The Winner Takes It All: Auxin—The Main Player during Plant Embryogenesis


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Role of Cytoskeleton in Embryo Development
3. Cytoskeleton Elements are Regulated by Auxin
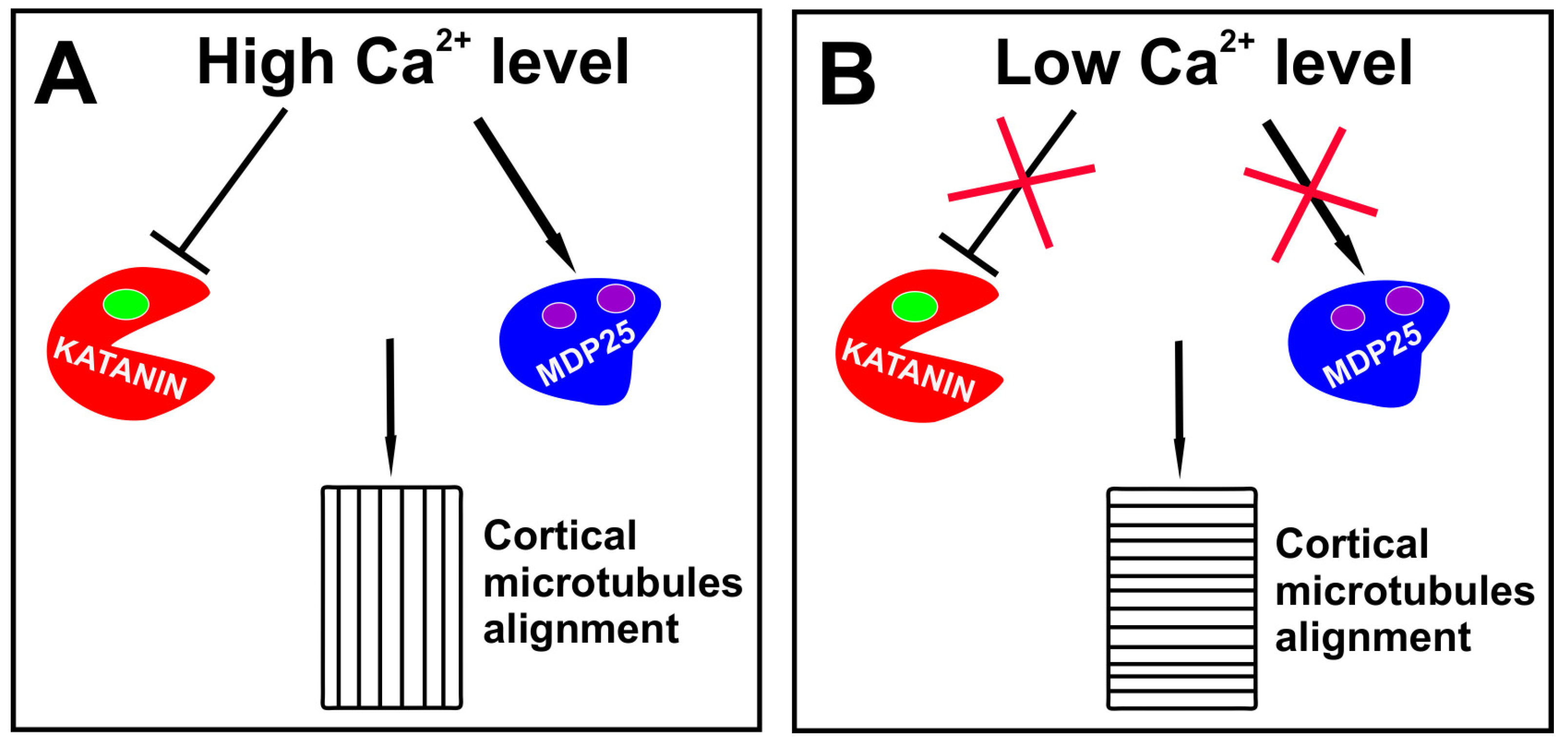
4. The Players Regulating Auxin-Dependent Microtubule Alignment During Embryogenesis
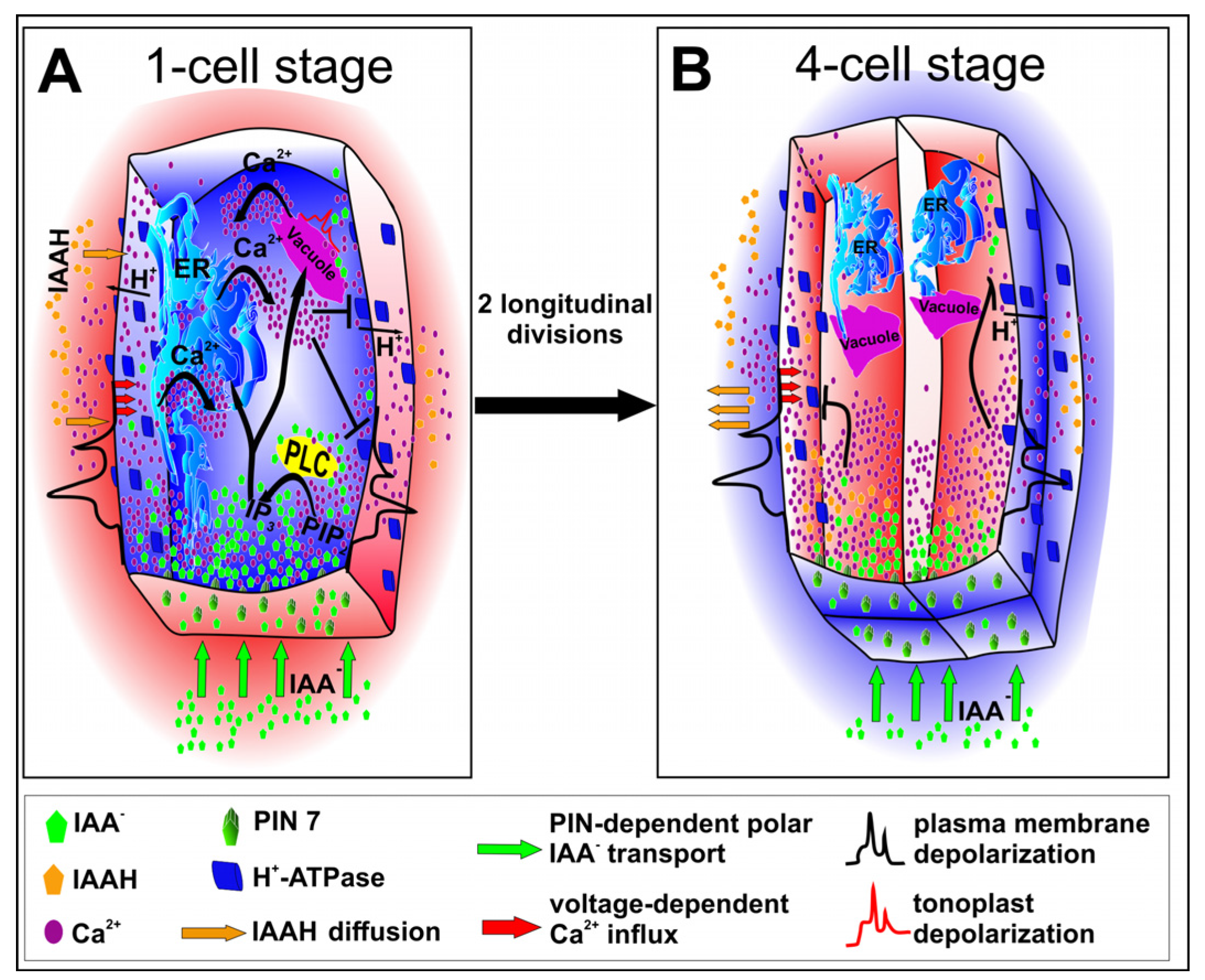
5. Calcium Ions at the Service of Auxin
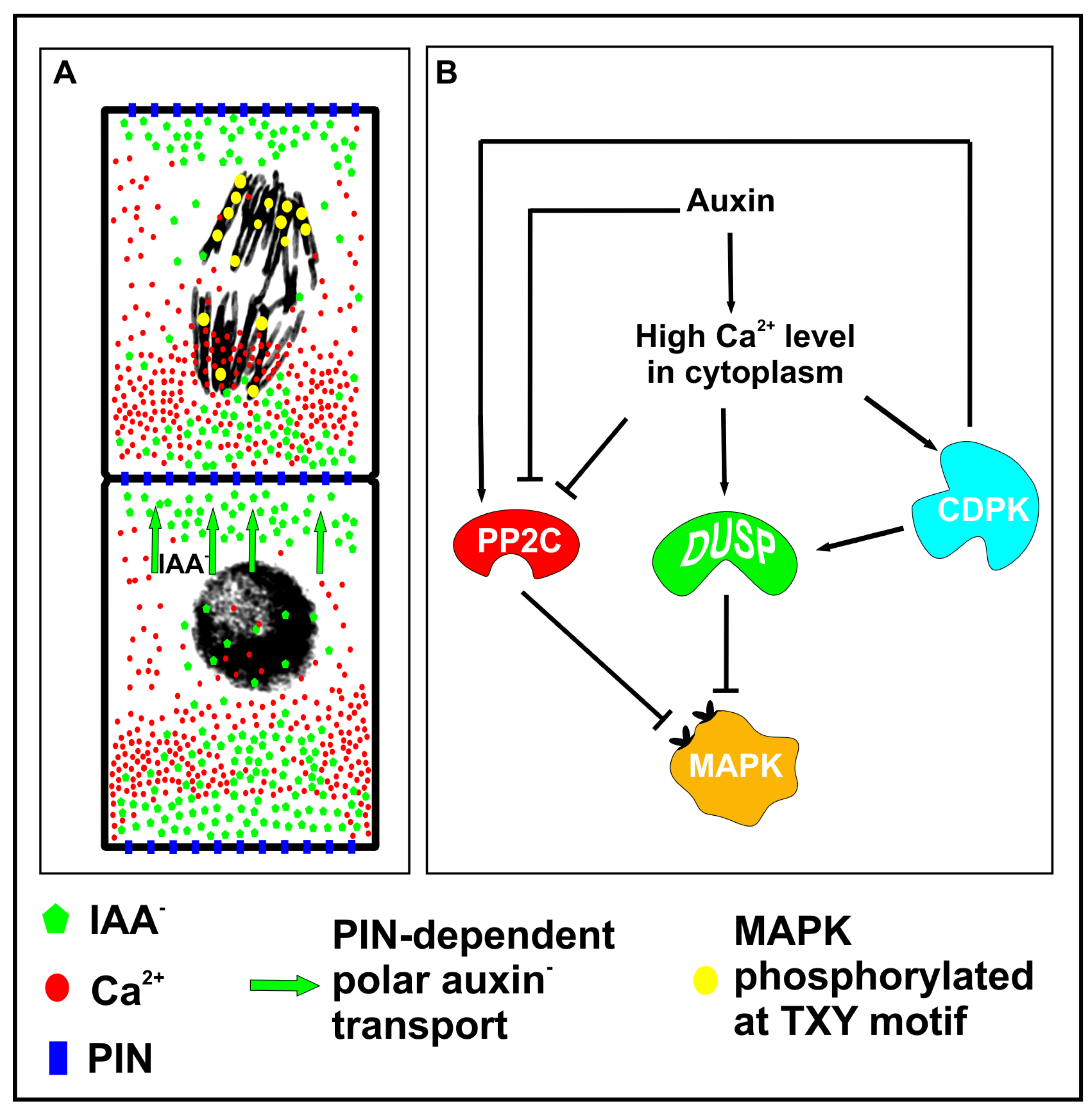
6. Asymmetric Division Depends on Auxin
7. New Mechanism Regulating Intracellular Asymmetry and Cellular Patterning
8. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Robert, H.S.; Grunewald, W.; Sauer, M.; Cannoot, B.; Soriano, M.; Swarup, R.; Weijers, D.; Bennett, M.; Boutilier, K.; Friml, J. Plant embryogenesis requires aux/lax-mediated auxin influx. Development 2015, 142, 702–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaiwongsar, S.; Strohm, A.K.; Su, S.-H.; Krysan, P.J. Genetic analysis of the Arabidopsis protein kinases MAP3Kε1 and MAP3Kε2 indicates roles in cell expansion and embryo development. Front. Plant Sci. 2012, 3, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckardt, N.A. Genetic and epigenetic regulation of embryogenesis. Plant Cell Online 2006, 18, 781–784. [Google Scholar] [CrossRef]
- Bayer, M.; Slane, D.; Jürgens, G. Early plant embryogenesis—Dark ages or dark matter? Curr. Opin. Plant Biol. 2017, 35, 30–36. [Google Scholar] [CrossRef]
- Capron, A.; Chatfield, S.; Provart, N.; Berleth, T. Embryogenesis: Pattern formation from a single cell. In The Arabidopsis Book; American Society of Plant Biologists: Rockville, MD, USA, 2009. [Google Scholar]
- Zhao, P.; Shi, D.Q.; Yang, W.C. Patterning the embryo in higher plants: Emerging pathways and challenges. Front. Biol. China 2011, 6, 3–11. [Google Scholar] [CrossRef]
- Zaban, B.; Liu, W.; Jiang, X.; Nick, P. Plant cells use auxin efflux to explore geometry. Sci. Rep. 2014, 4, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Friml, J.; Vieten, A.; Sauer, M.; Weijers, D.; Schwarz, H.; Hamann, T.; Offringa, R.; Jürgens, G. Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 2003, 426, 147–153. [Google Scholar] [CrossRef]
- Möller, B.; Weijers, D. Auxin control of embryo patterning. Cold Spring Harb. Perspect. Biol. 2009, 1, a001545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S.; Bayer, M.; Lukowitz, W. Taking the very first steps: From polarity to axial domains in the early Arabidopsis embryo. J. Exp. Bot. 2011, 62, 1687–1697. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Dhonukshe, P.; Brewer, P.B.; Friml, J. Spatiotemporal asymmetric auxin distribution: A means to coordinate plant development. Cell. Mol. Life Sci. 2006, 63, 2738–2754. [Google Scholar] [CrossRef]
- Spitzer, C.; Reyes, F.C.; Buono, R.; Sliwinski, M.K.; Haas, T.J.; Otegui, M.S. The ESCRT-Related CHMP1A and B proteins mediate multivesicular body sorting of auxin carriers in Arabidopsis and are required for plant development. Plant Cell 2009, 21, 749–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimata, Y.; Higaki, T.; Kawashima, T.; Kurihara, D.; Sato, Y.; Yamada, T.; Hasezawa, S.; Berger, F.; Higashiyama, T.; Ueda, M. Cytoskeleton dynamics control the first asymmetric cell division in Arabidopsis zygote. Proc. Natl. Acad. Sci. USA 2016, 113, 14157–14162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunda, P.; Baum, B. The actin cytoskeleton in spindle assembly and positioning. Trends Cell Biol. 2009, 19, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Wendrich, J.R.; Weijers, D. The Arabidopsis embryo as a miniature morphogenesis model. New Phytol. 2013, 199, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Laux, T. The asymmetric division of the Arabidopsis zygote: From cell polarity to an embryo axis. Sex. Plant Reprod. 2011, 24, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Begcy, K.; Dresselhaus, T.; Sun, M.-X. Does early embryogenesis in eudicots and monocots involve the same mechanism and molecular players? Plant Physiol. 2017, 173, 130–142. [Google Scholar] [CrossRef] [Green Version]
- Sablowski, R. Coordination of plant cell growth and division: Collective control or mutual agreement? Curr. Opin. Plant Biol. 2016, 34, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Besson, S.; Dumais, J. Universal rule for the symmetric division of plant cells. Proc. Natl. Acad. Sci. USA 2011, 108, 6294–6299. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, B.E.; Tobin, C.; Mjolsness, E.; Meyerowitz, E.M. Analysis of cell division patterns in the Arabidopsis shoot apical meristem. Proc. Natl. Acad. Sci. USA 2015, 112, 4815–4820. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; BarbierdeReuille, P.; Lane, B.; Bassel, G.W.; Prusinkiewicz, P.; Smith, R.S.; Weijers, D. Genetic control of plant development by overriding a geometric division rule. Dev. Cell 2014, 29, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Han, L.; Feng, Z.; Wang, G.; Liu, W.; Ma, Y.; Yu, Y.; Kong, Z. Orchestration of microtubules and the actin cytoskeleton in trichome cell shape determination by a plant-unique kinesin. Elife 2015, 4, e09351. [Google Scholar] [CrossRef] [PubMed]
- Sambade, A.; Findlay, K.; Schäffner, A.R.; Lloyd, C.W.; Buschmann, H. Actin-dependent and -independent functions of cortical microtubules in the differentiation of Arabidopsis leaf trichomes. Plant Cell 2014, 26, 1629–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blancaflor, E.B. Cortical actin filaments potentially interact with cortical microtubules in regulating polarity of cell expansion in primary roots of maize (Zea mays L.). J. Plant Growth Regul. 2000, 19, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Louveaux, M.; Julien, J.D.; Mirabet, V.; Boudaoud, A.; Hamant, O. Cell division plane orientation based on tensile stress in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2016, 113, E4294–E4303. [Google Scholar] [CrossRef] [Green Version]
- Verger, S.; Long, Y.; Boudaoud, A.; Hamant, O. A tension-adhesion feedback loop in plant epidermis. Elife 2018, 7, e34460. [Google Scholar] [CrossRef] [PubMed]
- Sampathkumar, A.; Krupinski, P.; Wightman, R.; Milani, P.; Berquand, A.; Boudaoud, A.; Hamant, O.; Jönsson, H.; Meyerowitz, E.M. Subcellular and supracellular mechanical stress prescribes cytoskeleton behavior in Arabidopsis cotyledon pavement cells. Elife 2014, 3, e01967. [Google Scholar] [CrossRef]
- Boutté, Y.; Crosnier, M.T.; Carraro, N.; Traas, J.; Satiat-Jeunemaitre, B. The plasma membrane recycling pathway and cell polarity in plants: Studies on PIN proteins. J. Cell Sci. 2006, 119, 1255–1265. [Google Scholar] [CrossRef] [Green Version]
- Heisler, M.G.; Hamant, O.; Krupinski, P.; Uyttewaal, M.; Ohno, C.; Jönsson, H.; Traas, J.; Meyerowitz, E.M. Alignment between PIN1 polarity and microtubule orientation in the shoot apical meristem reveals a tight coupling between morphogenesis and auxin transport. PLoS Biol. 2010, 8, e1000516. [Google Scholar] [CrossRef] [Green Version]
- Landrein, B.; Hamant, O. How mechanical stress controls microtubule behavior and morphogenesis in plants: History, experiments and revisited theories. Plant J. 2013, 75, 324–338. [Google Scholar] [CrossRef]
- Asnacios, A.; Hamant, O. The mechanics behind cell polarity. Trends Cell Biol. 2012, 22, 584–591. [Google Scholar] [CrossRef]
- Smith, L.G. Cytoskeletal control of plant cell shape: Getting the fine points. Curr. Opin. Plant Biol. 2003, 6, 63–73. [Google Scholar] [CrossRef]
- Peremyslov, V.V.; Prokhnevsky, A.I.; Dolja, V.V. Class XI myosins are required for development, cell expansion, and F-actin organization in Arabidopsis. Plant Cell 2010, 22, 1883–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovaleva, L.V.; Voronkov, A.S.; Zakharova, E.V. Role of auxin and cytokinin in the regulation of the actin cytoskeleton in the in vitro germinating male gametophyte of petunia. Russ. J. Plant Physiol. 2015, 62, 179–186. [Google Scholar] [CrossRef]
- Lee, Y.J.; Szumlanski, A.; Nielsen, E.; Yang, Z. Rho-GTPase-dependent filamentous actin dynamics coordinate vesicle targeting and exocytosis during tip growth. J. Cell Biol. 2008, 181, 1155–1168. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.; Zhang, R.; Zhang, M.; Diao, M.; Xue, Y.; Huang, S. Organizational innovation of apical actin filaments drives rapid pollen tube growth and turning. Mol. Plant 2017, 10, 930–947. [Google Scholar] [CrossRef] [Green Version]
- Eitzen, G. Actin remodeling to facilitate membrane fusion. Biochim. Biophys. Acta Mol. Cell Res. 2003, 1641, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Bademosi, A.T.; Luo, J.; Meunier, F.A. Actin remodeling in regulated exocytosis: Toward a mesoscopic view. Trends Cell Biol. 2018, 28, 685–697. [Google Scholar] [CrossRef] [Green Version]
- Heinemann, F.; Vogel, S.K.; Schwille, P. Lateral membrane diffusion modulated by a minimal actin cortex. Biophys. J. 2013, 104, 1465–1475. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Shi, H.; Chen, B.; Zhang, R.; Huang, S.; Fu, Y. Arabidopsis RIC1 severs actin filaments at the apex to regulate pollen tube growth. Plant Cell 2015, 27, 1140–1161. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, M.; Staehelin, L.A.; Mineyuki, Y. Actin-Microtubule Interaction in Plants, Cytoskeleton–Structure, Dynamics, Function and Disease; Jimenez-Lopez, J.C., Ed.; IntechOpen: Rijeka, Croatia, 2017. [Google Scholar]
- Sampathkumar, A.; Lindeboom, J.J.; Debolt, S.; Gutierrez, R.; Ehrhardt, D.W.; Ketelaar, T.; Persson, S. Live cell imaging reveals structural associations between the actin and microtubule cytoskeleton in arabidopsis. Plant Cell 2011, 23, 2302–2313. [Google Scholar] [CrossRef] [Green Version]
- Chakrabortty, B.; Blilou, I.; Scheres, B.; Mulder, B.M. A computational framework for cortical microtubule dynamics in realistically shaped plant cells. PLoS Comput. Biol. 2018, 14, e1005959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabortty, B.; Willemsen, V.; de Zeeuw, T.; Liao, C.Y.; Weijers, D.; Mulder, B.; Scheres, B. A plausible microtubule-based mechanism for cell division orientation in plant embryogenesis. Curr. Biol. 2018, 28, 3031–3043.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brun, L.; Rupp, B.; Ward, J.J.; Nédélec, F. A theory of microtubule catastrophes and their regulation. Proc. Natl. Acad. Sci. USA 2009, 106, 21173–21178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, A.; Shaw, S.L. Update: Plant cortical microtubule arrays. Plant Physiol. 2018, 176, 94–105. [Google Scholar] [CrossRef] [Green Version]
- Vineyard, L.; Elliott, A.; Dhingra, S.; Lucas, J.R.; Shaw, S.L. Progressive transverse microtubule array organization in hormone-induced Arabidopsis hypocotyl cells. Plant Cell 2013, 25, 662–676. [Google Scholar] [CrossRef] [Green Version]
- Ivanchenko, M.G.; Napsucialy-Mendivil, S.; Dubrovsky, J.G. Auxin-induced inhibition of lateral root initiation contributes to root system shaping in Arabidopsis thaliana. Plant J. 2010, 64, 740–752. [Google Scholar] [CrossRef]
- Chen, X.; Grandont, L.; Li, H.; Hauschild, R.; Paque, S.; Abuzeineh, A.; Rakusová, H.; Benkova, E.; Perrot-Rechenmann, C.; Friml, J. Inhibition of cell expansion by rapid ABP1-mediated auxin effect on microtubules. Nature 2014, 516, 90–93. [Google Scholar] [CrossRef]
- Oda, Y. Cortical microtubule rearrangements and cell wall patterning. Front. Plant Sci. 2015, 6, 236. [Google Scholar] [CrossRef] [Green Version]
- Hamann, T.; Mayer, U.; Jürgens, G. The auxin-insensitive bodenlos mutation affects primary root formation and apical-basal patterning in the Arabidopsis embryo. Development 1999, 126, 1387–1395. [Google Scholar]
- Hamann, T.; Benkova, E.; Bäurle, I.; Kientz, M.; Jürgens, G. The Arabidopsis BODENLOS gene encodes an auxin response protein inhibiting MONOPTEROS-mediated embryo patterning. Genes Dev. 2002, 16, 1610–1615. [Google Scholar] [CrossRef] [Green Version]
- Sassi, M.; Traas, J. New insights in shoot apical meristem morphogenesis: Isotropy comes into play. Plant Signal. Behav. 2015, 10, e1000150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sassi, M.; Ali, O.; Boudon, F.; Cloarec, G.; Abad, U.; Cellier, C.; Chen, X.; Gilles, B.; Milani, P.; Friml, J.; et al. An auxin-mediated shift toward growth isotropy promotes organ formation at the shoot meristem in arabidopsis. Curr. Biol. 2014, 24, 2335–2342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wabnik, K.; Robert, H.S.; Smith, R.S.; Friml, J. Modeling framework for the establishment of the apical-basal embryonic axis in plants. Curr. Biol. 2013, 23, 2513–2518. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.Y.; Shi, D.Q.; Jia, P.F.; Tang, J.; Li, H.J.; Liu, J.; Yang, W.C. The arabidopsis receptor kinase ZAR1 is required for zygote asymmetric division and its daughter cell fate. PLoS Genet. 2016, 12, e1005933. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Guo, Z.; Song, L.; Wang, Y.; Cheng, Y. NCP1/AtMOB1A plays key roles in auxin-mediated arabidopsis development. PLoS Genet. 2016, 12, e1005923. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Shi, C.; Zhao, P.; Sun, M. Isolation of living apical and basal cell lineages of early proembryos for transcriptome analysis. Plant Reprod. 2019, 32, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Babu, Y.; Musielak, T.; Henschen, A.; Bayer, M. Suspensor length determines developmental progression of the embryo in Arabidopsis. Plant Physiol. 2013, 162, 1448–1458. [Google Scholar] [CrossRef] [Green Version]
- Jenik, P.D.; Barton, M.K. Surge and destroy: The role of auxin in plant embryogenesis. Development 2005, 132, 3577–3585. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, X.; Zhao, J.; Tang, X.; Tian, S.; Chen, J.; Shi, C.; Wang, W.; Zhang, L.; Feng, X.; et al. Direct evidence that suspensor cells have embryogenic potential that is suppressed by the embryo proper during normal embryogenesis. Proc. Natl. Acad. Sci. USA 2015, 112, 12432–12437. [Google Scholar] [CrossRef] [Green Version]
- Gooh, K.; Ueda, M.; Aruga, K.; Park, J.; Arata, H.; Higashiyama, T.; Kurihara, D. Live-cell imaging and optical manipulation of Arabidopsis early embryogenesis. Dev. Cell 2015, 34, 242–251. [Google Scholar] [CrossRef] [Green Version]
- Nick, P.; Han, M.J.; An, G. Auxin Stimulates its own transport by shaping actin filaments. Plant Physiol. 2009, 151, 155–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nick, P. Probing the actin-auxin oscillator. Plant Signal. Behav. 2010, 5, 94–98. [Google Scholar] [CrossRef]
- Waller, F.; Riemann, M.; Nick, P. A role for actin-driven secretion in auxin-induced growth. Protoplasma 2002, 219, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Geisler, M. Keeping it all together: Auxin-actin crosstalk in plant development. J. Exp. Bot. 2015, 66, 4983–4998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, H.W.; Chen, X.; Mei, Y. Function and regulation of phospholipid signalling in plants. Biochem. J. 2009, 421, 145–156. [Google Scholar] [CrossRef]
- Qu, L.H.; Zhou, X.; Li, X.; Li, S.S.; Zhao, J.; Zhao, P.; Liu, Y.; Sun, M.X. The autonomous cell fate specification of basal cell lineage: The initial round of cell fate specification occurs at the two-celled proembryo stage. Plant J. 2017, 91, 1051–1063. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Guo, Y.; Zhao, P.; Sun, M.X. Comparative analysis of WUSCHEL-related homeobox genes revealed their parent-of-origin and cell type-specific expression pattern during early embryogenesis in tobacco. Front. Plant Sci. 2018, 9, 311. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Sun, M.X. NtDRP is necessary for accurate zygotic division orientation and differentiation of basal cell lineage toward suspensor formation. New Phytol. 2016, 212, 598–612. [Google Scholar] [CrossRef] [Green Version]
- Lindeboom, J.J.; Nakamura, M.; Saltini, M.; Hibbel, A.; Walia, A.; Ketelaar, T.; Emons, A.M.C.; Sedbrook, J.C.; Kirik, V.; Mulder, B.M.; et al. CLASP stabilization of plus ends created by severing promotes microtubule creation and reorientation. J. Cell Biol. 2019, 218, 190–205. [Google Scholar] [CrossRef]
- Ambrose, J.C.; Shoji, T.; Kotzer, A.M.; Pighin, J.A.; Wasteneys, G.O. The Arabidopsis CLASP gene encodes a microtubule-associated protein involved in cell expansion and division. Plant Cell 2007, 19, 2763–2775. [Google Scholar] [CrossRef] [Green Version]
- Luptovčiak, I.; Komis, G.; Takáč, T.; Ovečka, M.; Šamaj, J. Katanin: A sword cutting microtubules for cellular, developmental, and physiological purposes. Front. Plant Sci. 2017, 8, 1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burk, D.H.; Ye, Z.H. Alteration of oriented deposition of cellulose microfibrils by mutation of a katanin-like microtubule-severing protein. Plant Cell 2002, 14, 2145–2160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luptovciak, I.; Samakovli, D.; Komis, G.; Šamaj, J. KATANIN 1 is essential for embryogenesis and seed formation in Arabidopsis. Front. Plant Sci. 2017, 8, 728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Y.; Dong, J. Focusing on the focus: What else beyond the master switches for polar cell growth? Mol. Plant 2015, 8, 582–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takáč, T.; Šamajová, O.; Pechan, T.; Luptovčiak, I.; Šamaj, J. Feedback microtubule control and microtubule-actin cross-talk in arabidopsis revealed by integrative proteomic and cell biology analysis of KATANIN 1 mutants. Mol. Cell. Proteomics 2017, 16, 1591–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanneste, S.; Friml, J. Calcium: The missing link in auxin action. Plants 2013, 2, 650–675. [Google Scholar] [CrossRef] [Green Version]
- Iwaya, N.; Akiyama, K.; Goda, N.; Tenno, T.; Fujiwara, Y.; Hamada, D.; Ikura, T.; Shirakawa, M.; Hiroaki, H. Effect of Ca 2+ on the microtubule-severing enzyme p60-katanin. Insight into the substrate-dependent activation mechanism. FEBS J. 2012, 279, 1339–1352. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Qin, T.; Zhang, Y.; Liu, X.; Sun, J.; Zhou, Y.; Zhu, L.; Zhang, Z.; Yuan, M.; et al. MDP25, a novel calcium regulatory protein, mediates hypocotyl cell elongation by destabilizing cortical microtubules in Arabidopsis. Plant Cell 2011, 23, 4411–4427. [Google Scholar] [CrossRef] [Green Version]
- Qin, T.; Li, J.; Yuan, M.; Mao, T. Characterization of the role of calcium in regulating the microtubule-destabilizing activity of MDP25. Plant Signal. Behav. 2012, 7, 708–710. [Google Scholar] [CrossRef] [Green Version]
- Scherer, G.F.E. AUXIN-BINDING-PROTEIN1, the second auxin receptor: What is the significance of a two-receptor concept in plant signal transduction? J. Exp. Bot. 2011, 62, 3339–3357. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Zhang, W. Regulation of developmental and environmental signaling by interaction between microtubules and membranes in plant cells. Protein Cell 2016, 7, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamant, O.; Heisler, M.G.; Jönsson, H.; Krupinski, P.; Uyttewaal, M.; Bokov, P.; Corson, F.; Sahlin, P.; Boudaoud, A.; Meyerowitz, E.M.; et al. Developmental patterning by mechanical signals in Arabidopsis. Science 2008, 322, 1650–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swarbreck, S.M.; Colaço, R.; Davies, J.M. Plant calcium-permeable channel. Plant Physiol. 2013, 163, 514–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falhof, J.; Pedersen, J.T.; Fuglsang, A.T.; Palmgren, M. Plasma membrane H+-ATPase regulation in the center of plant physiology. Mol. Plant 2016, 9, 323–337. [Google Scholar] [CrossRef] [Green Version]
- Dindas, J.; Scherzer, S.; Roelfsema, M.R.G.; Von Meyer, K.; Müller, H.M.; Al-Rasheid, K.A.S.; Palme, K.; Dietrich, P.; Becker, D.; Bennett, M.J.; et al. AUX1-mediated root hair auxin influx governs SCFTIR1/AFB-type Ca2+ signaling. Nat. Commun. 2018, 9, 1174. [Google Scholar] [CrossRef]
- Ephritikhine, G.; Barbier-Brygoo, H.; Muller, J.-F.; Guern, J. Auxin effect on the transmembrane potential difference of wild-type and mutant tobacco protoplasts exhibiting a differential sensitiity to auxin. Plant Physiol. 1987, 83, 801–804. [Google Scholar] [CrossRef]
- Felle, H.; Peters, W.; Palme, K. The electrical response of maize to auxins. BBA Biomembr. 1991, 1064, 199–204. [Google Scholar] [CrossRef]
- Krebs, M.; Waadt, R.; Wang, Y.; Dindas, J.; Rienmu, F.; Schumacher, K.; Wu, W.; Hedrich, R.; Roelfsema, M.R.G. Cytosolic Ca 2+ signals enhance the vacuolar ion conductivity of bulging Arabidopsis root hair cells. Mol. Plant 2015, 8, 1665–1674. [Google Scholar]
- Schönknecht, G. Calcium signals from the vacuole. Plants 2013, 2, 589–614. [Google Scholar] [CrossRef] [Green Version]
- Kopka, J.; Pical, C.; Gray, J.E.; Müller-Röber, B. Molecular and enzymatic characterization of three phosphoinositide-specific phospholipase C isoforms from potato. Plant Physiol. 1998, 116, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Gonzales, R.A.; Bhattacharyya, M.K. Characterization of a plasma membrane-associated phosphoinositide-specific phospholipase C from soybean. Plant J. 1995, 8, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Stael, S.; Wurzinger, B.; Mair, A.; Mehlmer, N.; Vothknecht, U.C.; Teige, M. Plant organellar calcium signalling: An emerging field. J. Exp. Bot. 2012, 63, 1525–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krinke, O.; Novotná, Z.; Valentová, O.; Martinec, J. Inositol trisphosphate receptor in higher plants: Is it real? J. Exp. Bot. 2007, 58, 361–376. [Google Scholar] [CrossRef] [Green Version]
- Kimata, Y.; Kato, T.; Higaki, T.; Kurihara, D.; Yamada, T.; Segami, S.; Morita, M.T.; Maeshima, M.; Hasezawa, S.; Higashiyama, T.; et al. Polar vacuolar distribution is essential for accurate asymmetric division of Arabidopsis zygotes. Proc. Natl. Acad. Sci. USA 2019, 116, 2338–2343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, C.Y.; Weijers, D. A toolkit for studying cellular reorganization during early embryogenesis in Arabidopsis thaliana. Plant J. 2018, 93, 963–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pottosin, I.I.; Schönknecht, G. Vacuolar calcium channels. J. Exp. Bot. 2007, 58, 1559–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bethke, P.C.; Jones, R.L. Ca2+-calmodulin modulates lon channel activity in storage protein vacuoles of barley aleurone cells. Plant Cell 1994, 6, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Anil, V.S.; Rao, K.S. Calcium-mediated signaling during sandalwood somatic embryogenesis. Role for exogenous calcium as second messenger. Plant Physiol. 2000, 123, 1301–1311. [Google Scholar] [CrossRef] [Green Version]
- Mahalakshmi, A.; Singla, B.; Khurana, J.P.; Khurana, P. Role of calcium-calmodulin in auxin-induced somatic embryogenesis in leaf base cultures of wheat (Triticum aestivum var. HD 2329). Plant Cell Tissue Organ Cult. 2007, 88, 167–174. [Google Scholar] [CrossRef]
- Zeng, F.; Zhang, X.; Cheng, L.; Hu, L.; Zhu, L.; Cao, J.; Guo, X. A draft gene regulatory network for cellular totipotency reprogramming during plant somatic embryogenesis. Genomics 2007, 90, 620–628. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, T.; Nishimura, M.; Shimazaki, K.I. Cytosolic concentration of Ca2+ regulates the plasma membrane H+-ATPase in guard cells of fava bean. Plant Cell 1995, 7, 1333–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Nisi, P.; Dell’Orto, M.; Pirovano, L.; Zocchi, G. Calcium-dependent phosphorylation regulates the plasma-membrane H+-ATPase activity of maize (Zea mays L.) roots. Planta 1999, 209, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Brault, M.; Amiar, Z.; Pennarun, A.-M.; Monestiez, M.; Zhang, Z.; Cornel, D.; Dellis, O.; Knight, H.; Bouteau, F.; Rona, J.-P. Plasma membrane depolarization induced by abscisic acid in Arabidopsis suspension cells involves reduction of proton pumping in addition to anion channel activation, which are both Ca21 dependent. Plant Physiol. 2004, 135, 231–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz-Lessdorf, B.; Hedrich, R. Protons and calcium modulate SV-type channels in the vacuolar-lysosomal compartment-channel interaction with calmodulin inhibitors. Planta 1995, 197, 655–671. [Google Scholar] [CrossRef]
- Balzan, S.; Johal, G.S.; Carraro, N. The role of auxin transporters in monocots development. Front. Plant Sci. 2014, 5, 393. [Google Scholar] [CrossRef] [Green Version]
- Lucas, J.R.; Nadeau, J.A.; Sack, F.D. Microtubule arrays and Arabidopsis stomatal development. J. Exp. Bot. 2006, 57, 71–79. [Google Scholar] [CrossRef]
- Le, J.; Liu, X.G.; Yang, K.Z.; Chen, X.L.; Zou, J.J.; Wang, H.Z.; Wang, M.; Vanneste, S.; Morita, M.; Tasaka, M.; et al. Auxin transport and activity regulate stomatal patterning and development. Nat. Commun. 2014, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, E.; Belcram, K.; Uyttewaal, M.; Duroc, Y.; Goussot, M.; Legland, D.; Laruelle, E.; De Tauzia-Moreau, M.L.; Pastuglia, M.; Bouchez, D. The preprophase band of microtubules controls the robustness of division orientation in plants. Science 2017, 356, 186–189. [Google Scholar] [CrossRef]
- Livanos, P.; Giannoutsou, E.; Apostolakos, P.; Galatis, B. Auxin as an inducer of asymmetrical division generating the subsidiary cells in stomatal complexes of Zea mays. Plant Signal. Behav. 2015, 10, e984531. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Tucker, E.; Hermann, M.; Laux, T. A molecular framework for the embryonic initiation of shoot meristem stem cells. Dev. Cell 2017, 40, 264–277.e4. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.J.; Park, J.W.; Lee, H.W.; Kim, J. Genome-wide analysis of the auxin-responsive transcriptome downstream of iaa1 and its expression analysis reveal the diversity and complexity of auxin-regulated gene expression. J. Exp. Bot. 2009, 60, 3935–3957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paponov, I.A.; Paponov, M.; Teale, W.; Menges, M.; Chakrabortee, S. Comprehensive transcriptome analysis of auxin responses in Arabidopsis. Mol. Plant 2008, 1, 321–337. [Google Scholar] [CrossRef]
- Habets, M.E.J.; Offringa, R. PIN-driven polar auxin transport in plant developmental plasticity: A key target for environmental and endogenous signals. New Phytol. 2014, 203, 362–377. [Google Scholar] [CrossRef]
- He, P.; Zhao, P.; Wang, L.; Zhang, Y.; Wang, X.; Xiao, H.; Yu, J.; Xiao, G. The PIN gene family in cotton (Gossypium hirsutum): Genome-wide identification and gene expression analyses during root development and abiotic stress responses. BMC Genom. 2017, 18, 507. [Google Scholar] [CrossRef] [Green Version]
- Smith, Z.R.; Long, J.A. Control of Arabidopsis apical-basal embryo polarity by antagonistic transcription factors. Nature 2010, 464, 423–426. [Google Scholar] [CrossRef] [Green Version]
- Rosspopoff, O.; Chelysheva, L.; Saffar, J.; Lecorgne, L.; Gey, D.; Caillieux, E.; Colot, V.; Roudier, F.; Hilson, P.; Berthomé, R.; et al. Direct conversion of root primordium into shoot meristem relies on timing of stem cell niche development. Development 2017, 144, 1187–1200. [Google Scholar] [CrossRef] [Green Version]
- Peres, L.E.P.; Kerbauy, G.B. High cytokinin accumulation following root tip excision changes the endogenous auxin-to-cytokinin ratio during root-to-shoot conversion in Catasetum fimbriatum Lindl (Orchidaceae). Plant Cell Rep. 1999, 18, 1002–1006. [Google Scholar] [CrossRef]
- Kareem, A.; Radhakrishnan, D.; Wang, X.; Bagavathiappan, S.; Trivedi, Z.B.; Sugimoto, K.; Xu, J.; Mähönen, A.P.; Prasad, K. Protocol: A method to study the direct reprogramming of lateral root primordia to fertile shoots. Plant Methods 2016, 12, 27. [Google Scholar] [CrossRef] [Green Version]
- Bigeard, J.; Hirt, H. Nuclear signaling of plant MAPKs. Front. Plant Sci. 2018, 9, 469. [Google Scholar] [CrossRef] [Green Version]
- Dóczi, R.; Bögre, L. The quest for MAP kinase substrates: Gaining momentum. Trends Plant Sci. 2018, 23, 918–932. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Mohanta, N.; Parida, P.; Bae, H. Mitogen Activated Protein Kinase (MPK) interacts with auxin influx carrier (OsAux/LAX1) involved in auxin signaling in plant. Biol. Proced. Online 2015, 17, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winnicki, K.; Żabka, A.; Polit, J.T.; Maszewski, J. Mitogen-activated protein kinases concentrate in the vicinity of chromosomes and may regulate directly cellular patterning in Vicia faba embryos. Planta 2018, 248, 307–322. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Chen, Y.; Luo, L.; Peck, S.C. Central roles and regulatory mechanisms of dual-specificity MAPK phosphatases in developmental and stress signaling. Front. Plant Sci. 2018, 871, 1697. [Google Scholar] [CrossRef] [PubMed]
- Bartels, S.; Besteiro, M.A.G.; Lang, D.; Ulm, R. Emerging functions for plant MAP kinase phosphatases. Trends Plant Sci. 2010, 15, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.D.; Callaham, D.A.; Gross, D.J.; Hepler, P.K. Free Ca2+ gradient in growing pollen tubes of lilium. J. Cell Sci. 1992, 101, 7–12. [Google Scholar]
- Jaffe, L.A.; Weisenseel, M.H.; Jaffe, L.F. Calcium accumulations within the growing tips of pollen tubes. J. Antibiot. (Tokyo) 1975, 67, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Zabka, A.; Polit, J.T.; Winnicki, K.; Paciorek, P.; Juszczak, J.; Nowak, M.; Maszewski, J. PIN2-like proteins may contribute to the regulation of morphogenetic processes during spermatogenesis in Chara vulgaris. Plant Cell Rep. 2016, 35, 1655–1669. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Sun, H.L.; Mei, C.; Wang, X.J.; Yan, L.; Liu, R.; Zhang, X.F.; Wang, X.F.; Zhang, D.P. The Arabidopsis Ca 2+-dependent protein kinase CPK12 negatively regulates abscisic acid signaling in seed germination and post-germination growth. New Phytol. 2011, 192, 61–73. [Google Scholar] [CrossRef]
- Inoue, S.I.; Takahashi, K.; Okumura-Noda, H.; Kinoshita, T. Auxin influx carrier AUX1 confers acid resistance for arabidopsis root elongation through the regulation of plasma membrane H+-ATPase. Plant Cell Physiol. 2016, 57, 2194–2201. [Google Scholar] [CrossRef] [Green Version]
- Maheshwari, P.; Du, H.; Sheen, J.; Assmann, S.M.; Albert, R. Model-driven discovery of calcium-related protein-phosphatase inhibition in plant guard cell signaling. PLoS Comput. Biol. 2019, 15, e1007429. [Google Scholar] [CrossRef]
- Rodriguez, P.L. Protein phosphatase 2C (PP2C) function in higher plants. Plant Mol. Biol. 1998, 38, 919–927. [Google Scholar] [CrossRef] [PubMed]
- Phengchat, R.; Takata, H.; Morii, K.; Inada, N.; Murakoshi, H.; Uchiyama, S.; Fukui, K. Calcium ions function as a booster of chromosome condensation. Sci. Rep. 2016, 6, 38281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hepler, P.K. Calcium: A central regulator of plant growth and development. Plant Cell 2005, 17, 2142–2155. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, L.F.; Robinson, K.R.; Nuccitelli, R. Local cation entry and self-electrophoresis as an intracellular localization mechanism. Ann. N. Y. Acad. Sci. 1974, 238, 372–389. [Google Scholar] [CrossRef] [PubMed]
- Raccaud, M.; Friman, E.T.; Alber, A.B.; Agarwal, H.; Deluz, C.; Kuhn, T.; Gebhardt, J.C.M.; Suter, D.M. Mitotic chromosome binding predicts transcription factor properties in interphase. Nat. Commun. 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raccaud, M.; Suter, D.M. Transcription factor retention on mitotic chromosomes: Regulatory mechanisms and impact on cell fate decisions. FEBS Lett. 2018, 592, 878–887. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winnicki, K. The Winner Takes It All: Auxin—The Main Player during Plant Embryogenesis. Cells 2020, 9, 606. https://doi.org/10.3390/cells9030606
Winnicki K. The Winner Takes It All: Auxin—The Main Player during Plant Embryogenesis. Cells. 2020; 9(3):606. https://doi.org/10.3390/cells9030606
Chicago/Turabian StyleWinnicki, Konrad. 2020. "The Winner Takes It All: Auxin—The Main Player during Plant Embryogenesis" Cells 9, no. 3: 606. https://doi.org/10.3390/cells9030606
APA StyleWinnicki, K. (2020). The Winner Takes It All: Auxin—The Main Player during Plant Embryogenesis. Cells, 9(3), 606. https://doi.org/10.3390/cells9030606