The Role of Tissue-resident γδ T Cells in Stress Surveillance and Tissue Maintenance

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Epidermal γδ T Cells in Maintenance and Repair
2.1. Epidermal Structure and Function
2.2. DETC Development and Trafficking to the Epidermis
2.3. DETC Functions in Homeostasis and Damage Repair
3. Intestinal Epithelial γδ T Cells in Maintenance and Repair
3.1. Structure and Function of the Intestinal Epithelium
3.2. γδ IEL Development and Trafficking to the Intestinal Epithelium
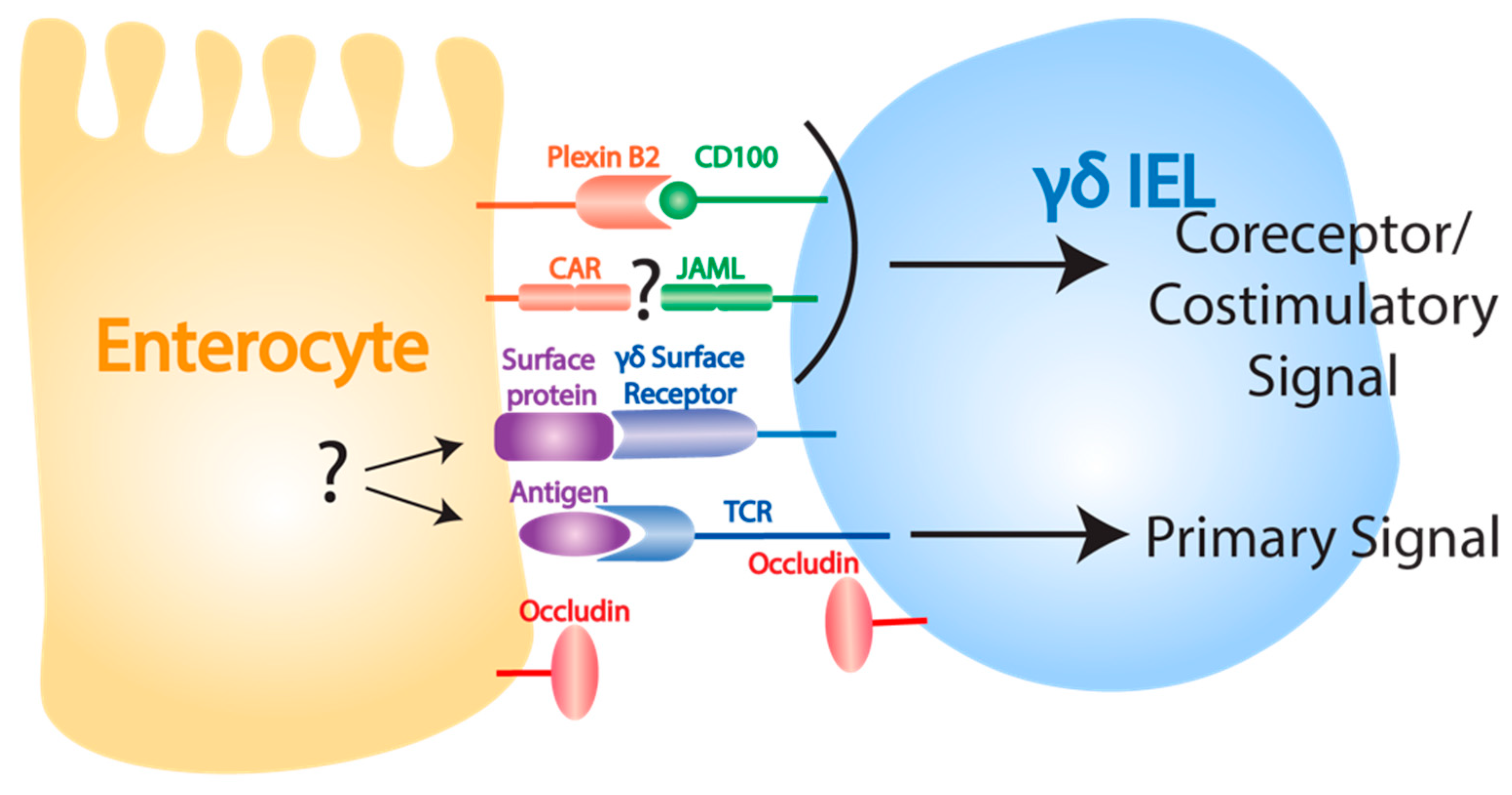
3.3. γδ IEL Functions in Homeostasis and Epithelial Damage Repair
4. Adipose Tissue-resident γδ T Cells in Inflammation and Obesity
4.1. Functions and Organization of Adipose Tissue
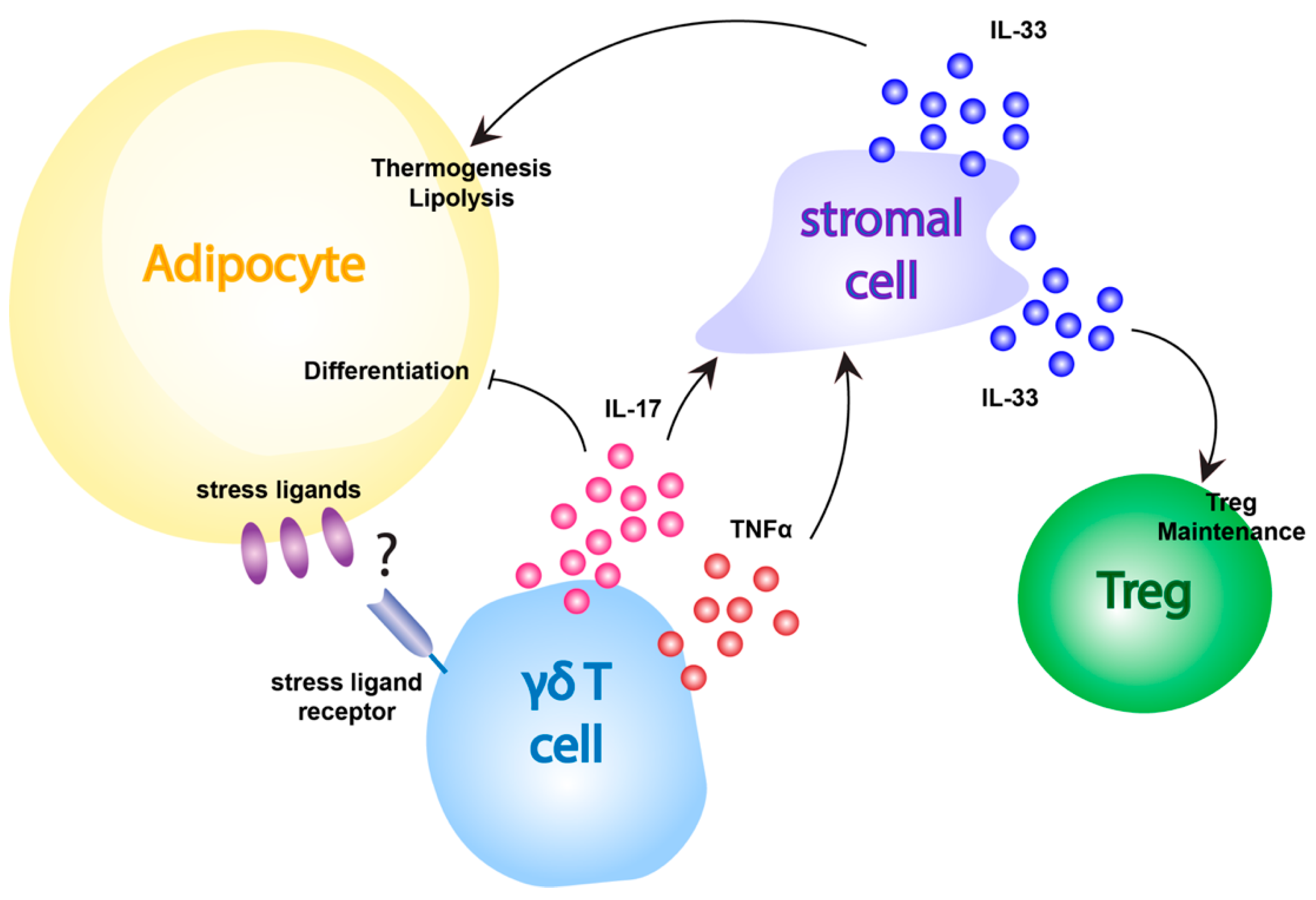
4.2. The Role of γδ T Cells in Obesity and Adipose Tissue Inflammation
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Bonneville, M.; Janeway, C.A., Jr.; Ito, K.; Haser, W.; Ishida, I.; Nakanishi, N.; Tonegawa, S. Intestinal intraepithelial lymphocytes are a distinct set of gamma delta T cells. Nature 1988, 336, 479–481. [Google Scholar] [CrossRef] [PubMed]
- Goodman, T.; Lefrancois, L. Expression of the gamma-delta T-cell receptor on intestinal CD8+ intraepithelial lymphocytes. Nature 1988, 333, 855–858. [Google Scholar] [CrossRef] [PubMed]
- Bucy, R.P.; Chen, C.L.; Cihak, J.; Losch, U.; Cooper, M.D. Avian T cells expressing gamma delta receptors localize in the splenic sinusoids and the intestinal epithelium. J. Immunol. 1988, 141, 2200–2205. [Google Scholar]
- Raulet, D.H. The structure, function, and molecular genetics of the gamma/delta T cell receptor. Annu. Rev. Immunol. 1989, 7, 175–207. [Google Scholar] [CrossRef]
- Koning, F.; Stingl, G.; Yokoyama, W.M.; Yamada, H.; Maloy, W.L.; Tschachler, E.; Shevach, E.M.; Coligan, J.E. Identification of a T3-associated gamma delta T cell receptor on Thy-1+ dendritic epidermal Cell lines. Science 1987, 236, 834–837. [Google Scholar] [CrossRef] [PubMed]
- Stingl, G.; Koning, F.; Yamada, H.; Yokoyama, W.M.; Tschachler, E.; Bluestone, J.A.; Steiner, G.; Samelson, L.E.; Lew, A.M.; Coligan, J.E.; et al. Thy-1+ dendritic epidermal cells express T3 antigen and the T-cell receptor gamma chain. Proc. Natl. Acad. Sci. USA 1987, 84, 4586–4590. [Google Scholar] [CrossRef] [Green Version]
- Krangel, M.S.; Yssel, H.; Brocklehurst, C.; Spits, H. A distinct wave of human T cell receptor gamma/delta lymphocytes in the early fetal thymus: Evidence for controlled gene rearrangement and cytokine production. J. Exp. Med. 1990, 172, 847–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heilig, J.S.; Tonegawa, S. Diversity of murine gamma genes and expression in fetal and adult T lymphocytes. Nature 1986, 322, 836–840. [Google Scholar] [CrossRef]
- Garman, R.D.; Doherty, P.J.; Raulet, D.H. Diversity, rearrangement, and expression of murine T cell gamma genes. Cell 1986, 45, 733–742. [Google Scholar] [CrossRef]
- Hayday, A.C. [gamma][delta] cells: A right time and a right place for a conserved third way of protection. Annu. Rev. Immunol. 2000, 18, 975–1026. [Google Scholar] [CrossRef]
- Caspar-Bauguil, S.; Cousin, B.; Galinier, A.; Segafredo, C.; Nibbelink, M.; Andre, M.; Casteilla, L.; Penicaud, L. Adipose tissues as an ancestral immune organ: Site-specific change in obesity. FEBS Lett. 2005, 579, 3487–3492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohlgruber, A.C.; Gal-Oz, S.T.; LaMarche, N.M.; Shimazaki, M.; Duquette, D.; Koay, H.F.; Nguyen, H.N.; Mina, A.I.; Paras, T.; Tavakkoli, A.; et al. Gammadelta T cells producing interleukin-17A regulate adipose regulatory T cell homeostasis and thermogenesis. Nat. Immunol. 2018, 19, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Sugahara, S.; Shimizu, T.; Yoshida, Y.; Aiba, T.; Yamagiwa, S.; Asakura, H.; Abo, T. Extrathymic derivation of gut lymphocytes in parabiotic mice. Immunology 1999, 96, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Park, C.O.; Geddes Sweeney, J.; Yoo, M.J.; Gaide, O.; Kupper, T.S. Dermal gammadelta T Cells Do Not Freely Re-Circulate Out of Skin and Produce IL-17 to Promote Neutrophil Infiltration during Primary Contact Hypersensitivity. PLoS ONE 2017, 12, e0169397. [Google Scholar] [CrossRef]
- Asarnow, D.M.; Goodman, T.; LeFrancois, L.; Allison, J.P. Distinct antigen receptor repertoires of two classes of murine epithelium-associated T cells. Nature 1989, 341, 60–62. [Google Scholar] [CrossRef]
- Havran, W.L.; Allison, J.P. Origin of Thy-1+ dendritic epidermal cells of adult mice from fetal thymic precursors. Nature 1990, 344, 68–70. [Google Scholar] [CrossRef]
- Itohara, S.; Farr, A.G.; Lafaille, J.J.; Bonneville, M.; Takagaki, Y.; Haas, W.; Tonegawa, S. Homing of a gamma delta thymocyte subset with homogeneous T-cell receptors to mucosal epithelia. Nature 1990, 343, 754–757. [Google Scholar] [CrossRef]
- Takagaki, Y.; DeCloux, A.; Bonneville, M.; Tonegawa, S. Diversity of gamma delta T-cell receptors on murine intestinal intra-epithelial lymphocytes. Nature 1989, 339, 712–714. [Google Scholar] [CrossRef]
- Havran, W.L.; Allison, J.P. Developmentally ordered appearance of thymocytes expressing different T-cell antigen receptors. Nature 1988, 335, 443–445. [Google Scholar] [CrossRef]
- Ito, K.; Bonneville, M.; Takagaki, Y.; Nakanishi, N.; Kanagawa, O.; Krecko, E.G.; Tonegawa, S. Different gamma delta T-cell receptors are expressed on thymocytes at different stages of development. Proc. Natl. Acad. Sci. USA 1989, 86, 631–635. [Google Scholar] [CrossRef] [Green Version]
- Xiong, N.; Baker, J.E.; Kang, C.; Raulet, D.H. The genomic arrangement of T cell receptor variable genes is a determinant of the developmental rearrangement pattern. Proc. Natl. Acad. Sci. USA 2004, 101, 260–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, Y.H.; Meyer, C.; Bonneville, M. gammadelta T cells: First line of defense and beyond. Annu. Rev. Immunol. 2014, 32, 121–155. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A.; Jones, B.; Hayday, A. Specificity and function of T cells bearing gamma delta receptors. Immunol. Today 1988, 9, 73–76. [Google Scholar] [CrossRef]
- Fuchs, E. Epithelial Skin Biology: Three Decades of Developmental Biology, a Hundred Questions Answered and a Thousand New Ones to Address. Curr. Top. Dev. Biol. 2016, 116, 357–374. [Google Scholar] [CrossRef] [Green Version]
- Wickett, R.R.; Visscher, M.O. Structure and function of the epidermal barrier. Am. J. Infect. Control 2006, 34, S98–S110. [Google Scholar] [CrossRef]
- Nestle, F.O.; Di Meglio, P.; Qin, J.Z.; Nickoloff, B.J. Skin immune sentinels in health and disease. Nat. Rev. Immunol. 2009, 9, 679–691. [Google Scholar] [CrossRef] [Green Version]
- Pasparakis, M.; Haase, I.; Nestle, F.O. Mechanisms regulating skin immunity and inflammation. Nat. Rev. Immunol. 2014, 14, 289–301. [Google Scholar] [CrossRef]
- Di Meglio, P.; Perera, G.K.; Nestle, F.O. The multitasking organ: Recent insights into skin immune function. Immunity 2011, 35, 857–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toulon, A.; Breton, L.; Taylor, K.R.; Tenenhaus, M.; Bhavsar, D.; Lanigan, C.; Rudolph, R.; Jameson, J.; Havran, W.L. A role for human skin-resident T cells in wound healing. J. Exp. Med. 2009, 206, 743–750. [Google Scholar] [CrossRef] [Green Version]
- Elbe, A.; Foster, C.A.; Stingl, G. T-cell receptor alpha beta and gamma delta T cells in rat and human skin--are they equivalent? Semin. Immunol. 1996, 8, 341–349. [Google Scholar] [CrossRef]
- Dupuy, P.; Heslan, M.; Fraitag, S.; Hercend, T.; Dubertret, L.; Bagot, M. T-cell receptor-gamma/delta bearing lymphocytes in normal and inflammatory human skin. J. Investig. Dermatol. 1990, 94, 764–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebert, L.M.; Meuter, S.; Moser, B. Homing and function of human skin gammadelta T cells and NK cells: Relevance for tumor surveillance. J. Immunol. 2006, 176, 4331–4336. [Google Scholar] [CrossRef] [PubMed]
- Holtmeier, W.; Pfander, M.; Hennemann, A.; Zollner, T.M.; Kaufmann, R.; Caspary, W.F. The TCR-delta repertoire in normal human skin is restricted and distinct from the TCR-delta repertoire in the peripheral blood. J. Investig. Dermatol. 2001, 116, 275–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtmeier, W.; Pfander, M.; Zollner, T.M.; Kaufmann, R.; Caspary, W.F. Distinct TCR delta repertoires are present in the cutaneous lesions and inflamed duodenum of patients with dermatitis herpetiformis. Exp. Dermatol. 2002, 11, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Havran, W.L.; Chien, Y.H.; Allison, J.P. Recognition of self antigens by skin-derived T cells with invariant gamma delta antigen receptors. Science 1991, 252, 1430–1432. [Google Scholar] [CrossRef]
- Strid, J.; Roberts, S.J.; Filler, R.B.; Lewis, J.M.; Kwong, B.Y.; Schpero, W.; Kaplan, D.H.; Hayday, A.C.; Girardi, M. Acute upregulation of an NKG2D ligand promotes rapid reorganization of a local immune compartment with pleiotropic effects on carcinogenesis. Nat. Immunol. 2008, 9, 146–154. [Google Scholar] [CrossRef]
- Xiong, N.; Kang, C.; Raulet, D.H. Positive selection of dendritic epidermal gammadelta T cell precursors in the fetal thymus determines expression of skin-homing receptors. Immunity 2004, 21, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Aono, A.; Enomoto, H.; Yoshida, N.; Yoshizaki, K.; Kishimoto, T.; Komori, T. Forced expression of terminal deoxynucleotidyl transferase in fetal thymus resulted in a decrease in gammadelta T cells and random dissemination of Vgamma3Vdelta1 T cells in skin of newborn but not adult mice. Immunology 2000, 99, 489–497. [Google Scholar] [CrossRef]
- Jameson, J.M.; Cauvi, G.; Witherden, D.A.; Havran, W.L. A keratinocyte-responsive gamma delta TCR is necessary for dendritic epidermal T cell activation by damaged keratinocytes and maintenance in the epidermis. J. Immunol. 2004, 172, 3573–3579. [Google Scholar] [CrossRef] [Green Version]
- Mallick-Wood, C.A.; Lewis, J.M.; Richie, L.I.; Owen, M.J.; Tigelaar, R.E.; Hayday, A.C. Conservation of T cell receptor conformation in epidermal gammadelta cells with disrupted primary Vgamma gene usage. Science 1998, 279, 1729–1733. [Google Scholar] [CrossRef]
- Hara, H.; Kishihara, K.; Matsuzaki, G.; Takimoto, H.; Tsukiyama, T.; Tigelaar, R.E.; Nomoto, K. Development of dendritic epidermal T cells with a skewed diversity of gamma delta TCRs in V delta 1-deficient mice. J. Immunol. 2000, 165, 3695–3705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Xia, M.; Sun, A.; Saylor, C.M.; Xiong, N. CCR10 is important for the development of skin-specific gammadeltaT cells by regulating their migration and location. J. Immunol. 2010, 185, 5723–5731. [Google Scholar] [CrossRef]
- Xia, M.; Qi, Q.; Jin, Y.; Wiest, D.L.; August, A.; Xiong, N. Differential roles of IL-2-inducible T cell kinase-mediated TCR signals in tissue-specific localization and maintenance of skin intraepithelial T cells. J. Immunol. 2010, 184, 6807–6814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Campbell, J.J.; Kupper, T.S. Embryonic trafficking of gammadelta T cells to skin is dependent on E/P selectin ligands and CCR4. Proc. Natl. Acad. Sci. USA 2010, 107, 7443–7448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Creus, A.; Van Beneden, K.; Stevenaert, F.; Debacker, V.; Plum, J.; Leclercq, G. Developmental and functional defects of thymic and epidermal V gamma 3 cells in IL-15-deficient and IFN regulatory factor-1-deficient mice. J. Immunol. 2002, 168, 6486–6493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, K.; Sunaga, S.; Komagata, Y.; Kodaira, Y.; Mabuchi, A.; Karasuyama, H.; Yokomuro, K.; Miyazaki, J.I.; Ikuta, K. Interleukin 7 receptor-deficient mice lack gammadelta T cells. Proc. Natl. Acad. Sci. USA 1996, 93, 7172–7177. [Google Scholar] [CrossRef] [Green Version]
- Ye, S.K.; Maki, K.; Lee, H.C.; Ito, A.; Kawai, K.; Suzuki, H.; Mak, T.W.; Chien, Y.; Honjo, T.; Ikuta, K. Differential roles of cytokine receptors in the development of epidermal gamma delta T cells. J. Immunol. 2001, 167, 1929–1934. [Google Scholar] [CrossRef] [Green Version]
- Lewis, J.M.; Girardi, M.; Roberts, S.J.; Barbee, S.D.; Hayday, A.C.; Tigelaar, R.E. Selection of the cutaneous intraepithelial gammadelta+ T cell repertoire by a thymic stromal determinant. Nat. Immunol. 2006, 7, 843–850. [Google Scholar] [CrossRef]
- Boyden, L.M.; Lewis, J.M.; Barbee, S.D.; Bas, A.; Girardi, M.; Hayday, A.C.; Tigelaar, R.E.; Lifton, R.P. Skint1, the prototype of a newly identified immunoglobulin superfamily gene cluster, positively selects epidermal gammadelta T cells. Nat. Genet. 2008, 40, 656–662. [Google Scholar] [CrossRef] [Green Version]
- Turchinovich, G.; Hayday, A.C. Skint-1 identifies a common molecular mechanism for the development of interferon-gamma-secreting versus interleukin-17-secreting gammadelta T cells. Immunity 2011, 35, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Macleod, A.S.; Havran, W.L. Functions of skin-resident gammadelta T cells. Cell Mol. Life Sci. 2011, 68, 2399–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jameson, J.; Havran, W.L. Skin gammadelta T-cell functions in homeostasis and wound healing. Immunol. Rev. 2007, 215, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Chodaczek, G.; Papanna, V.; Zal, M.A.; Zal, T. Body-barrier surveillance by epidermal gammadelta TCRs. Nat. Immunol. 2012, 13, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Girardi, M.; Lewis, J.M.; Filler, R.B.; Hayday, A.C.; Tigelaar, R.E. Environmentally responsive and reversible regulation of epidermal barrier function by gammadelta T cells. J. Investig. Dermatol. 2006, 126, 808–814. [Google Scholar] [CrossRef] [Green Version]
- Sharp, L.L.; Jameson, J.M.; Cauvi, G.; Havran, W.L. Dendritic epidermal T cells regulate skin homeostasis through local production of insulin-like growth factor 1. Nat. Immunol. 2005, 6, 73–79. [Google Scholar] [CrossRef]
- Boismenu, R.; Feng, L.; Xia, Y.Y.; Chang, J.C.; Havran, W.L. Chemokine expression by intraepithelial gamma delta T cells. Implications for the recruitment of inflammatory cells to damaged epithelia. J. Immunol. 1996, 157, 985–992. [Google Scholar]
- Girardi, M.; Lewis, J.; Glusac, E.; Filler, R.B.; Geng, L.; Hayday, A.C.; Tigelaar, R.E. Resident skin-specific gammadelta T cells provide local, nonredundant regulation of cutaneous inflammation. J. Exp. Med. 2002, 195, 855–867. [Google Scholar] [CrossRef] [Green Version]
- Havran, W.L.; Jameson, J.M. Epidermal T cells and wound healing. J. Immunol. 2010, 184, 5423–5428. [Google Scholar] [CrossRef]
- Jameson, J.; Ugarte, K.; Chen, N.; Yachi, P.; Fuchs, E.; Boismenu, R.; Havran, W.L. A role for skin gammadelta T cells in wound repair. Science 2002, 296, 747–749. [Google Scholar] [CrossRef]
- Keyes, B.E.; Liu, S.; Asare, A.; Naik, S.; Levorse, J.; Polak, L.; Lu, C.P.; Nikolova, M.; Pasolli, H.A.; Fuchs, E. Impaired Epidermal to Dendritic T Cell Signaling Slows Wound Repair in Aged Skin. Cell 2016, 167, 1323–1338; e1314. [Google Scholar] [CrossRef] [Green Version]
- Jameson, J.M.; Cauvi, G.; Sharp, L.L.; Witherden, D.A.; Havran, W.L. Gammadelta T cell-induced hyaluronan production by epithelial cells regulates inflammation. J. Exp. Med. 2005, 201, 1269–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLeod, A.S.; Hemmers, S.; Garijo, O.; Chabod, M.; Mowen, K.; Witherden, D.A.; Havran, W.L. Dendritic epidermal T cells regulate skin antimicrobial barrier function. J. Clin. Investig. 2013, 123, 4364–4374. [Google Scholar] [CrossRef] [PubMed]
- Komori, H.K.; Witherden, D.A.; Kelly, R.; Sendaydiego, K.; Jameson, J.M.; Teyton, L.; Havran, W.L. Cutting edge: Dendritic epidermal gammadelta T cell ligands are rapidly and locally expressed by keratinocytes following cutaneous wounding. J. Immunol. 2012, 188, 2972–2976. [Google Scholar] [CrossRef] [PubMed]
- Witherden, D.A.; Verdino, P.; Rieder, S.E.; Garijo, O.; Mills, R.E.; Teyton, L.; Fischer, W.H.; Wilson, I.A.; Havran, W.L. The junctional adhesion molecule JAML is a costimulatory receptor for epithelial gammadelta T cell activation. Science 2010, 329, 1205–1210. [Google Scholar] [CrossRef] [Green Version]
- Verdino, P.; Witherden, D.A.; Havran, W.L.; Wilson, I.A. The molecular interaction of CAR and JAML recruits the central cell signal transducer PI3K. Science 2010, 329, 1210–1214. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Mohamed, R.H.; Kajikawa, M.; Koizumi, J.; Tanaka, M.; Fugo, K.; Otsuka, N.; Maenaka, K.; Yagita, H.; Chiba, H.; et al. Involvement of an NKG2D ligand H60c in epidermal dendritic T cell-mediated wound repair. J. Immunol. 2012, 188, 3972–3979. [Google Scholar] [CrossRef] [Green Version]
- Whang, M.I.; Guerra, N.; Raulet, D.H. Costimulation of dendritic epidermal gammadelta T cells by a new NKG2D ligand expressed specifically in the skin. J. Immunol. 2009, 182, 4557–4564. [Google Scholar] [CrossRef] [Green Version]
- Witherden, D.A.; Watanabe, M.; Garijo, O.; Rieder, S.E.; Sarkisyan, G.; Cronin, S.J.; Verdino, P.; Wilson, I.A.; Kumanogoh, A.; Kikutani, H.; et al. The CD100 receptor interacts with its plexin B2 ligand to regulate epidermal gammadelta T cell function. Immunity 2012, 37, 314–325. [Google Scholar] [CrossRef] [Green Version]
- Girardi, M.; Oppenheim, D.E.; Steele, C.R.; Lewis, J.M.; Glusac, E.; Filler, R.; Hobby, P.; Sutton, B.; Tigelaar, R.E.; Hayday, A.C. Regulation of cutaneous malignancy by gammadelta T cells. Science 2001, 294, 605–609. [Google Scholar] [CrossRef]
- MacLeod, A.S.; Rudolph, R.; Corriden, R.; Ye, I.; Garijo, O.; Havran, W.L. Skin-resident T cells sense ultraviolet radiation-induced injury and contribute to DNA repair. J. Immunol. 2014, 192, 5695–5702. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, M.M.; Lovato, P.; MacLeod, A.S.; Witherden, D.A.; Skov, L.; Dyring-Andersen, B.; Dabelsteen, S.; Woetmann, A.; Odum, N.; Havran, W.L.; et al. IL-1beta-dependent activation of dendritic epidermal T cells in contact hypersensitivity. J. Immunol. 2014, 192, 2975–2983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, M.M.; Dyring-Andersen, B.; Schmidt, J.D.; Witherden, D.; Lovato, P.; Woetmann, A.; Odum, N.; Poulsen, S.S.; Havran, W.L.; Geisler, C.; et al. NKG2D-dependent activation of dendritic epidermal T cells in contact hypersensitivity. J. Investig. Dermatol. 2015, 135, 1311–1319. [Google Scholar] [CrossRef] [Green Version]
- van der Flier, L.G.; Clevers, H. Stem cells, self-renewal, and differentiation in the intestinal epithelium. Annu. Rev. Physiol. 2009, 71, 241–260. [Google Scholar] [CrossRef] [PubMed]
- Crosnier, C.; Stamataki, D.; Lewis, J. Organizing cell renewal in the intestine: Stem cells, signals and combinatorial control. Nat. Rev. Genet. 2006, 7, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Allaire, J.M.; Crowley, S.M.; Law, H.T.; Chang, S.Y.; Ko, H.J.; Vallance, B.A. The Intestinal Epithelium: Central Coordinator of Mucosal Immunity. Trends Immunol. 2018, 39, 677–696. [Google Scholar] [CrossRef] [PubMed]
- Okumura, R.; Takeda, K. Roles of intestinal epithelial cells in the maintenance of gut homeostasis. Exp. Mol. Med. 2017, 49, e338. [Google Scholar] [CrossRef] [Green Version]
- Kurashima, Y.; Kiyono, H. Mucosal Ecological Network of Epithelium and Immune Cells for Gut Homeostasis and Tissue Healing. Annu. Rev. Immunol. 2017, 35, 119–147. [Google Scholar] [CrossRef]
- Darlington, D.; Rogers, A.W. Epithelial lymphocytes in the small intestine of the mouse. J. Anat. 1966, 100, 813–830. [Google Scholar]
- Ferguson, A. Intraepithelial lymphocytes of the small intestine. Gut 1977, 18, 921–937. [Google Scholar] [CrossRef] [Green Version]
- Cheroutre, H. IELs: Enforcing law and order in the court of the intestinal epithelium. Immunol. Rev. 2005, 206, 114–131. [Google Scholar] [CrossRef]
- Hayday, A.; Theodoridis, E.; Ramsburg, E.; Shires, J. Intraepithelial lymphocytes: Exploring the Third Way in immunology. Nat. Immunol. 2001, 2, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Cheroutre, H.; Madakamutil, L. Acquired and natural memory T cells join forces at the mucosal front line. Nat. Rev. Immunol. 2004, 4, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Guy-Grand, D.; Cerf-Bensussan, N.; Malissen, B.; Malassis-Seris, M.; Briottet, C.; Vassalli, P. Two gut intraepithelial CD8+ lymphocyte populations with different T cell receptors: A role for the gut epithelium in T cell differentiation. J. Exp. Med. 1991, 173, 471–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camerini, V.; Panwala, C.; Kronenberg, M. Regional specialization of the mucosal immune system. Intraepithelial lymphocytes of the large intestine have a different phenotype and function than those of the small intestine. J. Immunol. 1993, 151, 1765–1776. [Google Scholar] [PubMed]
- Pereira, P.; Hermitte, V.; Lembezat, M.P.; Boucontet, L.; Azuara, V.; Grigoriadou, K. Developmentally regulated and lineage-specific rearrangement of T cell receptor Valpha/delta gene segments. Eur. J. Immunol. 2000, 30, 1988–1997. [Google Scholar] [CrossRef]
- Xiong, N.; Raulet, D.H. Development and selection of gammadelta T cells. Immunol. Rev. 2007, 215, 15–31. [Google Scholar] [CrossRef]
- Ishikawa, H.; Naito, T.; Iwanaga, T.; Takahashi-Iwanaga, H.; Suematsu, M.; Hibi, T.; Nanno, M. Curriculum vitae of intestinal intraepithelial T cells: Their developmental and behavioral characteristics. Immunol. Rev. 2007, 215, 154–165. [Google Scholar] [CrossRef]
- Bandeira, A.; Itohara, S.; Bonneville, M.; Burlen-Defranoux, O.; Mota-Santos, T.; Coutinho, A.; Tonegawa, S. Extrathymic origin of intestinal intraepithelial lymphocytes bearing T-cell antigen receptor gamma delta. Proc. Natl. Acad. Sci. USA 1991, 88, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Guy-Grand, D.; Azogui, O.; Celli, S.; Darche, S.; Nussenzweig, M.C.; Kourilsky, P.; Vassalli, P. Extrathymic T cell lymphopoiesis: Ontogeny and contribution to gut intraepithelial lymphocytes in athymic and euthymic mice. J. Exp. Med. 2003, 197, 333–341. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Nguyen, H.; Kang, J. Interleukin 15 controls the generation of the restricted T cell receptor repertoire of gamma delta intestinal intraepithelial lymphocytes. Nat. Immunol. 2005, 6, 1263–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Duncan, G.S.; Takimoto, H.; Mak, T.W. Abnormal development of intestinal intraepithelial lymphocytes and peripheral natural killer cells in mice lacking the IL-2 receptor beta chain. J. Exp. Med. 1997, 185, 499–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodolce, J.P.; Boone, D.L.; Chai, S.; Swain, R.E.; Dassopoulos, T.; Trettin, S.; Ma, A. IL-15 receptor maintains lymphoid homeostasis by supporting lymphocyte homing and proliferation. Immunity 1998, 9, 669–676. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.J.; Acero, L.F.; Zal, T.; Schluns, K.S. Trans-presentation of IL-15 by intestinal epithelial cells drives development of CD8alphaalpha IELs. J. Immunol. 2009, 183, 1044–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki-Ohara, K.; Nishimura, H.; Mitani, A.; Yoshikai, Y. Interleukin-15 preferentially promotes the growth of intestinal intraepithelial lymphocytes bearing gamma delta T cell receptor in mice. Eur. J. Immunol. 1997, 27, 2885–2891. [Google Scholar] [CrossRef]
- Zhu, Y.; Cui, G.; Miyauchi, E.; Nakanishi, Y.; Mukohira, H.; Shimba, A.; Abe, S.; Tani-Ichi, S.; Hara, T.; Nakase, H.; et al. Intestinal epithelial cell-derived IL-15 determines local maintenance and maturation of intraepithelial lymphocytes in the intestine. Int. Immunol. 2019, dxz082. [Google Scholar] [CrossRef]
- Lai, Y.G.; Hou, M.S.; Hsu, Y.W.; Chang, C.L.; Liou, Y.H.; Tsai, M.H.; Lee, F.; Liao, N.S. IL-15 does not affect IEL development in the thymus but regulates homeostasis of putative precursors and mature CD8 alpha alpha+ IELs in the intestine. J. Immunol. 2008, 180, 3757–3765. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.D.; Ethridge, A.D.; Lipstein, R.; Kumar, S.; Wang, Y.; Jabri, B.; Turner, J.R.; Edelblum, K.L. Epithelial IL-15 Is a Critical Regulator of gammadelta Intraepithelial Lymphocyte Motility within the Intestinal Mucosa. J. Immunol. 2018, 201, 747–756. [Google Scholar] [CrossRef] [Green Version]
- Wurbel, M.A.; Philippe, J.M.; Nguyen, C.; Victorero, G.; Freeman, T.; Wooding, P.; Miazek, A.; Mattei, M.G.; Malissen, M.; Jordan, B.R.; et al. The chemokine TECK is expressed by thymic and intestinal epithelial cells and attracts double- and single-positive thymocytes expressing the TECK receptor CCR9. Eur. J. Immunol. 2000, 30, 262–271. [Google Scholar] [CrossRef]
- Wurbel, M.A.; Malissen, M.; Guy-Grand, D.; Meffre, E.; Nussenzweig, M.C.; Richelme, M.; Carrier, A.; Malissen, B. Mice lacking the CCR9 CC-chemokine receptor show a mild impairment of early T- and B-cell development and a reduction in T-cell receptor gammadelta(+) gut intraepithelial lymphocytes. Blood 2001, 98, 2626–2632. [Google Scholar] [CrossRef]
- Wurbel, M.A.; Malissen, M.; Guy-Grand, D.; Malissen, B.; Campbell, J.J. Impaired accumulation of antigen-specific CD8 lymphocytes in chemokine CCL25-deficient intestinal epithelium and lamina propria. J. Immunol. 2007, 178, 7598–7606. [Google Scholar] [CrossRef] [Green Version]
- Guehler, S.R.; Bluestone, J.A.; Barrett, T.A. Activation and peripheral expansion of murine T-cell receptor gamma delta intraepithelial lymphocytes. Gastroenterology 1999, 116, 327–334. [Google Scholar] [CrossRef]
- Jensen, K.D.; Shin, S.; Chien, Y.H. Cutting edge: Gammadelta intraepithelial lymphocytes of the small intestine are not biased toward thymic antigens. J. Immunol. 2009, 182, 7348–7351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chennupati, V.; Worbs, T.; Liu, X.; Malinarich, F.H.; Schmitz, S.; Haas, J.D.; Malissen, B.; Forster, R.; Prinz, I. Intra- and intercompartmental movement of gammadelta T cells: Intestinal intraepithelial and peripheral gammadelta T cells represent exclusive nonoverlapping populations with distinct migration characteristics. J. Immunol. 2010, 185, 5160–5168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelblum, K.L.; Shen, L.; Weber, C.R.; Marchiando, A.M.; Clay, B.S.; Wang, Y.; Prinz, I.; Malissen, B.; Sperling, A.I.; Turner, J.R. Dynamic migration of gammadelta intraepithelial lymphocytes requires occludin. Proc. Natl. Acad. Sci. USA 2012, 109, 7097–7102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelblum, K.L.; Singh, G.; Odenwald, M.A.; Lingaraju, A.; El Bissati, K.; McLeod, R.; Sperling, A.I.; Turner, J.R. gammadelta Intraepithelial Lymphocyte Migration Limits Transepithelial Pathogen Invasion and Systemic Disease in Mice. Gastroenterology 2015, 148, 1417–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, M.M.; Witherden, D.A.; Havran, W.L. gammadelta T cells in homeostasis and host defence of epithelial barrier tissues. Nat. Rev. Immunol. 2017, 17, 733–745. [Google Scholar] [CrossRef]
- Khairallah, C.; Chu, T.H.; Sheridan, B.S. Tissue Adaptations of Memory and Tissue-Resident Gamma Delta T Cells. Front. Immunol. 2018, 9, 2636. [Google Scholar] [CrossRef]
- Shires, J.; Theodoridis, E.; Hayday, A.C. Biological insights into TCRgammadelta+ and TCRalphabeta+ intraepithelial lymphocytes provided by serial analysis of gene expression (SAGE). Immunity 2001, 15, 419–434. [Google Scholar] [CrossRef] [Green Version]
- Fahrer, A.M.; Konigshofer, Y.; Kerr, E.M.; Ghandour, G.; Mack, D.H.; Davis, M.M.; Chien, Y.H. Attributes of gammadelta intraepithelial lymphocytes as suggested by their transcriptional profile. Proc. Natl. Acad. Sci. USA 2001, 98, 10261–10266. [Google Scholar] [CrossRef] [Green Version]
- van Wijk, F.; Cheroutre, H. Mucosal T cells in gut homeostasis and inflammation. Expert Rev. Clin. Immunol. 2010, 6, 559–566. [Google Scholar] [CrossRef] [Green Version]
- Born, W.; Cady, C.; Jones-Carson, J.; Mukasa, A.; Lahn, M.; O’Brien, R. Immunoregulatory functions of gamma delta T cells. Adv. Immunol. 1999, 71, 77–144. [Google Scholar] [PubMed]
- Kuhl, A.A.; Pawlowski, N.N.; Grollich, K.; Loddenkemper, C.; Zeitz, M.; Hoffmann, J.C. Aggravation of intestinal inflammation by depletion/deficiency of gammadelta T cells in different types of IBD animal models. J. Leukoc. Biol. 2007, 81, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Inagaki-Ohara, K.; Chinen, T.; Matsuzaki, G.; Sasaki, A.; Sakamoto, Y.; Hiromatsu, K.; Nakamura-Uchiyama, F.; Nawa, Y.; Yoshimura, A. Mucosal T cells bearing TCRgammadelta play a protective role in intestinal inflammation. J. Immunol. 2004, 173, 1390–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, S.J.; Smith, A.L.; West, A.B.; Wen, L.; Findly, R.C.; Owen, M.J.; Hayday, A.C. T-cell alpha beta + and gamma delta + deficient mice display abnormal but distinct phenotypes toward a natural, widespread infection of the intestinal epithelium. Proc. Natl. Acad. Sci. USA 1996, 93, 11774–11779. [Google Scholar] [CrossRef] [Green Version]
- Bhagat, G.; Naiyer, A.J.; Shah, J.G.; Harper, J.; Jabri, B.; Wang, T.C.; Green, P.H.; Manavalan, J.S. Small intestinal CD8+TCRgammadelta+NKG2A+ intraepithelial lymphocytes have attributes of regulatory cells in patients with celiac disease. J. Clin. Investig. 2008, 118, 281–293. [Google Scholar] [CrossRef]
- Komano, H.; Fujiura, Y.; Kawaguchi, M.; Matsumoto, S.; Hashimoto, Y.; Obana, S.; Mombaerts, P.; Tonegawa, S.; Yamamoto, H.; Itohara, S.; et al. Homeostatic regulation of intestinal epithelia by intraepithelial gamma delta T cells. Proc. Natl. Acad. Sci. USA 1995, 92, 6147–6151. [Google Scholar] [CrossRef] [Green Version]
- Boismenu, R.; Havran, W.L. Modulation of epithelial cell growth by intraepithelial gamma delta T cells. Science 1994, 266, 1253–1255. [Google Scholar] [CrossRef]
- Yang, H.; Antony, P.; Wildhaber, B.; Teitelbaum, D. Intestinal Intraepithelial Lymphocyte gamma delta T cell-derived keratinocyte growth factor modulates epithelial growth in the mouse. J. Immunol. 2004, 172, 4151–4158. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chou, K.; Fuchs, E.; Havran, W.L.; Boismenu, R. Protection of the intestinal mucosa by intraepithelial gamma delta T cells. Proc. Natl. Acad. Sci. USA 2002, 99, 14338–14343. [Google Scholar] [CrossRef] [Green Version]
- Dalton, J.E.; Cruickshank, S.M.; Egan, C.E.; Mears, R.; Newton, D.J.; Andrew, E.M.; Lawrence, B.; Howell, G.; Else, K.J.; Gubbels, M.J.; et al. Intraepithelial gammadelta+ lymphocytes maintain the integrity of intestinal epithelial tight junctions in response to infection. Gastroenterology 2006, 131, 818–829. [Google Scholar] [CrossRef]
- Weitkamp, J.H.; Rosen, M.J.; Zhao, Z.; Koyama, T.; Geem, D.; Denning, T.L.; Rock, M.T.; Moore, D.J.; Halpern, M.D.; Matta, P.; et al. Small intestinal intraepithelial TCRgammadelta+ T lymphocytes are present in the premature intestine but selectively reduced in surgical necrotizing enterocolitis. PLoS ONE 2014, 9, e99042. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.; Isaacson, P.G.; Diss, T.C.; MacDonald, T.T. Expression of disulfide-linked and non-disulfide-linked forms of the T cell receptor gamma/delta heterodimer in human intestinal intraepithelial lymphocytes. Eur. J. Immunol. 1989, 19, 1335–1338. [Google Scholar] [CrossRef] [PubMed]
- Kutlu, T.; Brousse, N.; Rambaud, C.; Le Deist, F.; Schmitz, J.; Cerf-Bensussan, N. Numbers of T cell receptor (TCR) alpha beta+ but not of TcR gamma delta+ intraepithelial lymphocytes correlate with the grade of villous atrophy in coeliac patients on a long term normal diet. Gut 1993, 34, 208–214. [Google Scholar] [CrossRef] [Green Version]
- Mayassi, T.; Ladell, K.; Gudjonson, H.; McLaren, J.E.; Shaw, D.G.; Tran, M.T.; Rokicka, J.J.; Lawrence, I.; Grenier, J.C.; van Unen, V.; et al. Chronic Inflammation Permanently Reshapes Tissue-Resident Immunity in Celiac Disease. Cell 2019, 176, 967–981; e919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bucht, A.; Soderstrom, K.; Esin, S.; Grunewald, J.; Hagelberg, S.; Magnusson, I.; Wigzell, H.; Gronberg, A.; Kiessling, R. Analysis of gamma delta V region usage in normal and diseased human intestinal biopsies and peripheral blood by polymerase chain reaction (PCR) and flow cytometry. Clin. Exp. Immunol. 1995, 99, 57–64. [Google Scholar] [CrossRef]
- McVay, L.D.; Li, B.; Biancaniello, R.; Creighton, M.A.; Bachwich, D.; Lichtenstein, G.; Rombeau, J.L.; Carding, S.R. Changes in human mucosal gamma delta T cell repertoire and function associated with the disease process in inflammatory bowel disease. Mol. Med. 1997, 3, 183–203. [Google Scholar] [CrossRef] [Green Version]
- Hoytema van Konijnenburg, D.P.; Reis, B.S.; Pedicord, V.A.; Farache, J.; Victora, G.D.; Mucida, D. Intestinal Epithelial and Intraepithelial T Cell Crosstalk Mediates a Dynamic Response to Infection. Cell 2017, 171, 783–794; e713. [Google Scholar] [CrossRef]
- Meehan, T.F.; Witherden, D.A.; Kim, C.H.; Sendaydiego, K.; Ye, I.; Garijo, O.; Komori, H.K.; Kumanogoh, A.; Kikutani, H.; Eckmann, L.; et al. Protection against colitis by CD100-dependent modulation of intraepithelial gammadelta T lymphocyte function. Mucosal Immunol. 2014, 7, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Pazirandeh, A.; Sultana, T.; Mirza, M.; Rozell, B.; Hultenby, K.; Wallis, K.; Vennstrom, B.; Davis, B.; Arner, A.; Heuchel, R.; et al. Multiple phenotypes in adult mice following inactivation of the Coxsackievirus and Adenovirus Receptor (Car) gene. PLoS ONE 2011, 6, e20203. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.S.; Severson, K.M.; Vaishnava, S.; Behrendt, C.L.; Yu, X.; Benjamin, J.L.; Ruhn, K.A.; Hou, B.; DeFranco, A.L.; Yarovinsky, F.; et al. Gammadelta intraepithelial lymphocytes are essential mediators of host-microbial homeostasis at the intestinal mucosal surface. Proc. Natl. Acad. Sci. USA 2011, 108, 8743–8748. [Google Scholar] [CrossRef] [Green Version]
- Trayhurn, P. Adipocyte biology. Obes Rev. 2007, 8 (Suppl. 1), 41–44. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Latif, N.; Yacoub, M. Adipose tissue: Friend or foe? Nat. Rev. Cardiol 2012, 9, 689–702. [Google Scholar] [CrossRef] [PubMed]
- Zwick, R.K.; Guerrero-Juarez, C.F.; Horsley, V.; Plikus, M.V. Anatomical, Physiological, and Functional Diversity of Adipose Tissue. Cell Metab. 2018, 27, 68–83. [Google Scholar] [CrossRef] [PubMed]
- Tchkonia, T.; Thomou, T.; Zhu, Y.; Karagiannides, I.; Pothoulakis, C.; Jensen, M.D.; Kirkland, J.L. Mechanisms and metabolic implications of regional differences among fat depots. Cell Metab. 2013, 17, 644–656. [Google Scholar] [CrossRef] [Green Version]
- Shuster, A.; Patlas, M.; Pinthus, J.H.; Mourtzakis, M. The clinical importance of visceral adiposity: A critical review of methods for visceral adipose tissue analysis. Br. J. Radiol. 2012, 85, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Spiegelman, B.M. What we talk about when we talk about fat. Cell 2014, 156, 20–44. [Google Scholar] [CrossRef] [Green Version]
- Exley, M.A.; Hand, L.; O’Shea, D.; Lynch, L. Interplay between the immune system and adipose tissue in obesity. J. Endocrinol. 2014, 223, R41–R48. [Google Scholar] [CrossRef] [Green Version]
- Winer, S.; Winer, D.A. The adaptive immune system as a fundamental regulator of adipose tissue inflammation and insulin resistance. Immunol. Cell Biol. 2012, 90, 755–762. [Google Scholar] [CrossRef]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Wensveen, F.M.; Valentic, S.; Sestan, M.; Turk Wensveen, T.; Polic, B. The “Big Bang” in obese fat: Events initiating obesity-induced adipose tissue inflammation. Eur. J. Immunol. 2015, 45, 2446–2456. [Google Scholar] [CrossRef]
- Wu, H.; Ghosh, S.; Perrard, X.D.; Feng, L.; Garcia, G.E.; Perrard, J.L.; Sweeney, J.F.; Peterson, L.E.; Chan, L.; Smith, C.W.; et al. T-cell accumulation and regulated on activation, normal T cell expressed and secreted upregulation in adipose tissue in obesity. Circulation 2007, 115, 1029–1038. [Google Scholar] [CrossRef] [Green Version]
- Rausch, M.E.; Weisberg, S.; Vardhana, P.; Tortoriello, D.V. Obesity in C57BL/6J mice is characterized by adipose tissue hypoxia and cytotoxic T-cell infiltration. Int. J. Obes. (Lond) 2008, 32, 451–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kintscher, U.; Hartge, M.; Hess, K.; Foryst-Ludwig, A.; Clemenz, M.; Wabitsch, M.; Fischer-Posovszky, P.; Barth, T.F.; Dragun, D.; Skurk, T.; et al. T-lymphocyte infiltration in visceral adipose tissue: A primary event in adipose tissue inflammation and the development of obesity-mediated insulin resistance. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1304–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, S.; Manabe, I.; Nagasaki, M.; Eto, K.; Yamashita, H.; Ohsugi, M.; Otsu, M.; Hara, K.; Ueki, K.; Sugiura, S.; et al. CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med. 2009, 15, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Strissel, K.J.; DeFuria, J.; Shaul, M.E.; Bennett, G.; Greenberg, A.S.; Obin, M.S. T-cell recruitment and Th1 polarization in adipose tissue during diet-induced obesity in C57BL/6 mice. Obesity (Silver Spring) 2010, 18, 1918–1925. [Google Scholar] [CrossRef]
- Feuerer, M.; Herrero, L.; Cipolletta, D.; Naaz, A.; Wong, J.; Nayer, A.; Lee, J.; Goldfine, A.B.; Benoist, C.; Shoelson, S.; et al. Lean, but not obese, fat is enriched for a unique population of regulatory T cells that affect metabolic parameters. Nat. Med. 2009, 15, 930–939. [Google Scholar] [CrossRef]
- Lynch, L.; Nowak, M.; Varghese, B.; Clark, J.; Hogan, A.E.; Toxavidis, V.; Balk, S.P.; O’Shea, D.; O’Farrelly, C.; Exley, M.A. Adipose tissue invariant NKT cells protect against diet-induced obesity and metabolic disorder through regulatory cytokine production. Immunity 2012, 37, 574–587. [Google Scholar] [CrossRef] [Green Version]
- Lynch, L.; O’Shea, D.; Winter, D.C.; Geoghegan, J.; Doherty, D.G.; O’Farrelly, C. Invariant NKT cells and CD1d(+) cells amass in human omentum and are depleted in patients with cancer and obesity. Eur. J. Immunol. 2009, 39, 1893–1901. [Google Scholar] [CrossRef]
- Mehta, P.; Nuotio-Antar, A.M.; Smith, C.W. gammadelta T cells promote inflammation and insulin resistance during high fat diet-induced obesity in mice. J. Leukoc. Biol. 2015, 97, 121–134. [Google Scholar] [CrossRef] [Green Version]
- Le Menn, G.; Sibille, B.; Murdaca, J.; Rousseau, A.S.; Squillace, R.; Vergoni, B.; Cormont, M.; Niot, I.; Grimaldi, P.A.; Mothe-Satney, I.; et al. Decrease in alphabeta/gammadelta T-cell ratio is accompanied by a reduction in high-fat diet-induced weight gain, insulin resistance, and inflammation. FASEB J. 2019, 33, 2553–2562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, K.R.; Costanzo, A.E.; Jameson, J.M. Dysfunctional gammadelta T cells contribute to impaired keratinocyte homeostasis in mouse models of obesity. J. Investig. Dermatol. 2011, 131, 2409–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, K.R.; Mills, R.E.; Costanzo, A.E.; Jameson, J.M. Gammadelta T cells are reduced and rendered unresponsive by hyperglycemia and chronic TNFalpha in mouse models of obesity and metabolic disease. PLoS ONE 2010, 5, e11422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.; Cheung, K.P.; Limon, N.; Costanzo, A.; Barba, C.; Miranda, N.; Gargas, S.; Johnson, A.M.F.; Olefsky, J.M.; Jameson, J.M. Obesity Modulates Intestinal Intraepithelial T Cell Persistence, CD103 and CCR9 Expression, and Outcome in Dextran Sulfate Sodium-Induced Colitis. J. Immunol. 2019, 203, 3427–3435. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, E.L.; Shchukina, I.; Asher, J.L.; Sidorov, S.; Artyomov, M.N.; Dixit, V.D. Ketogenesis activates metabolically protective γδ T cells in visceral adipose tissue. Nat. Metab. 2020, 2, 50–61. [Google Scholar] [CrossRef]
- Zuniga, L.A.; Shen, W.J.; Joyce-Shaikh, B.; Pyatnova, E.A.; Richards, A.G.; Thom, C.; Andrade, S.M.; Cua, D.J.; Kraemer, F.B.; Butcher, E.C. IL-17 regulates adipogenesis, glucose homeostasis, and obesity. J. Immunol. 2010, 185, 6947–6959. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, C.; Zeng, X.; Resch, J.M.; Jedrychowski, M.P.; Yang, Z.; Desai, B.N.; Banks, A.S.; Lowell, B.B.; Mathis, D.; et al. gammadelta T cells and adipocyte IL-17RC control fat innervation and thermogenesis. Nature 2020, 578, 610–614. [Google Scholar] [CrossRef]
- Flajnik, M.F.; Kasahara, M. Origin and evolution of the adaptive immune system: Genetic events and selective pressures. Nat. Rev. Genet. 2010, 11, 47–59. [Google Scholar] [CrossRef] [Green Version]
- Litman, G.W.; Rast, J.P.; Fugmann, S.D. The origins of vertebrate adaptive immunity. Nat. Rev. Immunol. 2010, 10, 543–553. [Google Scholar] [CrossRef] [Green Version]
- Serrano, M.; Moreno, M.; Bassols, J.; Moreno-Navarrete, J.M.; Ortega, F.; Ricart, W.; Fernandez-Real, J.M. Coxsackie and adenovirus receptor is increased in adipose tissue of obese subjects: A role for adenovirus infection? J. Clin. Endocrinol. Metab. 2015, 100, 1156–1163. [Google Scholar] [CrossRef] [Green Version]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, M.D.; Witherden, D.A.; Havran, W.L. The Role of Tissue-resident γδ T Cells in Stress Surveillance and Tissue Maintenance. Cells 2020, 9, 686. https://doi.org/10.3390/cells9030686
Johnson MD, Witherden DA, Havran WL. The Role of Tissue-resident γδ T Cells in Stress Surveillance and Tissue Maintenance. Cells. 2020; 9(3):686. https://doi.org/10.3390/cells9030686
Chicago/Turabian StyleJohnson, Margarete D., Deborah A. Witherden, and Wendy L. Havran. 2020. "The Role of Tissue-resident γδ T Cells in Stress Surveillance and Tissue Maintenance" Cells 9, no. 3: 686. https://doi.org/10.3390/cells9030686