Transcriptional and Ultrastructural Analyses Suggest Novel Insights into Epithelial Barrier Impairment in Celiac Disease

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Filtering Criteria of Public RNA-Seq Datasets
2.2. RNA-Seq Analysis
2.3. Patients’ Biopsy Samples
2.4. Ultrastructural Analyses
2.5. Statistical Analysis
2.6. Ethical Statement
3. Results
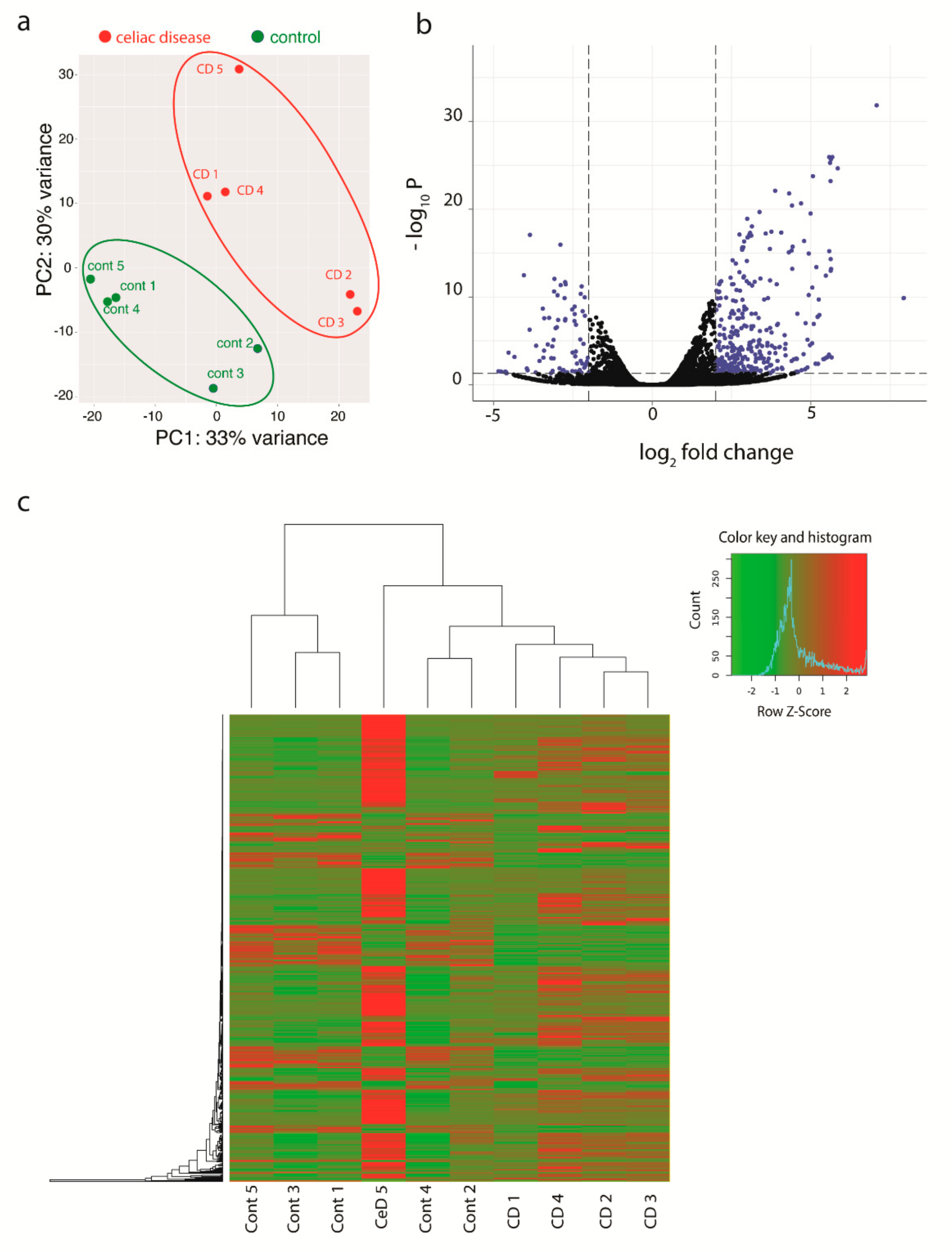
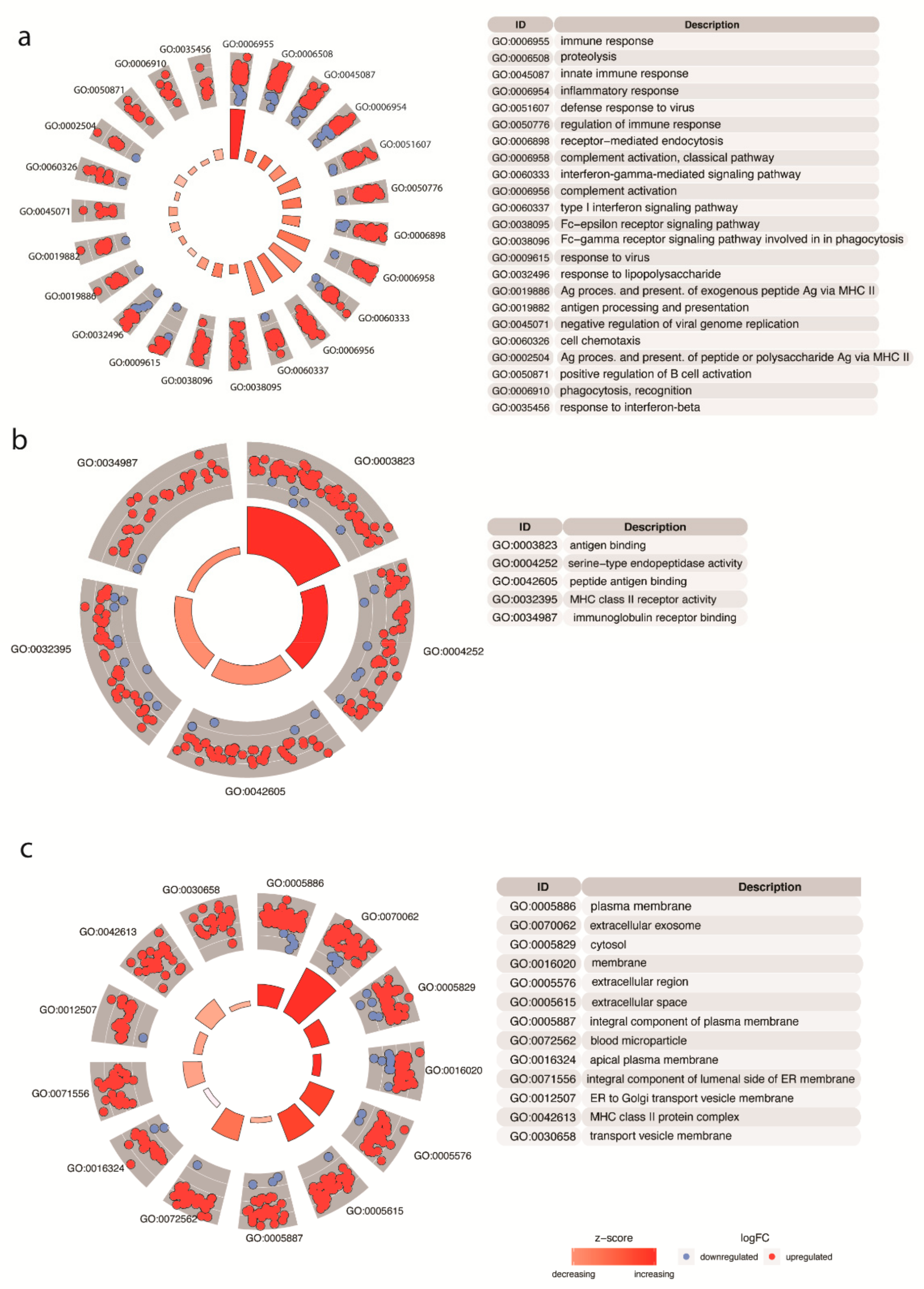
3.1. Transcriptional Analyses
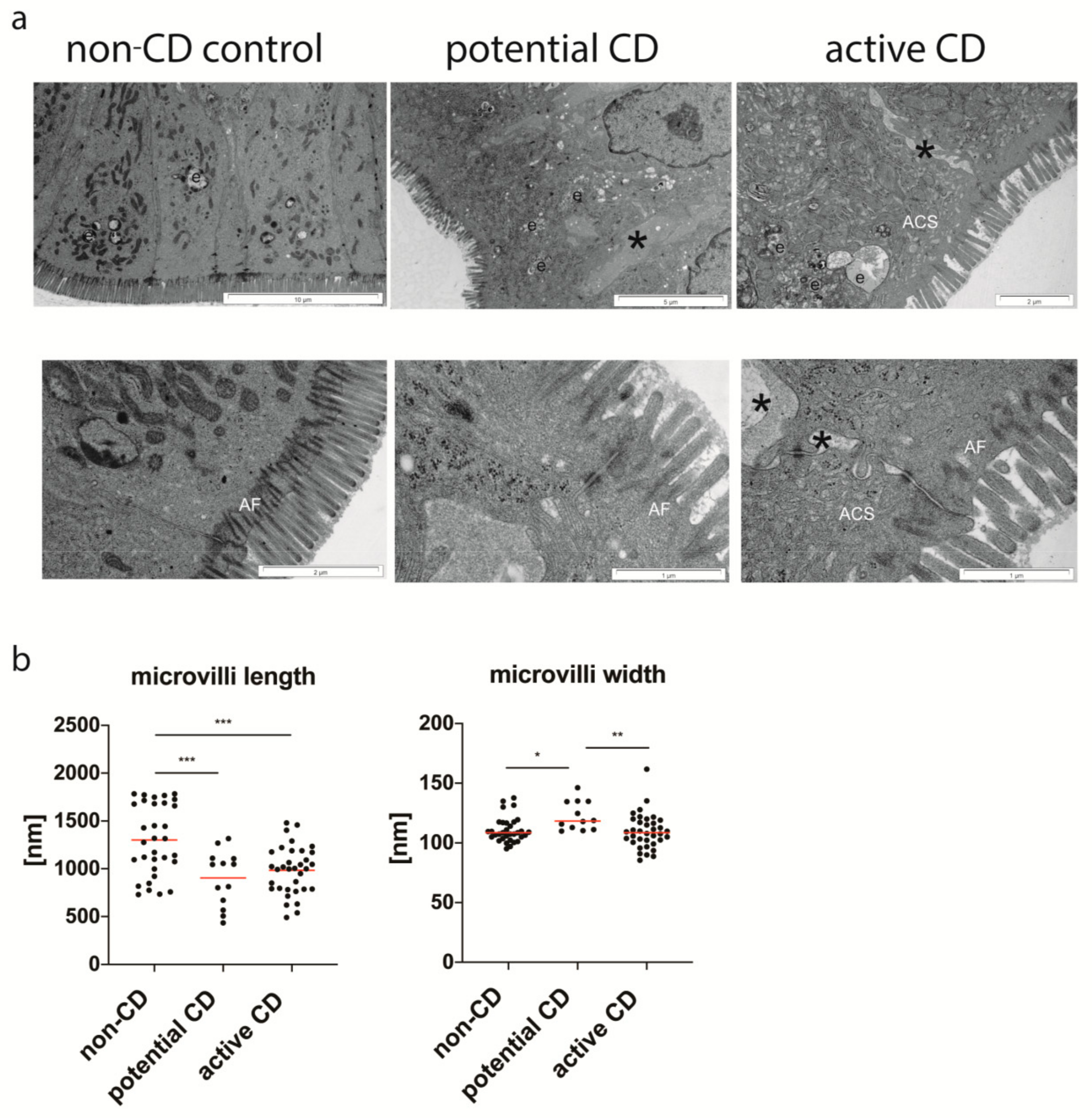
3.2. Ultrastructure of Enterocytes
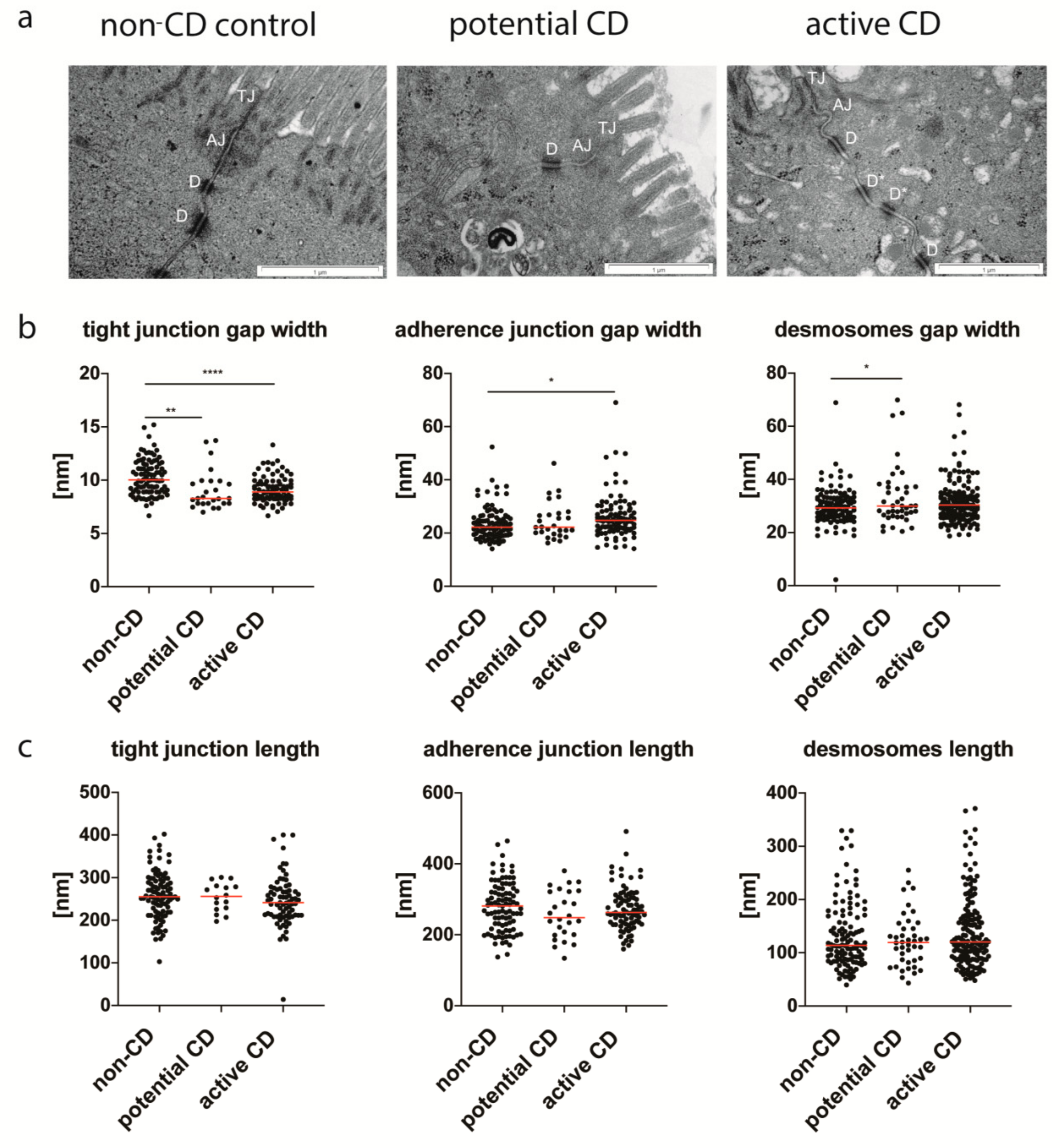
3.3. Enterocyte Intercellular Junctions
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Troncone, R.; Discepolo, V. Celiac disease and autoimmunity. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 9–11. [Google Scholar] [CrossRef]
- Sollid, L.M.; Qiao, S.W.; Anderson, R.P.; Gianfrani, C.; Koning, F. Nomenclature and listing of celiac disease relevant gluten T-cell epitopes restricted by HLA-DQ molecules. Immunogenetics 2012, 64, 455–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Bergen, J.; Mulder, C.J.; Mearin, M.L.; Koning, F. Local communication among mucosal immune cells in patients with celiac disease. Gastroenterology 2015, 148, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Cukrowska, B.; Sowińska, A.; Bierła, J.B.; Czarnowska, E.; Rybak, A.; Grzybowska-Chlebowczyk, U. Intestinal epithelium, intraepithelial lymphocytes and the gut microbiota–Key players in the pathogenesis of celiac disease. World J. Gastroenterol. 2017, 23, 7505–7518. [Google Scholar] [CrossRef] [PubMed]
- Valitutti, F.; Fasano, A. Breaking down barriers: How understanding celiac disease pathogenesis informed the development of novel treatments. Dig. Dis. Sci. 2019, 64, 1748–1758. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A.; Not, T.; Wang, W.; Uzzau, S.; Berti, I.; Tommasini, A.; Goldblum, S.E. Zonulin, a newly discovered modulator of intestinal permeability, and its expression in coeliac disease. Lancet 2000, 355, 1518–1519. [Google Scholar] [CrossRef]
- El Asmar, R.; Panigrahi, P.; Bamford, P.; Berti, I.; Not, T.; Coppa, G.V.; Catassi, C.; Fasano, A. Host-dependent zonulin secretion causes the impairment of the small intestine barrier function after bacterial exposure. Gastroenterology 2001, 123, 1607–1615. [Google Scholar] [CrossRef]
- Lammers, K.M.; Lu, R.; Brownley, J.; Lu, B.; Gerard, C.; Thomas, K.; Rallabhandi, P.; Shea-Donohue, T.; Tamiz, A.; Alkan, S.; et al. Gliadin induces an increase in intestinal permeability and zonulin release by binding to the chemokine receptor CXCR3. Gastroenterology 2008, 135, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Clemente, M.G.; De Virgiliis, S.; Kang, J.S.; Macatagney, R.; Musu, M.P.; Di Pierro, M.R.; Drago, S.; Congia, M.; Fasano, A. Early effects of gliadin on enterocyte intracellular signalling involved in intestinal barrier function. Gut 2003, 52, 218–223. [Google Scholar] [CrossRef]
- Szakál, D.N.; Gyorffy, H.; Arató, A.; Cseh, A.; Molnár, K.; Papp, M.; Dezsofi, A.; Veres, G. Mucosal expression of claudins 2, 3 and 4 in proximal and distal part of duodenum in children with celiac disease. Virchows Arch. 2010, 456, 245–250. [Google Scholar] [CrossRef]
- Ciccocioppo, R.; Finamore, A.; Ara, C.; Di Sabatino, A.; Mengheri, E.; Corazza, G.R. Altered expression, localization, and phosphorylation of epithelial junctional proteins in celiac disease. Am. J. Clin. Pathol. 2006, 125, 502–511. [Google Scholar] [CrossRef]
- Goswami, P.; Das, P.; Verma, A.K.; Prakash, S.; Das, T.K.; Nag, T.C.; Ahuja, V.; Gupta, S.D.; Makharia, G.K. Are alternations of tight junctions at molecular and ultrastructural level different in duodenal biopsies of patients with celiac disease and Crohn’s disease? Virchows Arch. 2014, 465, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Prakash, S.; Sreenivas, V.; Das, T.K.; Ahuja, V.; Gupta, S.D.; Makharia, G.K. Structural and functional changes in the tight junctions of asymptomatic and serology-negative first-degree relatives of patients with celiac disease. J. Clin. Gastroenterol. 2016, 50, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic. Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Oberhuber, G.; Granditsch, G.; Vogelsang, H. The histopathology of celiac disease: Time for a standardized report scheme for pathologists. Eur. J. Gastroenterol. Hepatol. 1999, 11, 1185–1194. [Google Scholar] [CrossRef]
- Husby, S.; Koletzko, S.; Korponay-Szabó, I.R.; Mearin, M.L.; Phillips, A.; Shamir, R.; Troncone, R.; Giersiepen, K.; Branski, D.; Catassi, C.; et al. ESPGHAN Working Group on Coeliac Disease Diagnosis; ESPGHAN Gastroenterology Committee; European Society for Pediatric Gastroenterology, Hepatology, and Nutrition. European Society for Pediatric Gastroenterology, Hepatology, and Nutrition guidelines for the diagnosis of coeliac disease. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 136–160. [Google Scholar] [CrossRef]
- Dyduch, A.; Karczewska, K.; Grzybek, H.; Kamiński, M. Transmission electron microscopy of microvilli of intestinal epithelial cells in celiac disease in remission and transient gluten enteropathy in children after a gluten-free diet. J. Pediatr. Gastroenterol. Nutr. 1993, 16, 269–272. [Google Scholar] [CrossRef]
- Skrzypek, T.; Valverde Piedra, J.L.; Skrzypek, H.; Kazimierczak, W.; Biernat, M.; Zabielski, R. Gradual disappearance of vacuolated enterocytes in the small intestine of neonatal piglets. J. Physiol. Pharmacol. 2007, 58, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Baintner, K. Transmission of antibodies from mother to young: Evolutionary strategies in a proteolytic environment. Vet. Immunol. Immunopathol. 2007, 117, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Berin, M.C.; Kiliaan, A.J.; Yang, P.C.; Groot, J.A.; Kitamura, Y.; Perdue, M.H. The influence of mast cells on pathways of transepithelial antigen transport in rat intestine. J. Immunol. 1998, 161, 2561–2566. [Google Scholar] [PubMed]
- Kersting, S.; Bruewer, M.; Schuermann, G.; Klotz, A.; Utech, M.; Hansmerten, M.; Krieglstein, C.F.; Senninger, N.; Schulzke, J.D.; Naim, H.Y.; et al. Antigen transport and cytoskeletal characteristics of a distinct enterocyte population in inflammatory bowel diseases. Am. J. Pathol. 2004, 165, 425–437. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.C.; Berin, M.C.; Yu, L.C.; Conrad, D.H.; Perdue, M.H. Enhanced intestinal transepithelial antigen transport in allergic rats is mediated by IgE and CD23 (FcepsilonRII). J. Clin. Investig. 2000, 106, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Prakash, S.; Kaur, G.; Sreenivas, V.; Ahuja, V.; Gupta, S.D.; Makharia, G.K. Prevalence of celiac disease among first-degree relatives of Indian celiac disease patients. Dig. Liver. Dis. 2016, 48, 255–259. [Google Scholar] [CrossRef]
- Auricchio, R.; Mandile, R.; Del Vecchio, M.R.; Scapaticci, S.; Galatola, M.; Maglio, M.; Discepolo, V.; Miele, E.; Cielo, D.; Troncone, R.; et al. Progression of celiac disease in children with antibodies against tissue transglutaminase and normal duodenal architecture. Gastroenterology 2019, 157, 413–420.e3. [Google Scholar] [CrossRef]
- Kozakova, H.; Schwarzer, M.; Tuckova, L.; Srutkova, D.; Czarnowska, E.; Rosiak, I.; Hudcovic, T.; Schabussova, I.; Hermanova, P.; Zakostelska, Z.; et al. Colonization of germ-free mice with a mixture of three lactobacillus strains enhances the integrity of gut mucosa and ameliorates allergic sensitization. Cell Mol. Immunol. 2016, 13, 251–262. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Title | Project Accession Number | No of Experiment |
|---|---|---|---|
| 1 | Expression of long non-coding RNAs in autoimmunity and linkage to enhancer function and autoimmune disease risk genetic variants | PRJNA357628 | 15 |
| 2 | Transcriptome of celiac disease | PRJNA327491 | 42 |
| 3 | Chronic inflammation permanently reshapes tissue-resident immunity in celiac disease (human) | PRJNA509448 | 34 |
| 4 | RNA sequencing of intestinal mucosa in celiac patients | PRJNA528755 | 42 |
| 5 | The interplay between IL-15, gluten, and HLA-DQ8 drives the development of coeliac disease in mice (human) | PRJNA556711 | 95 |
| No of Patient | Sex | Age [in Years] | The Level of Anti-tTG2-IgA/[AU/mL] | Histology | Diagnosis |
|---|---|---|---|---|---|
| 1. | Male | 8 | 10 | Marsh 0 | Potential CD |
| 2. | Male | 7 | 19 | Marsh 1 | Potential CD |
| 3. | Female | 6 | 24 | Marsh 1 | Potential CD |
| 4. | Female | 10 | 31 | Marsh 2 | Active CD |
| 5. | Male | 7 | 54 | Marsh 3a | Active CD |
| 6. | Male | 7 | >100 | Marsh 3b | Active CD |
| 7. | Female | 14 | >100 | Marsh 3b | Active CD |
| 8. | Female | 10 | >100 | Marsh 3b | Active CD |
| 9. | Male | 9 | >100 | Marsh 3b | Active CD |
| 10. | Male | 9 | Negative* | Marsh 3b | Active CD |
| 11. | Male | 6 | >100 | Marsh 3b | Active CD |
| 12. | Female | 3 | >100 | Marsh 3c | Active CD |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sowińska, A.; Morsy, Y.; Czarnowska, E.; Oralewska, B.; Konopka, E.; Woynarowski, M.; Szymańska, S.; Ejmont, M.; Scharl, M.; Bierła, J.B.; et al. Transcriptional and Ultrastructural Analyses Suggest Novel Insights into Epithelial Barrier Impairment in Celiac Disease. Cells 2020, 9, 516. https://doi.org/10.3390/cells9020516
Sowińska A, Morsy Y, Czarnowska E, Oralewska B, Konopka E, Woynarowski M, Szymańska S, Ejmont M, Scharl M, Bierła JB, et al. Transcriptional and Ultrastructural Analyses Suggest Novel Insights into Epithelial Barrier Impairment in Celiac Disease. Cells. 2020; 9(2):516. https://doi.org/10.3390/cells9020516
Chicago/Turabian StyleSowińska, Agnieszka, Yasser Morsy, Elżbieta Czarnowska, Beata Oralewska, Ewa Konopka, Marek Woynarowski, Sylwia Szymańska, Maria Ejmont, Michael Scharl, Joanna B. Bierła, and et al. 2020. "Transcriptional and Ultrastructural Analyses Suggest Novel Insights into Epithelial Barrier Impairment in Celiac Disease" Cells 9, no. 2: 516. https://doi.org/10.3390/cells9020516
APA StyleSowińska, A., Morsy, Y., Czarnowska, E., Oralewska, B., Konopka, E., Woynarowski, M., Szymańska, S., Ejmont, M., Scharl, M., Bierła, J. B., Wawrzyniak, M., & Cukrowska, B. (2020). Transcriptional and Ultrastructural Analyses Suggest Novel Insights into Epithelial Barrier Impairment in Celiac Disease. Cells, 9(2), 516. https://doi.org/10.3390/cells9020516