TLR2 Signaling Pathway Combats Streptococcus uberis Infection by Inducing Mitochondrial Reactive Oxygen Species Production

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain, Cell Culture, and Treatment
2.2. Mice and Treatment
2.3. Histological Observation and Immunohistochemistry
2.4. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.5. Total Protein Extraction and Western Blotting
2.6. Measurement of Reactive Oxygen Species (ROS) and Mitochondrial Reactive Oxygen Species (mROS)
2.7. Assay of TNF-α, IL-1β, and IL-6 by ELISA
2.8. Detection of NAGase, Total Antioxidant Capacity (T-AOC), Superoxide Dismutase (SOD), Malondialdehyde (MDA), and Uncoupling Protein 2 (UCP2)
2.9. Viable Bacterial Count Assay
2.10. Statistical Analysis
3. Results
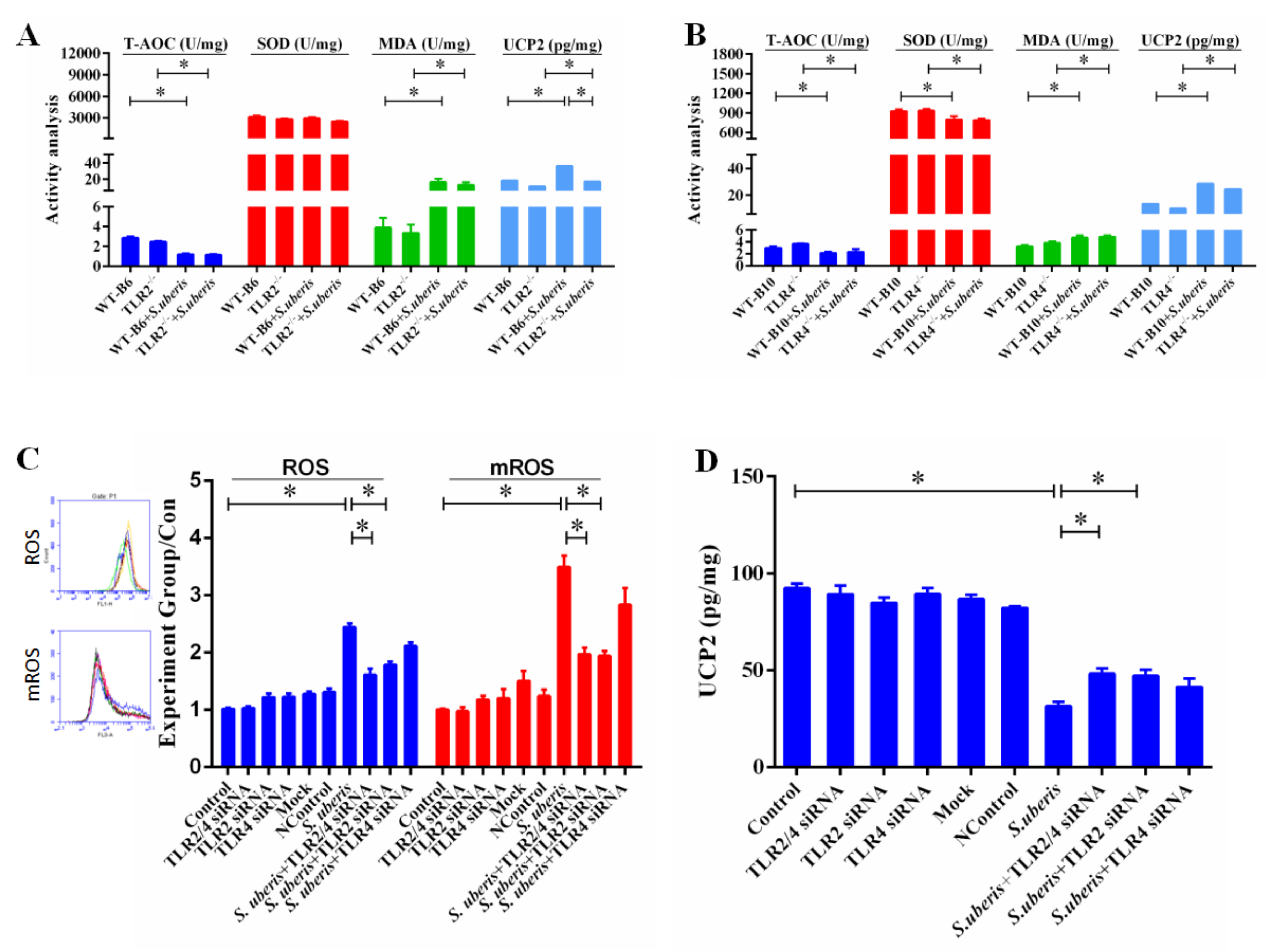
3.1. TLR2 Mediates Tissue Damage and Anti-S. uberis Infection in Mammary Glands
3.2. TLR2 and TLR4 Deficiencies Affect the Secretion of Cytokines in S.uberis Infection
3.3. MyD88-Dependent Pathway Predominates in S. uberis Infection
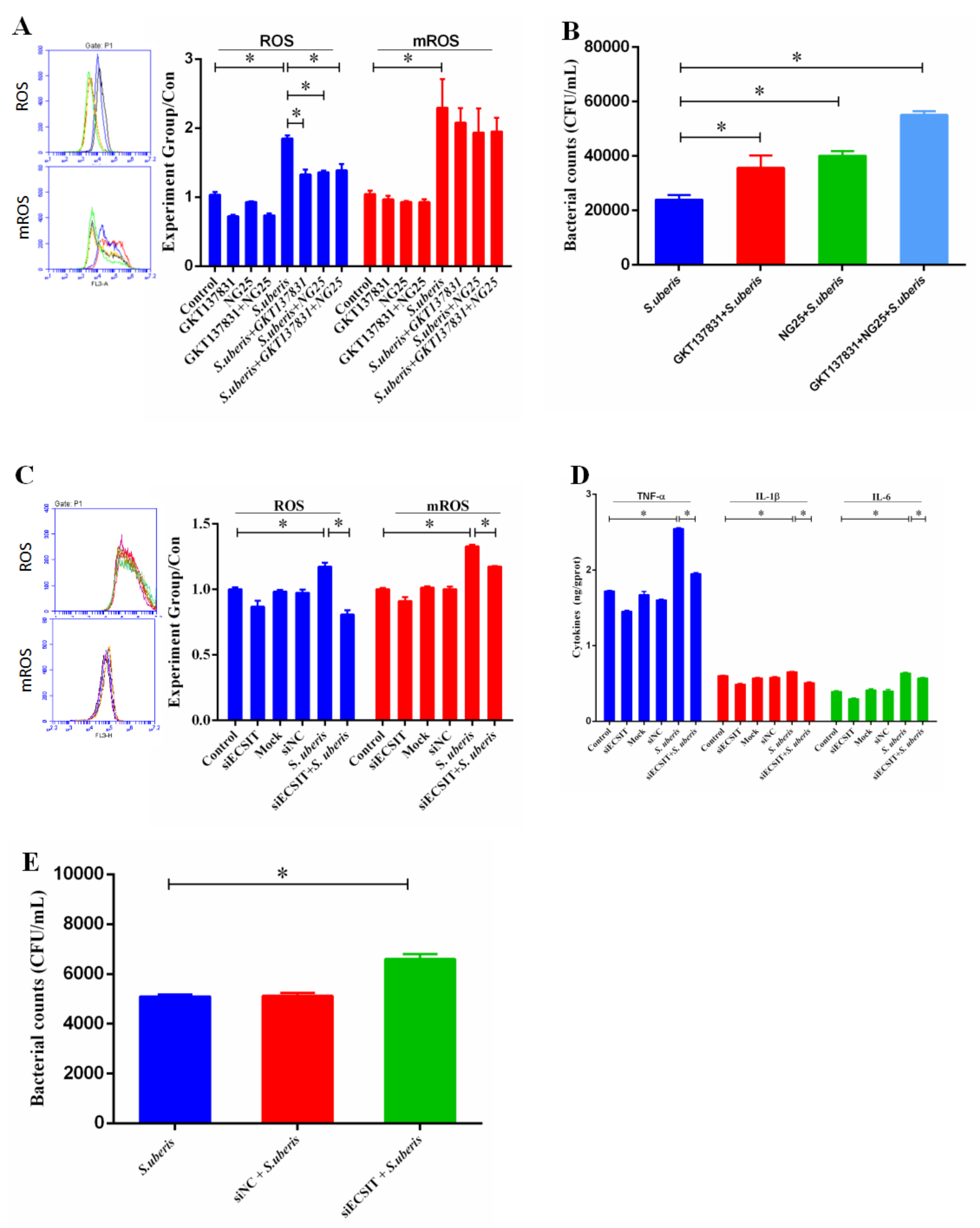
3.4. TRAF6 and ECSIT Participate in Signal Sensing from Toll-Like Receptors (TLRs) in S. uberis Infection
3.5. TLRs Mediate Redox Status in Mammary Glands During S. uberis Infection
3.6. mROS Plays an Important Role Against S. uberis Infection in Mammary Epithelial Cells (MECs)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bergonier, D.; de Crémoux, R.; Rupp, R.; Lagriffoul, G.; Berthelot, X. Mastitis of dairy small ruminants. Vet. Res. 2003, 34, 689–716. [Google Scholar] [CrossRef] [Green Version]
- Rocco-Machado, N.; Cosentino-Gomes, D.; Nascimento, M.T.; Paes-Vieira, L.; Khan, Y.A.; Mittra, B.; Andrews, N.W.; Meyer-Fernandes, J.R. Leishmania amazonensis ferric iron reductase (LFR1) is a bifunctional enzyme: Unveiling a NADPH oxidase activity. Free Radic. Biol. Med. 2019, 143, 341–353. [Google Scholar] [CrossRef]
- Kessel, S.; Wittenberg, C.E. Die Gelenkinfektion des jungen Menschen durch Streptococcus uberis, ein Erreger der Mastitis beim Rind—eine Einzelfalldarstellung. Z. Orthop. Unfall. 2008, 146, 507–509. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Zhang, J.; Ma, Z.; Zheng, L. The role of NADPH oxidase in taurine attenuation of Streptococcus uberis-induced mastitis in rats. Int. Immunopharmacol. 2013, 16, 429–435. [Google Scholar] [CrossRef]
- Strandberg, Y.; Gray, C.; Vuocolo, T.; Donaldson, L.; Broadway, M.; Tellam, R. Lipopolysaccharide and lipoteichoic acid induce different innate immune responses in bovine mammary epithelial cells. Cytokine 2005, 31, 72–86. [Google Scholar] [CrossRef]
- Zanoni, I.; Granucci, F. Regulation and dysregulation of innate immunity by NFAT signaling downstream of pattern recognition receptors (PRRs). Eur. J. Immunol. 2012, 42, 1924–1931. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. TLR signaling. Semin. Immunol. 2007, 19, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, J.; Laganière, J.; Mehl, I.R.; Barish, G.D.; Chong, L.W.; Li, X.; Scheffler, I.E.; Mock, D.C.; Bataille, A.R.; Robert, F.; et al. Nuclear receptor ERR alpha and coactivator PGC-1 beta are effectors of IFN-gamma-induced host defense. Genes Dev. 2007, 21, 1909–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibeagha-Awemu., E.M.; Lee, J.W.; Ibeagha, A.E.; Bannerman, D.D.; Paape, M.J.; Zhao, X. Bacterial lipopolysaccharide induces increased expression of toll-like receptor (TLR) 4 and downstream TLR signaling molecules in bovine mammary epithelial cells. Vet. Res. 2008, 39, 11. [Google Scholar] [CrossRef] [Green Version]
- Dai, B.; Zhang, J.; Liu, M.; Lu, J.; Zhang, Y.; Xu, Y.; Miao, J.; Yin, Y. The role of Ca (2+) mediated signaling pathways on the effect of taurine against Streptococcus uberis infection. Vet. Microbiol. 2016, 192, 26–33. [Google Scholar] [CrossRef]
- Li, B.; Xi, P.; Wang, Z.; Han, X.; Xu, Y.; Zhang, Y.; Miao, J. PI3K/Akt/mTOR signaling pathway participates in Streptococcus uberis-induced inflammation in mammary epithelial cells in concert with the classical TLRs/NF-ĸB pathway. Vet. Microbiol. 2018, 227, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Zheng, L.; Zhang, J.; Ma, Z.; Zhu, W.; Zou, S. The effect of taurine on the toll-like receptors/nuclear factor kappa B (TLRs/NF-κB) signaling pathway in Streptococcus uberis-induced mastitis in rats. Int. Immunopharmacol. 2011, 11, 1740–1746. [Google Scholar] [CrossRef]
- Liu, B.; Cheng, Y.; Zhang, B.; Bian, H.J.; Bao, J.K. Polygonatum cyrtonema lectin induces apoptosis and autophagy in human melanoma A375 cells through a mitochondria-mediated ROS-p38-p53 pathway. Cancer Lett. 2009, 275, 54–60. [Google Scholar] [CrossRef] [PubMed]
- West, A.P.; Brodsky, I.E.; Rahner, C.; Woo, D.K.; Erdjument-Bromage, H.; Tempst, P.; Walsh, M.C.; Choi, Y.; Shadel, G.S.; Ghosh, S. TLR signalling augments macrophage bactericidal activity through mitochondrial ROS. Nature 2011, 472, 476–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaser, H.; Dostert, C.; Mak, T.W.; Brenner, D. TNF and ROS Crosstalk in Inflammation. Trends Cell Biol. 2016, 26, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Hughes, G.; Murphy, M.P.; Ledgerwood, E.C. Mitochondrial reactive oxygen species regulate the temporal activation of nuclear factor kappaB to modulate tumour necrosis factor-induced apoptosis: Evidence from mitochondria-targeted antioxidants. Biochem. J. 2005, 389, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Zhang, H.; Shi, M.; Yu, Y.; Wang, H.; Cao, W.M.; Zhao, Y.; Zhang, H. TAK1 inhibitor NG25 enhances doxorubicin-mediated apoptosis in breast cancer cells. Sci. Rep. 2016, 6, 32737. [Google Scholar] [CrossRef] [Green Version]
- King, C.G.; Kobayashi, T.; Cejas, P.J.; Kim, T.; Yoon, K.; Kim, G.K.; Chiffoleau, E.; Hickman, S.P.; Walsh, P.T.; Turka, L.A.; et al. TRAF6 is a T cell-intrinsic negative regulator required for the maintenance of immune homeostasis. Nat. Med. 2006, 12, 1088–1092. [Google Scholar] [CrossRef]
- Pearson, L.J.; Williamson, J.H.; Turner, S.A.; Lacy-Hulbert, S.J.; Hillerton, J.E. Peripartum infection with Streptococcus uberis but not coagulase-negative Staphylococci reduced milk production in primiparous cows. J. Dairy Sci. 2013, 96, 158–164. [Google Scholar] [CrossRef] [Green Version]
- Trinidad, P.; Nickerson, S.C.; Adkinson, R.W. Histopathology of staphylococcal mastitis in unbred dairy heifers. J. Dairy Sci. 1990, 73, 639–647. [Google Scholar] [CrossRef]
- Li, M.; Xi, P.; Xu, Y.; Wang, Z.; Han, X.; Ren, W.; Phouthapane, V.; Miao, J. Taurine Attenuates Streptococcus uberis-Induced Bovine Mammary Epithelial Cells Inflammation via Phosphoinositides/Ca2+ Signaling. Front. Immunol. 2019, 10, 1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Z.; Wang, X.; Liu, M.; Zuo, J.; Xu, Y.; Han, X.; Vanhnaseng, P.; Miao, J. Role of Toll-like receptor 2 against Streptococcus uberis infection in primary mouse mammary epithelial cells. Int. Immunopharmacol. 2020, 79, 106142. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Fingar, D.C. Growing knowledge of the mTOR signaling network. Semin. Cell Dev. Biol. 2014, 36, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, M.; Zhang, N.; Li, D.; Liang, D.; Liu, Z.; Li, F.; Fu, Y.; Cao, Y.; Deng, X.; Yang, Z. Baicalin plays an anti-inflammatory role through reducing nuclear factor-κB and p38 phosphorylation in S. aureus-induced mastitis. Int. Immunopharmacol. 2013, 16, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Bhatelia, K.; Singh, K.; Singh, R. TLRs: Linking inflammation and breast cancer. Cell Signal. 2014, 26, 2350–2357. [Google Scholar] [CrossRef]
- Fitzgerald, K.A.; Palsson-McDermott, E.M.; Bowie, A.G.; Jefferies, C.A.; Mansell, A.S.; Brady, G.; Brint, E.; Dunne, A.; Gray, P.; Harte, M.T.; et al. Mal (MyD88-adapter-like) is required for Toll-like receptor-4 signal transduction. Nature 2001, 413, 78–83. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [Green Version]
- Aoyama, T.; Paik, Y.H.; Watanabe, S.; Laleu, B.; Gaggini, F.; Fioraso-Cartier, L.; Molango, S.; Heitz, F.; Merlot, C.; Szyndralewiez, C.; et al. Nicotinamide adenine dinucleotide phosphate oxidase in experimental liver fibrosis: GKT137831 as a novel potential therapeutic agent. Hepatology 2012, 56, 2316–2327. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Tang, Y.; Teng, L.; Wu, Y.; Zhao, X.; Pei, G. Association of beta-arrestin and TRAF6 negatively regulates Toll-like receptor-interleukin 1 receptor signaling. Nat. Immunol. 2006, 7, 139–147. [Google Scholar] [CrossRef]
- Wiersinga, W.J.; van der Poll, T.; White, N.J.; Day, N.P.; Peacock, S.J. Melioidosis: Insights into the pathogenicity of Burkholderia pseudomallei. Nat. Rev. Microbiol. 2006, 4, 272–282. [Google Scholar] [CrossRef]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, T.; Hatton, R.D.; Oliver, J.; Liu, X.; Elson, C.O.; Weaver, C.T. Regulatory T cell suppression and anergy are differentially regulated by proinflammatory cytokines produced by TLR-activated dendritic cells. J. Immunol. 2004, 173, 7249–7258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landskron, G.; De la Fuente, M.; Thuwajit, P.; Thuwajit, C.; Hermoso, M.A. Chronic inflammation and cytokines in the tumor microenvironment. J. Immunol. Res. 2014, 2014, 149185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Liu, Y.L.; Ye, F.; Xie, J.W.; Zeng, J.W.; Qin, L.; Xue, J.; Wang, Y.T.; Guo, K.M.; Ma, M.M.; et al. Free fatty acid-induced H2O2 activates TRPM2 to aggravate endothelial insulin resistance via Ca2+-dependent PERK/ATF4/TRB3 cascade in obese mice. Free Radic. Biol. Med. 2019, 143, 288–299. [Google Scholar] [CrossRef] [PubMed]
- West, A.P.; Shadel, G.S.; Ghosh, S. Mitochondria in innate immune responses. Nat. Rev. Immunol. 2011, 11, 389–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medzhitov, R. Recognition of microorganisms and activation of the immune response. Nature 2007, 449, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, F.R.G.; Lepelley, A.; Seeley, J.J.; Hayden, M.S.; Ghosh, S. An Essential Role for ECSIT in Mitochondrial Complex I Assembly and Mitophagy in Macrophages. Cell Rep. 2018, 22, 2654–2666. [Google Scholar] [CrossRef] [Green Version]
- Richardson, E.T.; Shukla, S.; Sweet, D.R.; Wearsch, P.A.; Tsichlis, P.N.; Boom, W.H.; Harding, C.V. Toll-like receptor 2-dependent extracellular signal-regulated kinase signaling in Mycobacterium tuberculosis-infected macrophages drives anti-inflammatory responses and inhibits Th1 polarization of responding T cells. Infect. Immun. 2015, 83, 2242–2254. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, S.J.; Eckmann, L.; Quayle, A.J.; Shen, L.; Zhang, Y.X.; Anderson, D.J.; Fierer, J.; Stephens, R.S.; Kagnoff, M.F. Secretion of proinflammatory cytokines by epithelial cells in response to Chlamydia infection suggests a central role for epithelial cells in chlamydial pathogenesis. J. Clin. Invest. 1997, 99, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Arsenijevic, D.; Onuma, H.; Pecqueur, C.; Raimbault, S.; Manning, B.S.; Miroux, B.; Couplan, E.; Alves-Guerra, M.C.; Goubern, M.; Surwit, R.; et al. Disruption of the uncoupling protein-2 gene in mice reveals a role in immunity and reactive oxygen species production. Nat. Genet. 2000, 26, 435–439. [Google Scholar] [CrossRef]
- Fremond, C.M.; Yeremeev, V.; Nicolle, D.M.; Jacobs, M.; Quesniaux, V.F.; Ryffel, B. Fatal Mycobacterium tuberculosis infection despite adaptive immune response in the absence of MyD88. J Clin. Invest. 2004, 114, 1790–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouillaud, F. UCP2, not a physiologically relevant uncoupler but a glucose sparing switch impacting ROS production and glucose sensing. Biochim. Biophys. Acta 2009, 1787, 377–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juan, M.E.; Wenzel, U.; Daniel, H.; Planas, J.M. Resveratrol induces apoptosis through ROS-dependent mitochondria pathway in HT-29 human colorectal carcinoma cells. J. Agric. Food Chem. 2008, 56, 4813–4818. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Wan, Z.; Wang, Z.; Zuo, J.; Xu, Y.; Han, X.; Phouthapane, V.; Miao, J. TLR2 Signaling Pathway Combats Streptococcus uberis Infection by Inducing Mitochondrial Reactive Oxygen Species Production. Cells 2020, 9, 494. https://doi.org/10.3390/cells9020494
Li B, Wan Z, Wang Z, Zuo J, Xu Y, Han X, Phouthapane V, Miao J. TLR2 Signaling Pathway Combats Streptococcus uberis Infection by Inducing Mitochondrial Reactive Oxygen Species Production. Cells. 2020; 9(2):494. https://doi.org/10.3390/cells9020494
Chicago/Turabian StyleLi, Bin, Zhixin Wan, Zhenglei Wang, Jiakun Zuo, Yuanyuan Xu, Xiangan Han, Vanhnaseng Phouthapane, and Jinfeng Miao. 2020. "TLR2 Signaling Pathway Combats Streptococcus uberis Infection by Inducing Mitochondrial Reactive Oxygen Species Production" Cells 9, no. 2: 494. https://doi.org/10.3390/cells9020494
APA StyleLi, B., Wan, Z., Wang, Z., Zuo, J., Xu, Y., Han, X., Phouthapane, V., & Miao, J. (2020). TLR2 Signaling Pathway Combats Streptococcus uberis Infection by Inducing Mitochondrial Reactive Oxygen Species Production. Cells, 9(2), 494. https://doi.org/10.3390/cells9020494