The Spindle Assembly Checkpoint Functions during Early Development in Non-Chordate Embryos


Abstract
1. Introduction
2. Materials and Methods
2.1. Gamete Collection and Fertilization
2.2. Drug Treatments
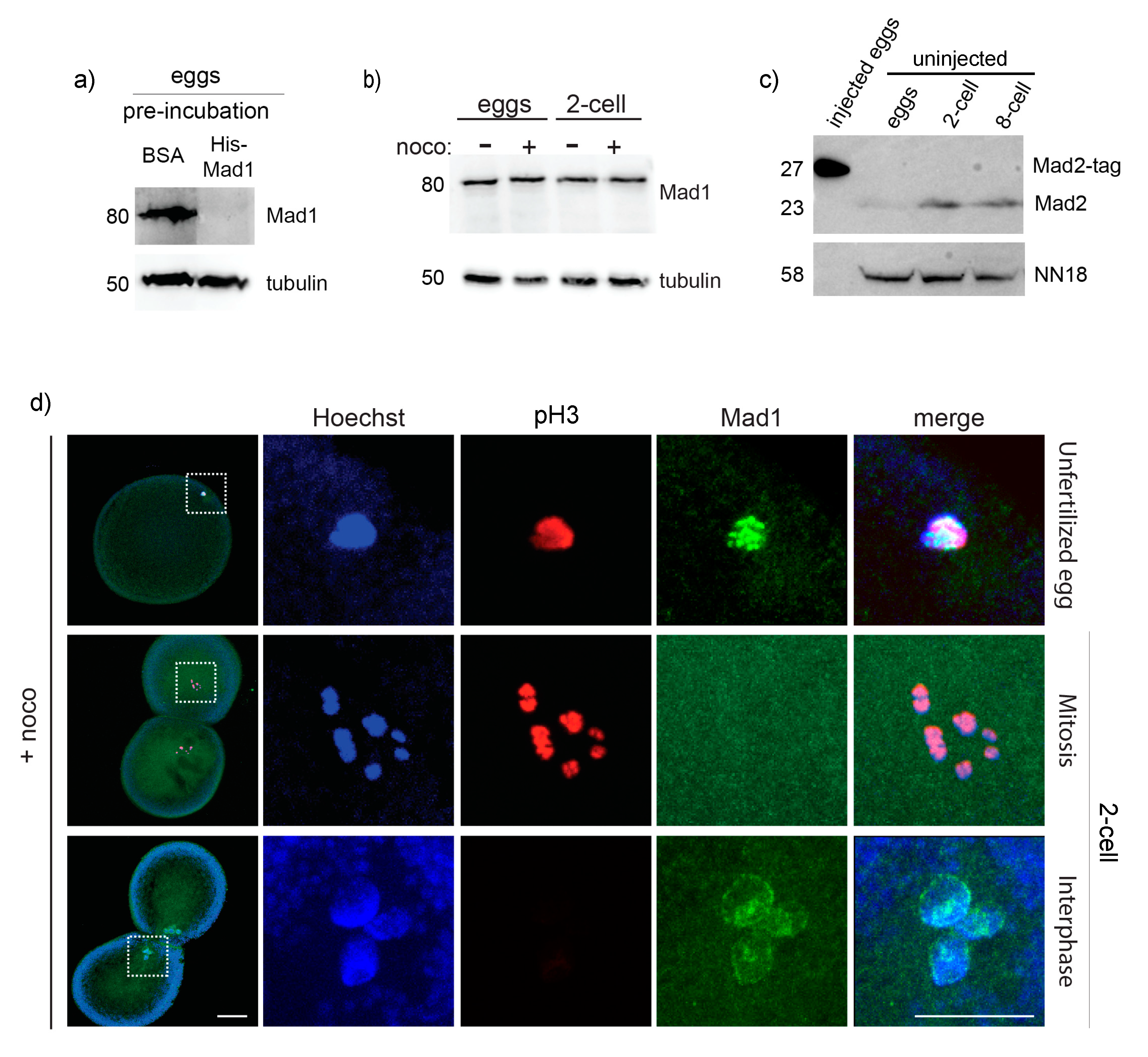
2.3. Generation of Mad1 and Mad2 Antibodies
2.4. Western Blot Analysis
2.5. Immunofluorescence
2.6. EdU (5-Ethynyl-2′-Deoxyuridine) Staining
2.7. Chromosome Spreads
2.8. Microinjection
2.9. Time-Lapse Microscopy
2.10. CellMask Staining
3. Results
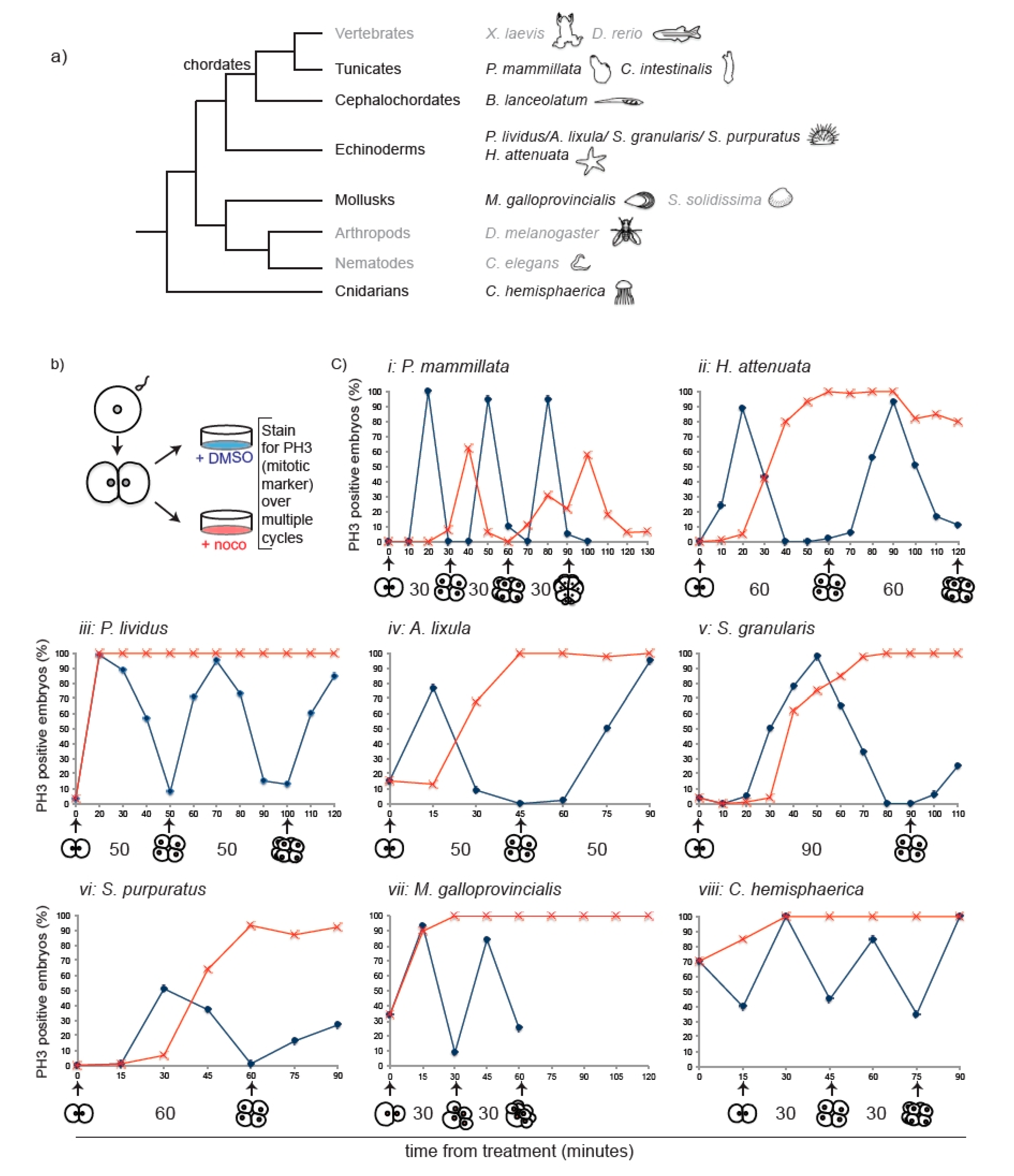
3.1. Multispecies Survey Identifies Two Classes of Embryos with Different Mitotic Responses to Spindle Defects
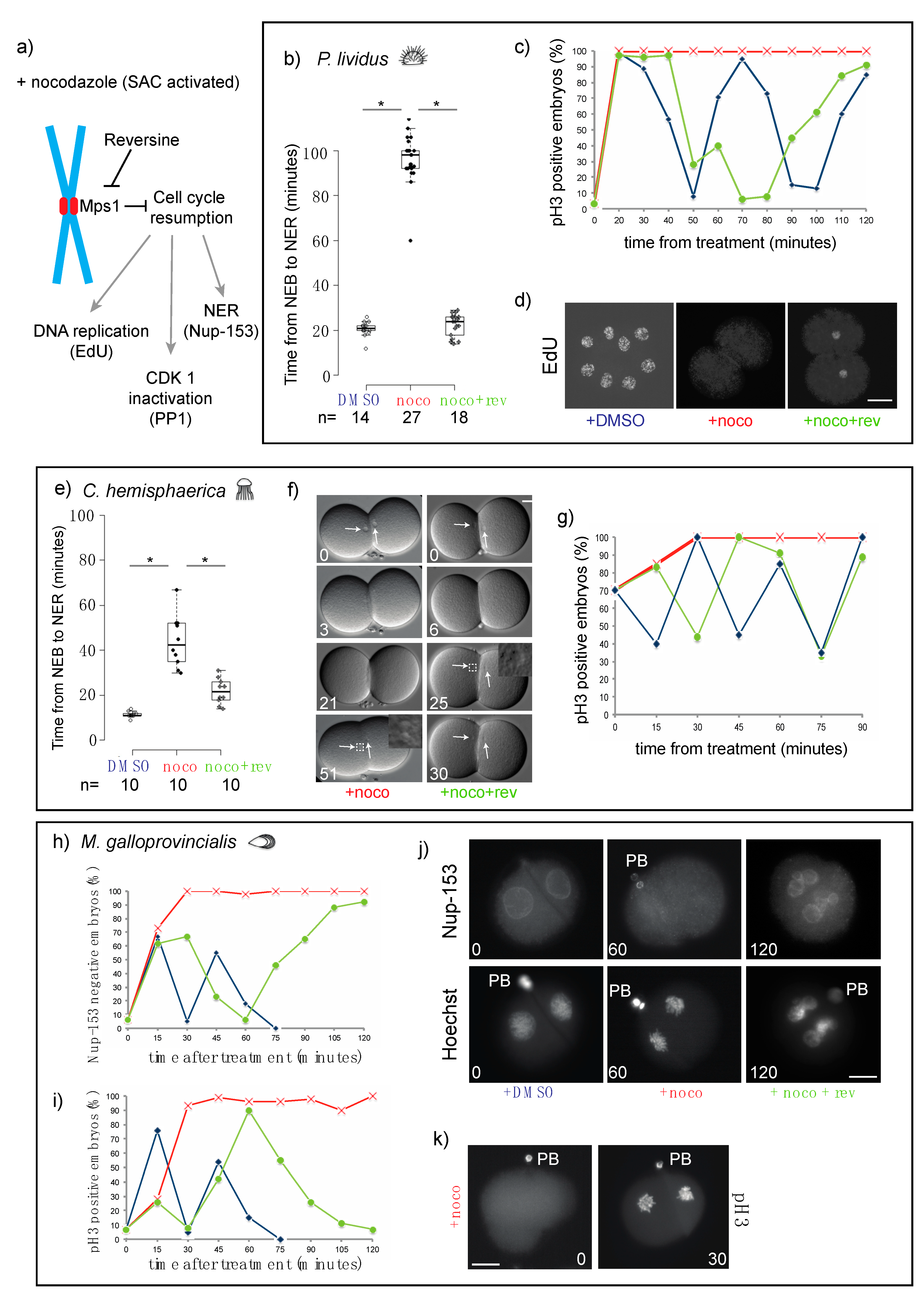
3.2. The Mitotic Delay Observed in Jellyfish, Sea Urchin, and Mussel Embryos Depends on the SAC Kinase Mps1
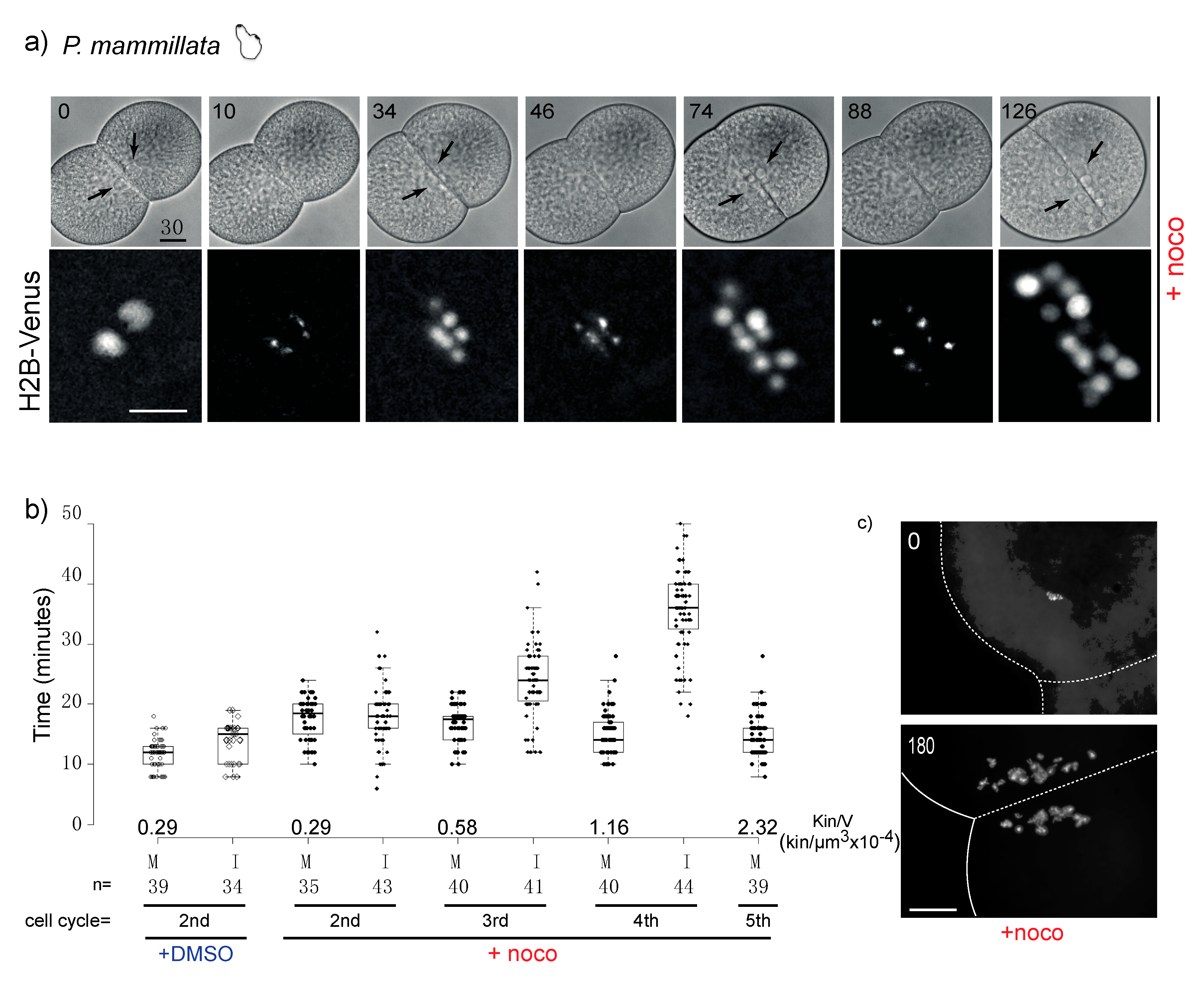
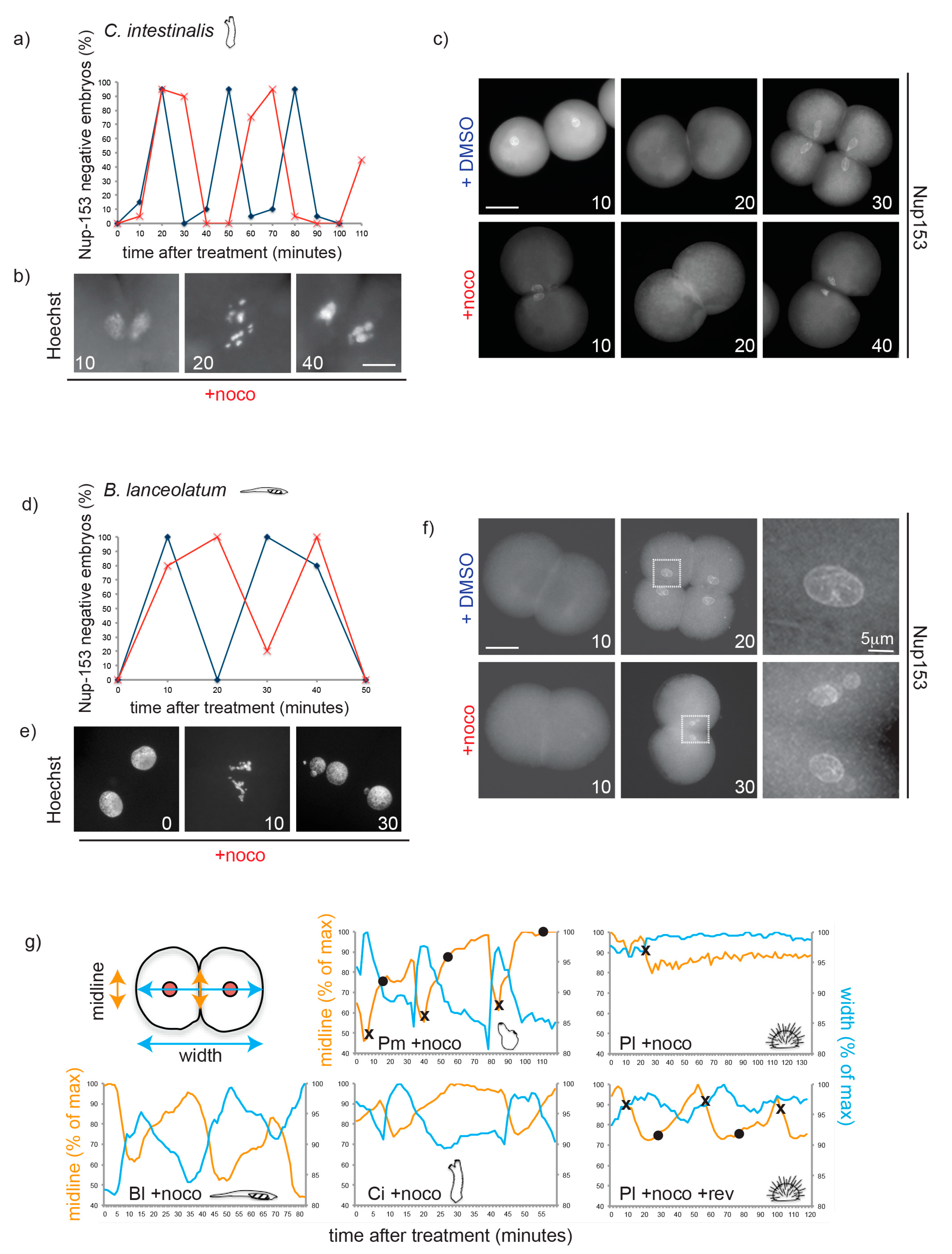
3.3. Chordate Embryos Do Not Arrest in Mitosis in the Presence of Spindle Perturbations
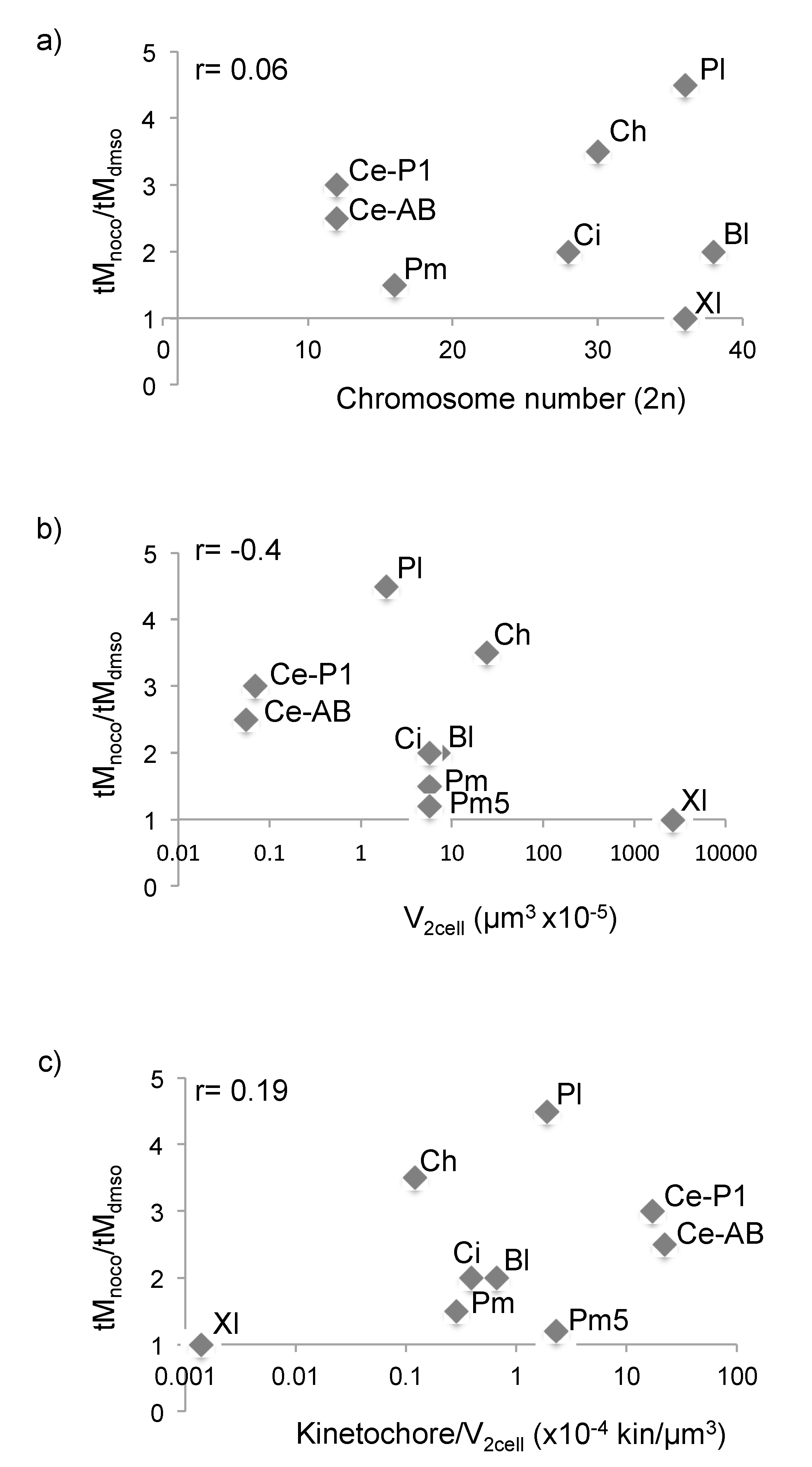
3.4. SAC Competence Does Not Correlate with Cell Size across Species
3.5. Mad1, Mad2 and Mps1 Do Not Localize to Unattached Kinetochores in P. mammillata Early Embryos
4. Discussion
4.1. SAC Activity in Embryos Defines Two Classes of Animals
4.2. SAC Activity in Relation to Kinetochore and Cytoplasm Content
4.3. SAC Deficient Embryos as an Evolutionary Novelty in the Chordate Lineage
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Musacchio, A. The molecular biology of spindle assembly checkpoint signaling dynamics. Curr. Biol. 2015, 25, R1002–R1018. [Google Scholar] [CrossRef] [PubMed]
- Serpico, A.F.; Grieco, D. Recent advances in understanding the role of CDK1 in the spindle assembly checkpoint. F1000Research 2020, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Ricke, R.M.; van Deursen, J.M. Aneuploidy in health, disease and aging. J. Cell Biol. 2013, 201, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Clute, P.; Masui, Y. Regulation of the appearance of division asynchrony and microtubule-dependent chromosome cycles in Xenopus laevis embryos. Dev. Biol. 1995, 171, 273–285. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gerhart, J.; Wu, M.; Kirschner, M. Cell cycle dynamics of an M-phase-specific cytoplasmic factor in Xenopus laevis oocytes and eggs. J. Cell Biol. 1984, 98, 1247–1255. [Google Scholar] [CrossRef]
- Ikegami, R.; Zhang, J.; Rivera-Bennetts, A.K.; Yager, T.D. Activation of the metaphase checkpoint and an apoptosis programme in the early zebrafish embryo, by treatment with the spindle-destabilizing agent nocodazole. Zygote 1997, 5, 329–350. [Google Scholar] [CrossRef]
- Zhang, M.; Kothari, P.; Lampson, M.A. Spindle assembly checkpoint acquisition at the mid-blastula transition. PLoS ONE 2015, 10, e0119285. [Google Scholar] [CrossRef]
- Kato, Y.; Tsunoda, Y. Synchronous division of mouse two-cell embryos with nocodazole in vitro. Reproduction 1992, 95, 39–43. [Google Scholar] [CrossRef]
- Vázquez-Diaz, C.; Paim, L.M.G.; FitzHarris, G. Cell-size-independent spindle checkpoint failure underlies chromosome segregation error in mouse embryos. Curr. Biol. 2019, 29, 1–9. [Google Scholar] [CrossRef]
- Hartwell, L.H.; Weinart, T.A. Checkpoint: Controls that ensure the order of cell cycle events. Science 1989, 246, 629–634. [Google Scholar] [CrossRef]
- Evans, T.; Rosenthal, E.T.; Youngblom, J.; Distel, D.; Hunt, T. Cyclin: A protein specified by maternal mRNA in sea urchin eggs that is destroyed at each cleavage division. Cell 1983, 33, 289–296. [Google Scholar] [CrossRef]
- Sluder, G.; Miller, F.J.; Thompson, E.A.; Wolf, D.E. Feedback control of the metaphase-anaphase transition in sea urchin zygotes: Role of maloriented chromosomes. J. Cell Biol. 1994, 126, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Hunt, T.; Luca, F.C.; Ruderman, J.V. The requirements for protein synthesis and degradation, and the control of destruction of cyclins A and B in the meiotic and mitotic cell cycles of the clam embryo. J. Cell Biol. 1992, 116, 707–724. [Google Scholar] [CrossRef] [PubMed]
- Shuster, C.B.; Burgess, D.R. Transitions regulating the timing of cytokinesis in embryonic cells. Curr. Biol. 2002, 12, 854–858. [Google Scholar] [CrossRef]
- Galli, M.; Morgan, D.O. Cell size determines the strength of the spindle assembly checkpoint during embryonic development. Dev. Cell 2016, 36, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Collin, P.; Nashchekina, O.; Walker, R.; Pines, J. The spindle assembly checkpoint works like a rheostat rather than a toggle switch. Nat. Cell Biol. 2013, 15, 1378–1385. [Google Scholar] [CrossRef]
- Minshull, J.; Sun, H.; Tonks, N.K.; Murray, A.W. A MAP kinase-dependent spindle assembly checkpoint in Xenopus egg extracts. Cell 1994, 79, 475–486. [Google Scholar] [CrossRef]
- Sardet, C.; McDougall, A.; Yasuo, H.; Chenevert, J.; Pruliere, G.; Dumollard, R.; Hudson, C.; Hebras, C.; Le Nguyen, N.; Paix, A. Embryological methods in ascidians: The Villefranche-sur-mer protocals. Methods Mol. Biol. 2011, 770, 365–400. [Google Scholar] [CrossRef]
- McDougall, A.; Lee, K.W.; Dumollard, R. Microinjection and 4D fluorescence imaging in the eggs and embryos of the ascidian Phallusia mammillata. Methods Mol. Biol. 2014, 1128, 175–185. [Google Scholar] [CrossRef]
- Theodosiou, M.; Colin, A.; Schulz, J.; Laudet, V.; Peyrieras, N.; Nicolas, J.-F.; Schubert, M.; Hirsinger, E. Amphioxus spawning behavior in an artificial seawater facility. J. Exp. Zool. Part B: Mol. Dev. Evol. 2011, 316, 263–275. [Google Scholar] [CrossRef]
- Houliston, E.; Momose, T.; Manuel, M. Clytia hemisphaerica: A jellyfish cousin joins the laboratory. Trends Genet. 2010, 26, 159–167. [Google Scholar] [CrossRef]
- Chenevert, J.; Pruliere, G.; Ishii, H.; Sardet, C.; Nishikata, T. Purification of mitochondrial proteins HSP60 and ATP Synthase from ascidian eggs: implications for antibody specificity. PLoS ONE 2013, 8, e52996. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Hendzel, M.J.; Wei, Y.; Mancini, M.A.; Van Hooser, A.; Ranalli, T.; Brinkley, B.R.; Bazett-Jones, D.P.; Allis, C.D. Mitosis-specific phosphorylation of histone H3 initiates primarily within pericentromeric heterochromatin during G2 and spreads in an ordered fashion coincident with mitotic chromosome condensation. Chromosoma 1997, 106, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Abrieu, A.; Magnaghi-Jaulin, L.; Kahana, J.A.; Peter, M.; Castro, A.; Vigneron, S.; Lorca, T.; Cleveland, D.W.; Labbe, J.C. Mps1 is a kinetochore-associated kinase essential for the vertebrate mitotic checkpoint. Cell 2001, 106, 83–93. [Google Scholar] [CrossRef]
- Sacristan, C.; Kops, G.J. Joined at the hip: Kinetochores, microtubules and spindle assembly checkpoint signaling. Trends Cell Biol. 2015, 25, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Santaguida, S.; Tighe, A.; D’Alise, A.M.; Taylor, S.S.; Musacchio, A. Dissecting the role of Mps1 in chromosome biorientation and the spindle checkpoint through the small molecule inhibitor reversine. J. Cell Biol. 2010, 190, 73–87. [Google Scholar] [CrossRef]
- Hewitt, L.; Tighe, A.; Santaguida, S.; White, A.M.; Jones, C.D.; Musacchio, A.; Green, S.; Taylor, S.S. Sustained Mps1 activity is required in mitosis to recruit O-Mad2 to the Mad1-C-Mad2 core complex. J. Cell Biol. 2010, 190, 25–34. [Google Scholar] [CrossRef]
- Lewis, C.W.; Taylor, R.G.; Kubara, P.M.; Marshall, K.; Meijer, L.; Golsteyn, R.M. A western blot assay to measure cyclin dependent kinase activity in cells or in vitro without the use of radioisotopes. FEBS Lett. 2013, 587, 3089–3095. [Google Scholar] [CrossRef]
- Wu, J.Q.; Guo, J.Y.; Tang, W.; Yang, C.S.; Freel, C.D.; Chen, C.; Nairn, A.C.; Kornblut, S. PP1-mediated dephosphorylation of phosphoproteins at mitotic exit is controlled by inhibitor-1 and PP1 phosphorylation. Nat. Cell Biol. 2009, 11, 644–651. [Google Scholar] [CrossRef]
- Rieder, C.L.; Cole, R. Microtubule disassembly delays the G2-M transition in vertebrates. Curr. Biol. 2000, 10, 1067–1070. [Google Scholar] [CrossRef]
- Lancaster, O.M.; Le Berre, M.; Dimitracopoulus, A.; Bonazzi, D.; Zlotek-Zlotekiewicz, E.; Picone, R.; Duke, T.; Piel, M.; Baum, B. Mitotic rounding alters cell geometry to ensure efficient bipolar spindle formation. Dev. Cell. 2013, 25, 270–283. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.H.; Heald, R. Nuclear size is regulated by Importin α and Ntf2 in Xenopus. Cell 2010, 143, 288–298. [Google Scholar] [CrossRef]
- Wühr, M.; Chen, Y.; Dumont, S.; Groen, A.C.; Needleman, D.J.; Salic, A.; Mitchison, T.J. Evidence for an upper limit to mitotic spindle length. Curr. Biol. 2008, 18, 1256–1261. [Google Scholar] [CrossRef] [PubMed]
- Colomber, D. Male chromosomes in two populations of Branchiostoma lanceolatum. Experientia 1974, 30, 353–355. [Google Scholar] [CrossRef] [PubMed]
- Duffy, L.; Sewell, M.A.; Murray, B.G. Chromosome number and chromosome variation in embryos of Evechinus chloroticus (Echinoidea: Echinometridae): Is there conservation of chromosome number in the phylum Echinodermata? New findings and a brief review. Invertebr. Reprod. Dev. 2007, 50, 219–231. [Google Scholar] [CrossRef]
- Lipani, C.; Vitturi, R.; Sconzo, G.; Barbata, G. Karyotype analysis of the sea urchin Paracentrotus lividus (Echinodermata): Evidence for a heteromorphic chromosome sex mechanism. Marine Biol. 1996, 127, 67–72. [Google Scholar] [CrossRef]
- Lacroix, B.; Letort, G.; Pitayu, L.; Sallé, J.; Stefanutti, M.; Maton, G.; Ladouceur, A.-M.; Canman, J.C.; Maddox, P.S.; Maddox, A.S. Microtubule dynamics scale with cell size to set spindle length and assembly timing. Dev. Cell 2018, 45, 496–511. [Google Scholar] [CrossRef]
- Eno, C.C.; Boettger, S.A.; Walker, C.W. Methods for karyotyping and for localization of developmentally relevant genes on the chromosomes of the purple sea urchin Strongylocentrotus purpuratus. Biol. Bull. 2009, 217, 306–312. [Google Scholar] [CrossRef]
- Dixon, D.R.; Flavell, N. A comparative study of the chromosomes of Mytilus edulis and Mytilus galloprovincialis. J. Mar. Biol. Assoc. U.K. 1986, 66, 219–228. [Google Scholar] [CrossRef]
- Perez-Mongiovi, D.; Malmanche, N.; Bousbaa, H.; Sunkel, C. Maternal expression of the checkpoint protein BubR1 is required for synchrony of syncytial nuclear division and polar body arrest in Drosophila melanogaster. Development 2005, 132, 4509–4520. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ladouceur, A.M.; Dorn, J.F.; Maddox, P.S. Mitotic chromosome length scales in response to both cell and nuclear size. J. Cell Biol. 2015, 209, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Greenan, G.; Brangwynne, C.P.; Jaesch, S.; Gharakhani, J.; Jülicher, F.; Hyman, A.A. Centrosome size sets mitotic spindle length in Caenorhabditis elegans embryos. Curr. Biol. 2010, 20, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Encalada, S.E.; Willis, J.; Lyczak, R.; Bowerman, B. A spindle checkpoint functions during mitosis in the early Caenorhabditis elegans embryo. Mol. Biol. Cell 2005, 16, 1056–1070. [Google Scholar] [CrossRef] [PubMed]
- Crowder, M.E.; Strelecka, M.; Wilbur, J.D.; Good, M.C.; von Dassow, G.; Heald, R. Comparative analysis of spindle morphometrics across metazoans. Curr. Biol. 2015, 25, 1542–1550. [Google Scholar] [CrossRef] [PubMed]
- Brozovic, M.; Dantec, C.; Dardaillon, J.; Dauga, D.; Faure, E.; Gineste, M.; Louis, A.; Naville, M.; Nitta, K.R.; Piette, J.; et al. ANISEED 2017: Extending the integrated ascidian database to the exploration and evolutionary comparison of genome-scale datasets. Nucleic Acids Res. 2018, 46, D718–D725. [Google Scholar] [CrossRef]
- Marlétaz, F.; Firbas, P.N.; Irimia, M. Amphioxus functional genomics and the origins of vertebrate gene regulation. Nature 2018, 564, 64–70. [Google Scholar] [CrossRef]
- Oulion, S.; Bertrand, S.; Belgacem, M.R.; Le Petillion, Y.; Escriva, H. Sequencing and analysis of the Mediterranean amphioxus (Branchiostoma lanceolatum) transcriptome. PLoS ONE 2012, 7, e36554. [Google Scholar] [CrossRef]
- Chen, R.H.; Shevchenko, A.; Mann, M.; Murray, A.W. Spindle checkpoint protein Xmad1 recruits Xmad2 to unattached kinetochores. J. Cell Biol. 1998, 143, 283–295. [Google Scholar] [CrossRef]
- Campbell, M.S.; Chan, G.K.; Yen, T.J. Mitotic checkpoint proteins HsMad1 and HsMad2 are associated with nuclear pore complexes in interphase. J. Cell Sci. 2001, 8, 2607–2618. [Google Scholar]
- Iouk, T.; Kerscher, O.; Scott, R.J.; Basrai, M.A.; Wozniak, R.W. The yeast nuclear pore complex functionally interacts with components of the spindle assembly checkpoint. J. Cell Biol. 2002, 159, 807–819. [Google Scholar] [CrossRef]
- Musacchio, A.; Salmon, E.D. The spindle checkpoint in time and space. Nat. Rev. Mol. Cell Biol. 2007, 8, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Kyogoku, H.; Kitajima, T.S. Large cytoplasm is linked to the error-prone nature of oocytes. Dev. Cell 2017, 41, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, W.; Daily, D.R.; Fogarty, P.; Yook, K.J.; Pimpinelli, S. Delays in anaphase initiation occur in individual nuclei of the syncytial Drosophila embryo. Mol. Biol. Cell 1993, 4, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Su, T.T.; Sprenger, F.; Digregorio, P.J.; Campbell, S.D.; O’Farrell, P.H. Exit from mitosis in Drosophila syncytial embryos requires proteolysis and cyclin degradation, and is associated with localized dephosphorylation. Genes Dev. 1998, 12, 1495–1503. [Google Scholar] [CrossRef]
- Rieder, C.L.; Khodjakov, A.; Paliulis, L.V.; Fortier, T.M.; Cole, R.W.; Sluder, G. Mitosis in vertebrate cells with two spindles: Implications for the metaphase/anaphase transition checkpoint and cleavage. Proc. Natl. Acad. Sci. USA 1997, 94, 5107–5112. [Google Scholar] [CrossRef]
- Gerhold, A.R.; Poupart, V.; Labbé, J.C.; Maddox, P.S. Spindle assembly checkpoint strength is linked to cell fate in the C. elegans embryo. Mol. Biol. Cell 2018, 29, 1435–1443. [Google Scholar] [CrossRef]
- Cather, J.N.; Render, J.A.; Freeman, G. The relation of time to direction and equality of cleavage in Ilyanassa embryos. Int. J. Invertebr. Reprod. Dev. 1985, 9, 179–194. [Google Scholar] [CrossRef]
- Wolf, R. The cytaster, a colchicine-sensitive migration organelle of cleavage nuclei in the insect egg. Dev. Biol. 1978, 62, 464–472. [Google Scholar] [CrossRef]
- Kaiser, J.; Went, D.F. Early embryonic development of the dipteran insect Heteropeza pygmaea in the presence of cytoskeleton drugs. Roux’s Arch. Dev. Biol. 1987, 196, 356–366. [Google Scholar] [CrossRef]
- Nelson, C.R.; Hwang, T.; Chen, P.-H.; Bhalla, N. TRIP13PCH−2 promotes Mad2 localization to unattached kinetochores in the spindle checkpoint response. J. Cell Biol. 2015, 211, 503–516. [Google Scholar] [CrossRef] [PubMed]
- Dou, Z.; Prifti, D.K.; Gui, P.; Liu, X.; Elowe, S.; Yao, X. Recent progress on the localization of the spindle assembly checkpoint machinery to kinetochores. Cells 2019, 8, 278. [Google Scholar] [CrossRef] [PubMed]
- Dumollard, R.; Levasseur, M.; Hebras, C.; Huitorel, P.; Carroll, M.; Chambon, J.-P.; McDougall, A. Mos limits the number of meiotic divisions in urochordate eggs. Development 2011, 138, 885–895. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cheng, X.; Zhang, Y.; Li, S.; Cui, H.; Zhang, L.; Shi, R.; Zhao, Z.; He, C.; Wang, C.; et al. Phosphorylation of Mps1 by BRAFV600E prevents Mps1 degradation and contributes to chromosome instability in melanoma. Oncogene 2013, 32, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, R.-H. Mps1 phosphorylation by MAP kinase is required for kinetochore localization of spindle checkpoint proteins. Curr. Biol. 2006, 16, 1764–1769. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Oocyte Diameter (µm) | Chromosome Number (2n) | Kin/V2cell × 10−4 (kin/µm3) | Vnucleus (µm3) | Spindle Length (µm) | SAC Competence Reference | |
|---|---|---|---|---|---|---|---|
| Chordates | Xenopus laevis | 1000 | 36 | 0.0014 | 3053 [33] | 53.5 [34] | − [5] |
| Danio rerio | 800 | 50 | 0.0037 | − [6] | |||
| Ciona intestinalis | 140 | 28 | 0.39 | − this study | |||
| Phallusia mammillata | 130 | 16 | 0.29 | 1345 | 20–30 | − this study | |
| Branchiostoma lanceolatum | 130 | 38 [35] | 0.66 | − this study | |||
| Echinoderms | Hacelia attenuata | 155 | 44 [36] | 0.45 | + this study | ||
| Paracentrotus lividus | 90 | 36 [37] | 1.9 | 1562 | 18 [38] | + this study | |
| Arbacia lixula | 80 | 44 [36] | 3.3 | + this study | |||
| Strongylocentrotus purpuratus | 80 | 42 [39] | 3.1 | + this study | |||
| Sphaerechinus granularis | 110 | 42 [36] | 1.2 | + this study | |||
| Mollusks | Mytilus galloprovincialis | 65 | 28 [40] | Small = 6.6 Large = 2.5 | 2393 | + this study | |
| Spisula solidissima | 60 | 28 | 6.7 | + [11] | |||
| Arthropods | Drosophila melanogaster | 500 | 8 | 0.018 | + [41] | ||
| Nematodes | Caenorhabditis elegans | 50 | 12 | AB = 22 [15] P1 = 17 [15] | 523 [42] | 13 [43] | + [44] |
| Cnidarians | Clytia hemisphaerica | 210 | 30 | 0.12 | 30–35 | + this study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chenevert, J.; Roca, M.; Besnardeau, L.; Ruggiero, A.; Nabi, D.; McDougall, A.; Copley, R.R.; Christians, E.; Castagnetti, S. The Spindle Assembly Checkpoint Functions during Early Development in Non-Chordate Embryos. Cells 2020, 9, 1087. https://doi.org/10.3390/cells9051087
Chenevert J, Roca M, Besnardeau L, Ruggiero A, Nabi D, McDougall A, Copley RR, Christians E, Castagnetti S. The Spindle Assembly Checkpoint Functions during Early Development in Non-Chordate Embryos. Cells. 2020; 9(5):1087. https://doi.org/10.3390/cells9051087
Chicago/Turabian StyleChenevert, Janet, Marianne Roca, Lydia Besnardeau, Antonella Ruggiero, Dalileh Nabi, Alex McDougall, Richard R. Copley, Elisabeth Christians, and Stefania Castagnetti. 2020. "The Spindle Assembly Checkpoint Functions during Early Development in Non-Chordate Embryos" Cells 9, no. 5: 1087. https://doi.org/10.3390/cells9051087
APA StyleChenevert, J., Roca, M., Besnardeau, L., Ruggiero, A., Nabi, D., McDougall, A., Copley, R. R., Christians, E., & Castagnetti, S. (2020). The Spindle Assembly Checkpoint Functions during Early Development in Non-Chordate Embryos. Cells, 9(5), 1087. https://doi.org/10.3390/cells9051087