Fatty Acid Oxidation and Mitochondrial Morphology Changes as Key Modulators of the Affinity for ADP in Rat Heart Mitochondria

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
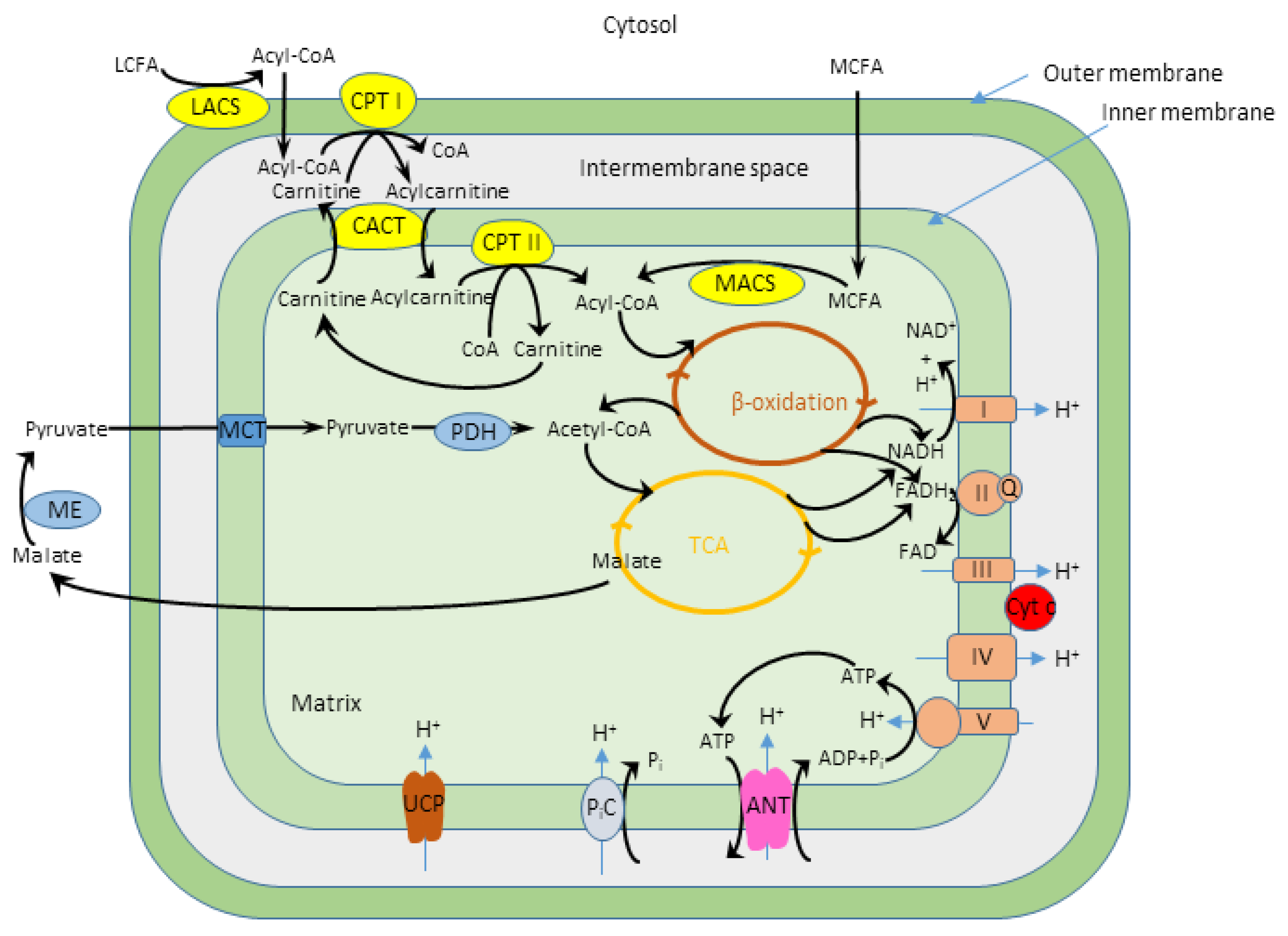
:1. Introduction
2. Materials and Methods
3. Results
3.1. Role of Oxidation of Fatty Acids in Regulation of Oxidative Phosphorylation and Mitochondrial Swelling
3.2. Regulation of Mitochondrial Respiration by Endogenous versus Exogenous ADP: Influence of Palmitoyl-l-Carnitine
3.3. Influence of Palmitoyl-l-Carnitine on the Permeability of Mitochondrial Outer Membrane for ADP
3.4. Influence of Creatine Kinase on Regulation of Mitochondrial Respiration Supported by Fatty Acids
3.5. Effects of Oncotic Pressure on Mitochondrial Fatty Acid Oxidation
3.6. Fatty Acid-Oxidation and Oncotic Pressure-induced Changes of Mitochondrial Morphology
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vignais, P.V. Molecular and physiological aspects of adenine nucleotide transport in mitochondria. Biochim. Biophys. Acta 1976, 456, 1–38. [Google Scholar] [CrossRef]
- Wojtczak, L. Effect of long-chain fatty acids and acyl-CoA on mitochondrial permeability, transport, and energy-coupling processes. J. Bioenerg. Biomembr. 1976, 8, 293–311. [Google Scholar] [PubMed]
- Qu, Q.; Zeng, F.; Liu, X.; Wang, Q.J.; Deng, F. Fatty acid oxidation and carnitine palmitoyltransferase I: Emerging therapeutic targets in cancer. Cell Death Dis. 2016, 7, e2226. [Google Scholar] [CrossRef] [PubMed]
- Roussel, J.; Thireau, J.; Brenner, C.; Saint, N.; Scheuermann, V.; Lacampagne, A.; Le Guennec, J.Y.; Fauconnier, J. Palmitoyl-carnitine increases RyR2 oxidation and sarcoplasmic reticulum Ca2+ leak in cardiomyocytes: Role of adenine nucleotide translocase. Biochim. Biophys. Acta 2015, 1852, 749–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopaschuk, G.D.; Ussher, J.R.; Folmes, C.D.; Jaswal, J.S.; Stanley, W.C. Myocardial fatty acid metabolism in health and disease. Physiol. Rev. 2010, 90, 207–258. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, M. The ADP and ATP transport in mitochondria and its carrier. Biochim. Biophys. Acta 2008, 1778, 1978–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saks, V.A.; Khuchua, Z.A.; Vasilyeva, E.V.; Belikova, O.; Kuznetsov, A.V. Metabolic compartmentation and substrate channelling in muscle cells. Role of coupled creatine kinases in in vivo regulation of cellular respiration—A synthesis. Mol. Cell Biochem. 1994, 133–134, 155–192. [Google Scholar] [CrossRef]
- Seppet, E.K.; Kaambre, T.; Sikk, P.; Tiivel, T.; Vija, H.; Tonkonogi, M.; Sahlin, K.; Kay, L.; Appaix, F.; Braun, U.; et al. Functional complexes of mitochondria with Ca,MgATPases of myofibrils and sarcoplasmic reticulum in muscle cells. Biochim. Biophys. Acta 2001, 1504, 379–395. [Google Scholar] [CrossRef] [Green Version]
- Toleikis, A.; Liobikas, J.; Trumbeckaite, S.; Majiene, D. Relevance of fatty acid oxidation in regulation of the outer mitochondrial membrane permeability for ADP. FEBS Lett. 2001, 509, 245–249. [Google Scholar] [CrossRef] [Green Version]
- Saks, V.A.; Kaambre, T.; Sikk, P.; Eimre, M.; Orlova, E.; Paju, K.; Piirsoo, A.; Appaix, F.; Kay, L.; Regitz-Zagrosek, V.; et al. Intracellular energetic units in red muscle cells. Biochem. J. 2001, 356, 643–657. [Google Scholar] [CrossRef]
- Saks, V.; Kuznetsov, A.V.; Gonzalez-Granillo, M.; Tepp, K.; Timohhina, N.; Karu-Varikmaa, M.; Kaambre, T.; Dos Santos, P.; Boucher, F.; Guzun, R. Intracellular Energetic Units regulate metabolism in cardiac cells. J. Mol. Cell Cardiol. 2012, 52, 419–436. [Google Scholar] [CrossRef] [PubMed]
- Dzeja, P.P.; Hoyer, K.; Tian, R.; Zhang, S.; Nemutlu, E.; Spindler, M.; Ingwall, J.S. Rearrangement of energetic and substrate utilization networks compensate for chronic myocardial creatine kinase deficiency. J. Physiol. 2011, 589, 5193–5211. [Google Scholar] [CrossRef] [PubMed]
- Kaasik, A.; Veksler, V.; Boehm, E.; Novotova, M.; Minajeva, A.; Ventura-Clapier, R. Energetic crosstalk between organelles: Architectural integration of energy production and utilization. Circ. Res. 2001, 89, 153–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toleikis, A.; Majiene, D.; Trumbeckaite, S.; Dagys, A.; Jasaitis, A. The effect of collagenase and temperature on mitochondrial respiratory parameters in saponin-skinned cardiac fibers. Biosci. Rep. 1996, 16, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Liobikas, J.; Kopustinskiene, D.M.; Toleikis, A. What controls the outer mitochondrial membrane permeability for ADP: Facts for and against the role of oncotic pressure. Biochim. Biophys. Acta 2001, 1505, 220–225. [Google Scholar] [CrossRef] [Green Version]
- Appaix, F.; Kuznetsov, A.V.; Usson, Y.; Kay, L.; Andrienko, T.; Olivares, J.; Kaambre, T.; Sikk, P.; Margreiter, R.; Saks, V. Possible role of cytoskeleton in intracellular arrangement and regulation of mitochondria. Exp. Physiol. 2003, 88, 175–190. [Google Scholar] [CrossRef] [Green Version]
- Guzun, R.; Timohhina, N.; Tepp, K.; Monge, C.; Kaambre, T.; Sikk, P.; Kuznetsov, A.V.; Pison, C.; Saks, V. Regulation of respiration controlled by mitochondrial creatine kinase in permeabilized cardiac cells in situ. Importance of system level properties. Biochim. Biophys. Acta 2009, 1787, 1089–1105. [Google Scholar] [CrossRef]
- Boudina, S.; Laclau, M.N.; Tariosse, L.; Daret, D.; Gouverneur, G.; Bonoron-Adele, S.; Saks, V.A.; Dos Santos, P. Alteration of mitochondrial function in a model of chronic ischemia in vivo in rat heart. Am. J. Physiol. Heart Circ. Physiol. 2002, 282, H821–H831. [Google Scholar] [CrossRef] [Green Version]
- Divakaruni, A.S.; Humphrey, D.M.; Brand, M.D. Fatty acids change the conformation of uncoupling protein 1 (UCP1). J. Biol. Chem. 2012, 287, 36845–36853. [Google Scholar] [CrossRef] [Green Version]
- Aquila, H.; Link, T.A.; Klingenberg, M. The uncoupling protein from brown fat mitochondria is related to the mitochondrial ADP/ATP carrier. Analysis of sequence homologies and of folding of the protein in the membrane. EMBO J. 1985, 4, 2369–2376. [Google Scholar] [CrossRef]
- Klingenberg, M. Mechanism and evolution of the uncoupling protein of brown adipose tissue. Trends Biochem. Sci. 1990, 15, 108–112. [Google Scholar] [CrossRef]
- Anmann, T.; Guzun, R.; Beraud, N.; Pelloux, S.; Kuznetsov, A.V.; Kogerman, L.; Kaambre, T.; Sikk, P.; Paju, K.; Peet, N.; et al. Different kinetics of the regulation of respiration in permeabilized cardiomyocytes and in HL-1 cardiac cells. Importance of cell structure/organization for respiration regulation. Biochim. Biophys. Acta 2006, 1757, 1597–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eimre, M.; Paju, K.; Pelloux, S.; Beraud, N.; Roosimaa, M.; Kadaja, L.; Gruno, M.; Peet, N.; Orlova, E.; Remmelkoor, R.; et al. Distinct organization of energy metabolism in HL-1 cardiac cell line and cardiomyocytes. Biochim. Biophys. Acta 2008, 1777, 514–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anmann, T.; Varikmaa, M.; Timohhina, N.; Tepp, K.; Shevchuk, I.; Chekulayev, V.; Saks, V.; Kaambre, T. Formation of highly organized intracellular structure and energy metabolism in cardiac muscle cells during postnatal development of rat heart. Biochim. Biophys. Acta 2014, 1837, 1350–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valcarce, C.; Navarrete, R.M.; Encabo, P.; Loeches, E.; Satrustegui, J.; Cuezva, J.M. Postnatal development of rat liver mitochondrial functions. The roles of protein synthesis and of adenine nucleotides. J. Biol. Chem. 1988, 263, 7767–7775. [Google Scholar] [PubMed]
- Valcarce, C.; Cuezva, J.M. Interaction of adenine nucleotides with the adenine nucleotide translocase regulates the developmental changes in proton conductance of the inner mitochondrial membrane. FEBS Lett. 1991, 294, 225–228. [Google Scholar] [CrossRef] [Green Version]
- Benard, G.; Faustin, B.; Passerieux, E.; Galinier, A.; Rocher, C.; Bellance, N.; Delage, J.P.; Casteilla, L.; Letellier, T.; Rossignol, R. Physiological diversity of mitochondrial oxidative phosphorylation. Am. J. Physiol. Cell Physiol. 2006, 291, C1172–C1182. [Google Scholar] [CrossRef]
- Fernandez-Vizarra, E.; Enriquez, J.A.; Perez-Martos, A.; Montoya, J.; Fernandez-Silva, P. Tissue-specific differences in mitochondrial activity and biogenesis. Mitochondrion 2011, 11, 207–213. [Google Scholar] [CrossRef]
- Kuznetsov, A.V.; Veksler, V.; Gellerich, F.N.; Saks, V.; Margreiter, R.; Kunz, W.S. Analysis of mitochondrial function in situ in permeabilized muscle fibers, tissues and cells. Nat. Protoc. 2008, 3, 965–976. [Google Scholar] [CrossRef]
- Holtzman, J.L. Calibration of the oxygen polarograph by the depletion of oxygen with hypoxanthine-xanthine oxidase-catalase. Anal. Chem. 1976, 48, 229–230. [Google Scholar] [CrossRef]
- Gornall, A.G.; Bardawill, C.J.; David, M.M. Determination of serum proteins by means of the biuret reaction. J. Biol. Chem. 1949, 177, 751–766. [Google Scholar] [PubMed]
- Sultan, A.; Sokolove, P.M. Free fatty acid effects on mitochondrial permeability: An overview. Arch. Biochem. Biophys. 2001, 386, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Kerner, J.; Hoppel, C. Fatty acid import into mitochondria. Biochim. Biophys. Acta 2000, 1486, 1–17. [Google Scholar] [CrossRef]
- Levitsky, D.O.; Skulachev, V.P. Carnitine: The carrier transporting fatty acyls into mitochondria by means of an electrochemical gradient of H+. Biochim. Biophys. Acta 1972, 275, 33–50. [Google Scholar] [CrossRef]
- Neely, J.R.; Feuvray, D. Metabolic products and myocardial ischemia. Am. J. Pathol. 1981, 102, 282–291. [Google Scholar] [PubMed]
- Piper, M.H.; Sezer, O.; Schwartz, P.; Hutter, J.F.; Schweickhardt, C.; Spieckermann, P.G. Acyl-carnitine effects on isolated cardiac mitochondria and erythrocytes. Basic Res. Cardiol. 1984, 79, 186–198. [Google Scholar] [CrossRef]
- Goni, F.M.; Requero, M.A.; Alonso, A. Palmitoylcarnitine, a surface-active metabolite. FEBS Lett. 1996, 390, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Tuunanen, H.; Engblom, E.; Naum, A.; Nagren, K.; Hesse, B.; Airaksinen, K.E.; Nuutila, P.; Iozzo, P.; Ukkonen, H.; Opie, L.H.; et al. Free fatty acid depletion acutely decreases cardiac work and efficiency in cardiomyopathic heart failure. Circulation 2006, 114, 2130–2137. [Google Scholar] [CrossRef] [Green Version]
- Taegtmeyer, H. Metabolism--the lost child of cardiology. J. Am. Coll. Cardiol. 2000, 36, 1386–1388. [Google Scholar] [CrossRef] [Green Version]
- Laclau, M.N.; Boudina, S.; Thambo, J.B.; Tariosse, L.; Gouverneur, G.; Bonoron-Adele, S.; Saks, V.A.; Garlid, K.D.; Dos Santos, P. Cardioprotection by ischemic preconditioning preserves mitochondrial function and functional coupling between adenine nucleotide translocase and creatine kinase. J. Mol. Cell Cardiol. 2001, 33, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Kuznetsov, A.V.; Tiivel, T.; Sikk, P.; Kaambre, T.; Kay, L.; Daneshrad, Z.; Rossi, A.; Kadaja, L.; Peet, N.; Seppet, E.; et al. Striking differences between the kinetics of regulation of respiration by ADP in slow-twitch and fast-twitch muscles in vivo. Eur. J. Biochem. 1996, 241, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Gellerich, F.N.; Schlame, M.; Bohnensack, R.; Kunz, W. Dynamic compartmentation of adenine nucleotides in the mitochondrial intermembrane space of rat-heart mitochondria. Biochim. Biophys. Acta 1987, 890, 117–126. [Google Scholar] [CrossRef]
- Saks, V.A.; Chernousova, G.B.; Gukovsky, D.E.; Smirnov, V.N.; Chazov, E.I. Studies of energy transport in heart cells. Mitochondrial isoenzyme of creatine phosphokinase: Kinetic properties and regulatory action of Mg2+ ions. Eur. J. Biochem. 1975, 57, 273–290. [Google Scholar] [CrossRef] [PubMed]
- Jacobus, W.E.; Saks, V.A. Creatine kinase of heart mitochondria: Changes in its kinetic properties induced by coupling to oxidative phosphorylation. Arch. Biochem. Biophys. 1982, 219, 167–178. [Google Scholar] [CrossRef]
- Meyer, R.A.; Sweeney, H.L.; Kushmerick, M.J. A simple analysis of the “phosphocreatine shuttle”. Am. J. Physiol. 1984, 246, C365–C377. [Google Scholar] [CrossRef]
- Fulton, A.B. How crowded is the cytoplasm? Cell 1982, 30, 345–347. [Google Scholar] [CrossRef]
- Bakeeva, L.E.; Chentsov, Y.S.; Jasaitis, A.A.; Skulachev, V.P. The effect of oncotic pressure on heart muscle mitochondria. Biochim. Biophys. Acta 1972, 275, 319–332. [Google Scholar] [CrossRef]
- Wicker, U.; Bucheler, K.; Gellerich, F.N.; Wagner, M.; Kapischke, M.; Brdiczka, D. Effect of macromolecules on the structure of the mitochondrial inter-membrane space and the regulation of hexokinase. Biochim. Biophys. Acta 1993, 1142, 228–239. [Google Scholar] [CrossRef]
- Wrogemann, K.; Nylen, E.G.; Adamson, I.; Pande, S.V. Functional studies on in situ-like mitochondria isolated in the presence of polyvinyl pyrrolidone. Biochim. Biophys. Acta 1985, 806, 1–8. [Google Scholar] [CrossRef]
- Gellerich, F.N.; Laterveer, F.D.; Korzeniewski, B.; Zierz, S.; Nicolay, K. Dextran strongly increases the Michaelis constants of oxidative phosphorylation and of mitochondrial creatine kinase in heart mitochondria. Eur. J. Biochem. 1998, 254, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Gellerich, F.N.; Wagner, M.; Kapischke, M.; Wicker, U.; Brdiczka, D. Effect of macromolecules on the regulation of the mitochondrial outer membrane pore and the activity of adenylate kinase in the inter-membrane space. Biochim. Biophys. Acta 1993, 1142, 217–227. [Google Scholar] [CrossRef]
- Gellerich, F.N.; Kapischke, M.; Kunz, W.; Neumann, W.; Kuznetsov, A.; Brdiczka, D.; Nicolay, K. The influence of the cytosolic oncotic pressure on the permeability of the mitochondrial outer membrane for ADP: Implications for the kinetic properties of mitochondrial creatine kinase and for ADP channelling into the intermembrane space. Mol. Cell Biochem. 1994, 133–134, 85–104. [Google Scholar] [CrossRef] [PubMed]
- Nohl, H. Age-dependent changes in the structure-function correlation of ADP/ATP-translocating mitochondrial membranes. Gerontology 1982, 28, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Lizana, L.; Bauer, B.; Orwar, O. Controlling the rates of biochemical reactions and signaling networks by shape and volume changes. Proc. Natl. Acad. Sci. USA 2008, 105, 4099–4104. [Google Scholar] [CrossRef] [Green Version]
- Cogliati, S.; Enriquez, J.A.; Scorrano, L. Mitochondrial Cristae: Where Beauty Meets Functionality. Trends Biochem. Sci. 2016, 41, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Seifert, E.L.; Estey, C.; Xuan, J.Y.; Harper, M.E. Electron transport chain-dependent and -independent mechanisms of mitochondrial H2O2 emission during long-chain fatty acid oxidation. J. Biol. Chem. 2010, 285, 5748–5758. [Google Scholar] [CrossRef] [Green Version]
- St-Pierre, J.; Buckingham, J.A.; Roebuck, S.J.; Brand, M.D. Topology of superoxide production from different sites in the mitochondrial electron transport chain. J. Biol. Chem. 2002, 277, 44784–44790. [Google Scholar] [CrossRef] [Green Version]
- Liesa, M.; Shirihai, O.S. Mitochondrial dynamics in the regulation of nutrient utilization and energy expenditure. Cell Metab. 2013, 17, 491–506. [Google Scholar] [CrossRef] [Green Version]
- Allouche, M.; Pertuiset, C.; Robert, J.L.; Martel, C.; Veneziano, R.; Henry, C.; dein, O.S.; Saint, N.; Brenner, C.; Chopineau, J. ANT-VDAC1 interaction is direct and depends on ANT isoform conformation in vitro. Biochem. Biophys. Res. Commun. 2012, 429, 12–17. [Google Scholar] [CrossRef]
- Bernardi, P. Mitochondrial H+ permeability through the ADP/ATP carrier. Nat. Metab. 2019, 1, 752–753. [Google Scholar] [CrossRef]
- Bertholet, A.M.; Chouchani, E.T.; Kazak, L.; Angelin, A.; Fedorenko, A.; Long, J.Z.; Vidoni, S.; Garrity, R.; Cho, J.; Terada, N.; et al. H(+) transport is an integral function of the mitochondrial ADP/ATP carrier. Nature 2019, 571, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Kramer, R.; Klingenberg, M. Electrophoretic control of reconstituted adenine nucleotide translocation. Biochemistry 1982, 21, 1082–1089. [Google Scholar] [CrossRef] [PubMed]
- Heerdt, B.G.; Houston, M.A.; Augenlicht, L.H. Growth properties of colonic tumor cells are a function of the intrinsic mitochondrial membrane potential. Cancer Res. 2006, 66, 1591–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, S.; Dzeja, P.P.; Faustino, R.S.; Perez-Terzic, C.; Behfar, A.; Terzic, A. Mitochondrial oxidative metabolism is required for the cardiac differentiation of stem cells. Nat. Clin. Pr. Cardiovasc. Med. 2007, 4, S60–S67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricquier, D. Mitochondrial uncoupling proteins. Curr. Opin. Drug Discov. Devel. 1999, 2, 497–504. [Google Scholar] [PubMed]
- Porporato, P.E.; Filigheddu, N.; Pedro, J.M.B.; Kroemer, G.; Galluzzi, L. Mitochondrial metabolism and cancer. Cell Res. 2018, 28, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Shammas, M.A.; Neri, P.; Koley, H.; Batchu, R.B.; Bertheau, R.C.; Munshi, V.; Prabhala, R.; Fulciniti, M.; Tai, Y.T.; Treon, S.P.; et al. Specific killing of multiple myeloma cells by (-)-epigallocatechin-3-gallate extracted from green tea: Biologic activity and therapeutic implications. Blood 2006, 108, 2804–2810. [Google Scholar] [CrossRef]
- Braicu, C.; Ladomery, M.R.; Chedea, V.S.; Irimie, A.; Berindan-Neagoe, I. The relationship between the structure and biological actions of green tea catechins. Food Chem. 2013, 141, 3282–3289. [Google Scholar] [CrossRef]
- Majumder, K.; Mine, Y.; Wu, J. The potential of food protein-derived anti-inflammatory peptides against various chronic inflammatory diseases. J. Sci. Food Agric. 2016, 96, 2303–2311. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Gan, R.Y.; Li, S.; Zhou, Y.; Li, A.N.; Xu, D.P.; Li, H.B. Antioxidant Phytochemicals for the Prevention and Treatment of Chronic Diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef]











© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toleikis, A.; Trumbeckaite, S.; Liobikas, J.; Pauziene, N.; Kursvietiene, L.; Kopustinskiene, D.M. Fatty Acid Oxidation and Mitochondrial Morphology Changes as Key Modulators of the Affinity for ADP in Rat Heart Mitochondria. Cells 2020, 9, 340. https://doi.org/10.3390/cells9020340
Toleikis A, Trumbeckaite S, Liobikas J, Pauziene N, Kursvietiene L, Kopustinskiene DM. Fatty Acid Oxidation and Mitochondrial Morphology Changes as Key Modulators of the Affinity for ADP in Rat Heart Mitochondria. Cells. 2020; 9(2):340. https://doi.org/10.3390/cells9020340
Chicago/Turabian StyleToleikis, Adolfas, Sonata Trumbeckaite, Julius Liobikas, Neringa Pauziene, Lolita Kursvietiene, and Dalia M. Kopustinskiene. 2020. "Fatty Acid Oxidation and Mitochondrial Morphology Changes as Key Modulators of the Affinity for ADP in Rat Heart Mitochondria" Cells 9, no. 2: 340. https://doi.org/10.3390/cells9020340