscle is modulated by IGFs [
65]. Our data indicated that relative expressions of IGF-I and IGF-IR genes were up-regulated by Leu. Similarly, a study in gilthead sea bream cultured myocytes demonstrated that Leu deficiency significantly reduced IGF-I mRNA expression [
66]. The previous study observed Leu increased skeletal muscle IGF-I concentration in resistance-trained men [
67]. Zhou et al. also demonstrated that dietary optimal Leu elevated liver IGF-I mRNA expression [
20]. Fish muscle growth is a complicated and precisely controlled process, including proliferation and differentiation of myoblasts [
68]. The IGF-I stimulates both proliferation and differentiation of myoblasts [
69] to promote muscle growth in fish [
65,
70]. Correlation analysis indicated that the IGF-I was positively correlated with muscle fiber diameter (r = 0.724,
P = 0.066,
Table 6), indicating that dietary Leu increasing muscle fiber hypertrophy in fish might be partly related to up-regulated IGF-I transcription. Moreover, skeletal muscle is generated from satellite cells that, when activated, proliferate, fuse, and differentiate to form new myofibers. This process is also regulated by several myogenic regulatory factors [
1,
61,
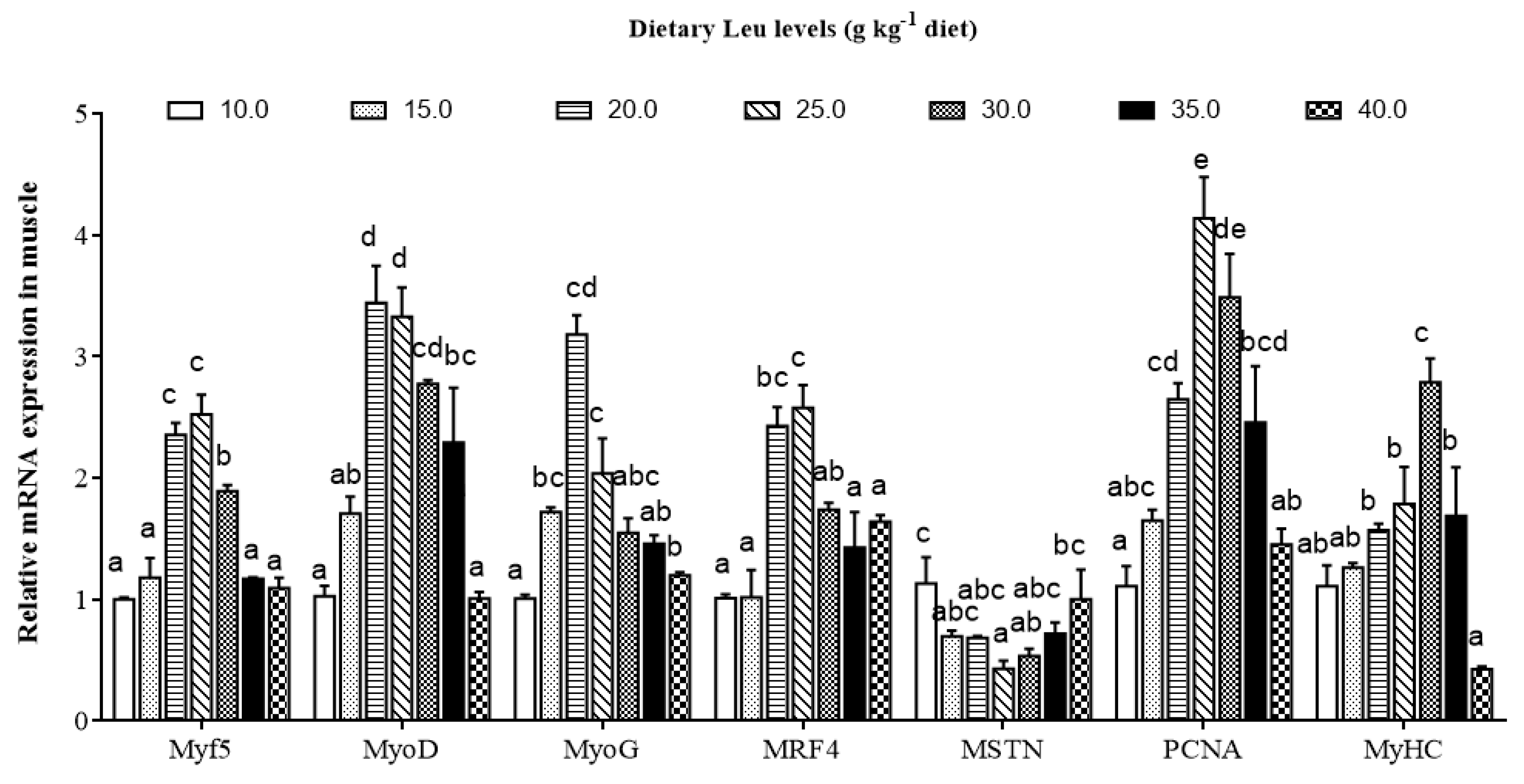
71]. During fish muscle growth, PCNA is a marker of proliferation expressed in activated satellite cells [
72]. The MyoD and Myf5 regulate the activation and proliferation of satellite cells, whereas myogenin and MRF4 act on cell differentiation [
73,
74]. The MyHC plays important roles in fish muscle growth via hyperplasia and hypertrophy of muscle fibers [
75]. The present results showed that dietary Leu up-regulated PCNA, Myf5, MyoD, MRF4, MyoG, and MyHC mRNA expressions in fish. This result was in good agreement with reports on C2C12 myoblasts, preterm rat satellite cells, and porcine myoblasts. These studies reported that Leu limitation prevents the differentiation of myoblasts and primary satellite cells [
24,
25,
26]. Chen et al. also demonstrated that Leu could promote proliferation of C2C12 myoblasts [
76]. Correlation analysis indicated that IGF-I was positively correlated with Myf5 (r = 0.837,
P = 0.019), MyoD (r = 0.820,
P = 0.024), MyoG (r = 0.977,
P = 0.000), and MRF4 (r = 0.768,
P = 0.044) mRNA levels (
Table 6), suggesting that dietary Leu increasing the muscle growth in fish might be partly related to up-regulated IGF-I transcription. MSTN, a member of the transforming growth factor-β superfamily, is expressed predominantly in skeletal muscle of fish, and the downstream function is to prevent the progression of myogenic cells into the cell division cycle [
77]. The present study showed dietary optimal Leu down-regulated MSTN mRNA expression in hybrid catfish. The result is similar to the finding in C2C12 myoblasts [
78]. The decrease of MSTN gene expression in the muscle with Leu might be related to miR-27a. Previous studies demonstrated that Leu induced proliferation promotion through miR-27a-mediated regulation of the MSTN in C2C12 cells [
76,
79]. Recently, growing evidence from fish suggests that muscle growth is regulated by essential amino acids [
80,
81,
82]. Alami-Durante et al. reported dietary methionine affected the expression of genes regulating specific transition points of myogenesis and the expression of muscle structural genes and growth factors involved in satellite cell activation and muscle growth [
81]. Michelato et al. reported that dietary histidine upregulated MyoD and MyoG mRNA expression and affected muscle hyperplasia of Nile tilapia juveniles
Oreochromis niloticus [
80]. Childress et al. also demonstrated that lysine supplementation of commercial fishmeal-free diet regulated MSTN and MyoG mRNA expression and controlled myogenesis in hybrid striped bass
Morone chrysops × M. saxatilis [
82]. These results suggested essential amino acids plays a critical role in regulating muscle growth. However, more studies are required to elucidate a more detailed mode in which Leu regulates muscle growth related gene expression in fish.
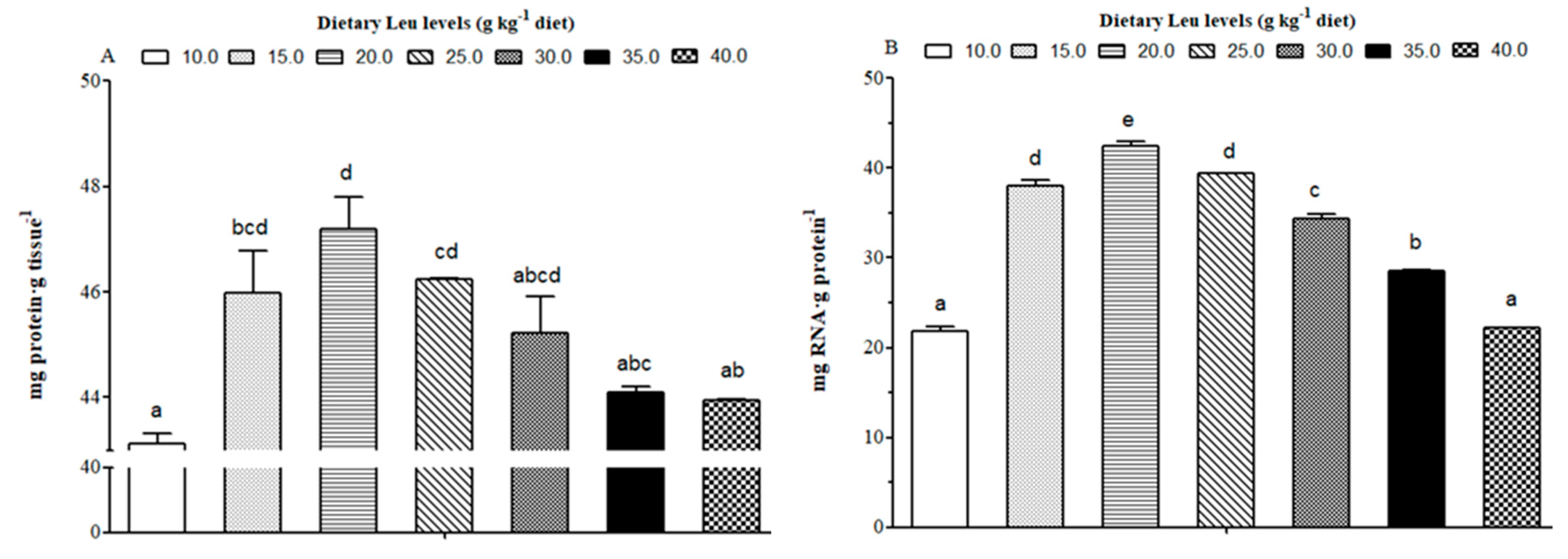
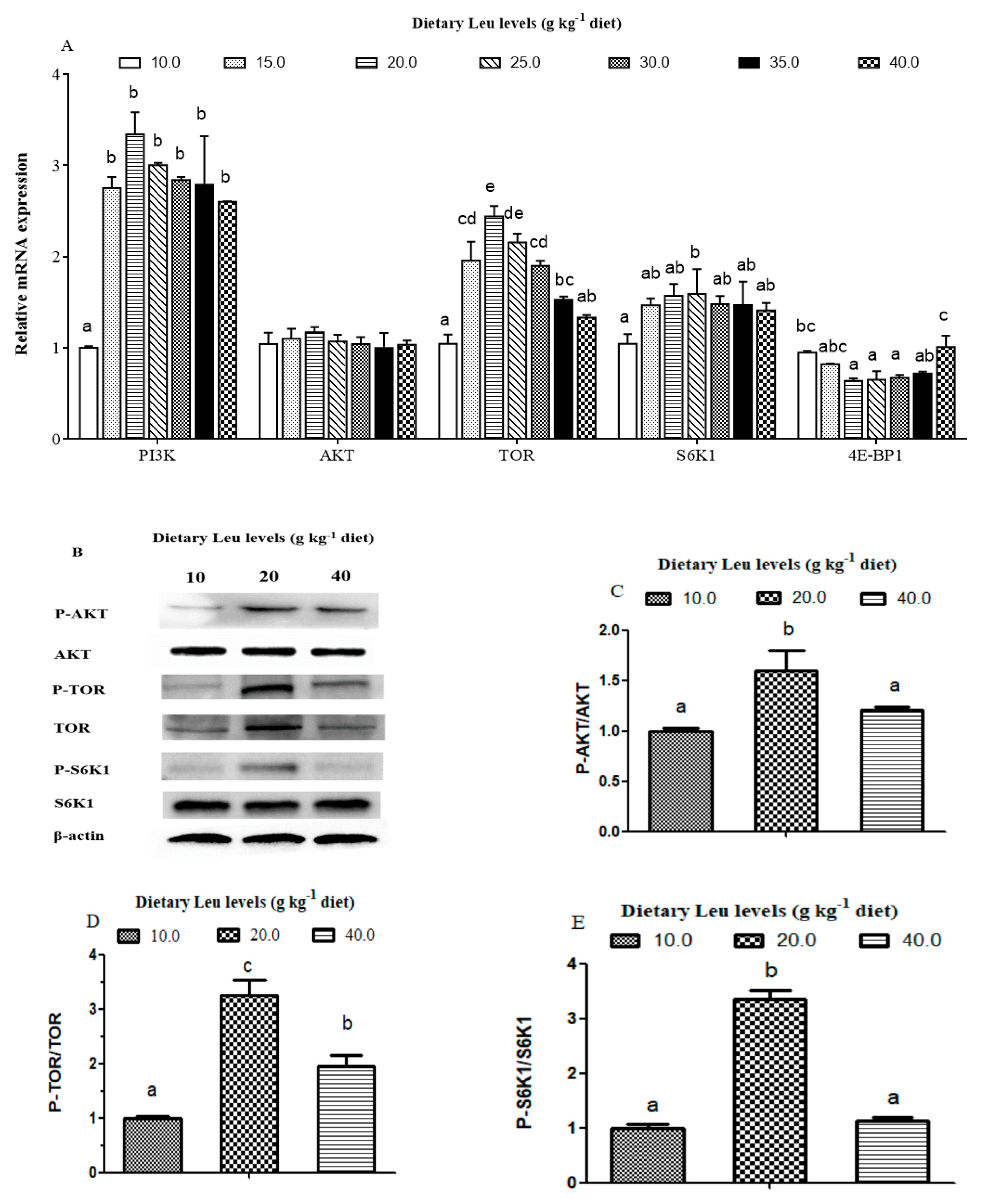
Leu is considered to be an important nutrition regulation factor involved in protein synthesis in muscle. The tissue RNA/protein ratio could reflect the capacity for protein synthesis in fish [
83]. In the present study, dietary Leu improved protein content and the RNA/protein ratio of muscle. The PI3K/AKT signaling pathway plays a crucial role in protein synthesis [
84,
85,
86]. The AKT phosphorylation is an important marker of the activation of the PI3K/AKT pathway [
8]. Despite their significance, however, their regulation by nutrients remains poorly understood in fish. The present study demonstrated dietary Leu supplementation resulted in up-regulation of PI3K mRNA levels. The phosphorylation level and phospho-total ratio of AKT were elevated in fish fed the 20 g Leu kg
−1 diet. The correlation analysis also indicated that muscle protein content was positively correlated with PI3K (r = 0.782,
P = 0.038) and AKT (r = 0.822,
P = 0.023) mRNA levels (
Table 6). These results indicated that the PI3K/AKT signaling pathway contributed to Leu-induced muscle protein synthesis. Meanwhile, the present results also showed IGF-I was positively correlated with mRNA expressions of PI3K (r = 0.637, P= 0.124) and AKT (r = 0.855,
P= 0.014), which suggested that dietary Leu promotes muscle protein synthesis by activating the PI3K/AKT signaling pathway via IGF-I. Similar results were observed in muscle of rainbow trout and fine flounder [
33,
87]. The TOR signaling pathway, a downstream component of the PI3K/AKT pathway, is necessary for stimulating translation initiation and enhancing muscle protein synthesis. Activated TOR, which has a major role in mRNA translation, promotes 4E-BP1 and S6K1 phosphorylation in fish [
88,
89]. Once phosphorylated, eukaryotic initiation factor 4E (eIF4E) is released by 4E-BP1 and then becomes the active eIF4G·eIF4E complex, which binds to mRNA and initiates translation. Phosphorylated S6K1 also takes part in translation initiation [
90]. The present data showed that dietary Leu supplementation increased muscle TOR and S6K1 mRNA levels and the phosphorylation of TOR and S6K1. The correlation analysis also indicated that muscle protein content was positively correlated with TOR (r = 0.987,
P= 0.000) and S6K1 (r = 0.791,
P= 0.034) mRNA levels, and negatively correlated with 4E-BP (r = −0.730,
P= 0.062) mRNA level (
Table 6), indicating that dietary Leu increasing muscle protein content might be partly related to elevate muscle protein synthesis via the TOR signaling pathway in fish. In mammals, Leu was proved to modulate protein synthesis directly through mTOR or indirectly through PI3K/AKT/mTOR signaling pathways [
91,
92,
93]. Lang et al. reported that TOR phosphorylation induced by Leu could be independent of the PI3K/AKT signaling pathway [
94]. These findings therefore suggest that in teleosts, as in mammals, Leu could activate the TOR or PI3K/AKT/TOR signaling pathways and contribute to muscle protein synthesis in fish. However, more studies are required to elucidate a more detailed mode in which Leu improves muscle protein synthesis in fish.
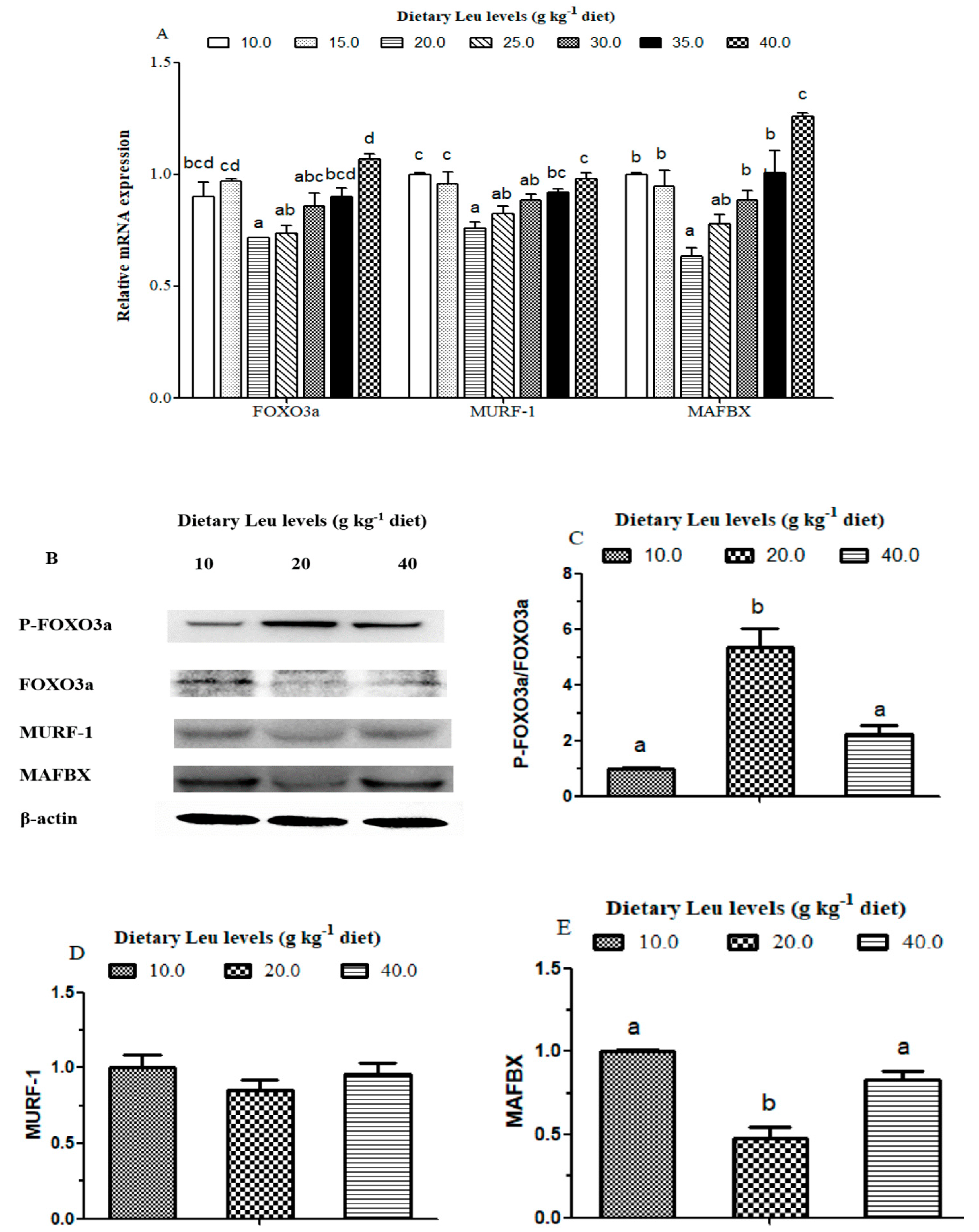
Protein degradation in mammals mainly occurs through two intracellular proteolytic systems: proteasomes and lysosomes [
95]. Most of the protein in skeletal muscle can be degraded via the UPP pathway [
39]. The AKT also appears to play a key role in protein degradation pathways, which phosphorylates and inactivates the FOXO family of transcription factors, promoting the export of FOXOs from the nucleus to the cytoplasm. The activation of AKT and inactivation of FOXO would down-regulate the activity of the UPP pathway, such as MAFBX and MURF-1 [
39], and consequently result in a decrease in protein degradation [
29,
96]. Several reports in mammals reported that Leu suppressed muscle protein degradation by the inhibition of the UPP pathway [
97,
98,
99]. In the present study, the mRNA levels of FOXO3a, MAFBX, and MURF-1 decreased for fish fed diets with increasing Leu levels up to 20 g kg
−1, suggesting that Leu may inhibit protein degradation by the UPP pathways. As expected, Western blotting results showed the phosphorylation level and phospho-total ratio of FOXO3a were elevated in fish fed the 20 g Leu kg
−1 diet. The protein levels of MAFBX were decreased in fish fed the 20 g Leu kg
-1 diet. The previous study on rainbow trout primary myocytes showed Leu reduced protein degradation by regulating MAFBX32 mRNA expression [
48]. The correlation analysis also indicated that muscle protein content was negatively correlated with FOXO3a (r = −0.649,
P= 0.115), MURF-1 (r = −0.838,
P= 0.019), and MAFBX (r = −0.808,
P= 0.028) mRNA levels (
Table 6), which suggested that the dietary Leu attenuated protein degradation was partly related to AKT/FOXO3a signaling pathway. To our best knowledge, this is the first report demonstrating that Leu could promote protein deposition by preventing protein degradation via the AKT/FOXO3a signaling pathway in fish. In addition, increasing evidence indicated that IGF-I could inhibit overall protein breakdown, degradation of myofibrillar proteins, and expression of MURF-1 [
100,
101]. Correlation analysis indicated that IGF-I was negatively correlated with FOXO3a (r = −0.809,
P = 0.028), MURF-1 (r = −0.923,
P = 0.003), and MAFBX (r = −0.883,
P= 0.008) mRNA levels (
Table 6), indicating that dietary Leu decreasing the muscle protein degradation in fish might be partly related to up-regulated IGF-I transcription. Taken together, these data indicated that Leu prevented protein degradation via the IGF-I/AKT/FOXO3a signaling pathway. To our knowledge, no study has focused on the effects of Leu on protein degradation related to the IGF-I/AKT/FOXO3a signaling pathway in fish.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}