A Multiparametric Assay Platform for Simultaneous In Vivo Assessment of Pronephric Morphology, Renal Function and Heart Rate in Larval Zebrafish

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Fish Keeping and Embryo Handling
2.3. Drug Treatment of Embryos
2.4. Generation of a 96-Well Dorsal-Lateral Template Tool
2.5. Preparation of Agarose Molds in Microtiter Plates
2.6. Fluorescein Isothiocyanate (FITC)-Inulin Injection
2.7. Image Acquisition
2.8. Data Handling and Visualization
2.9. Image Analysis
2.10. Statistical Analysis
3. Results
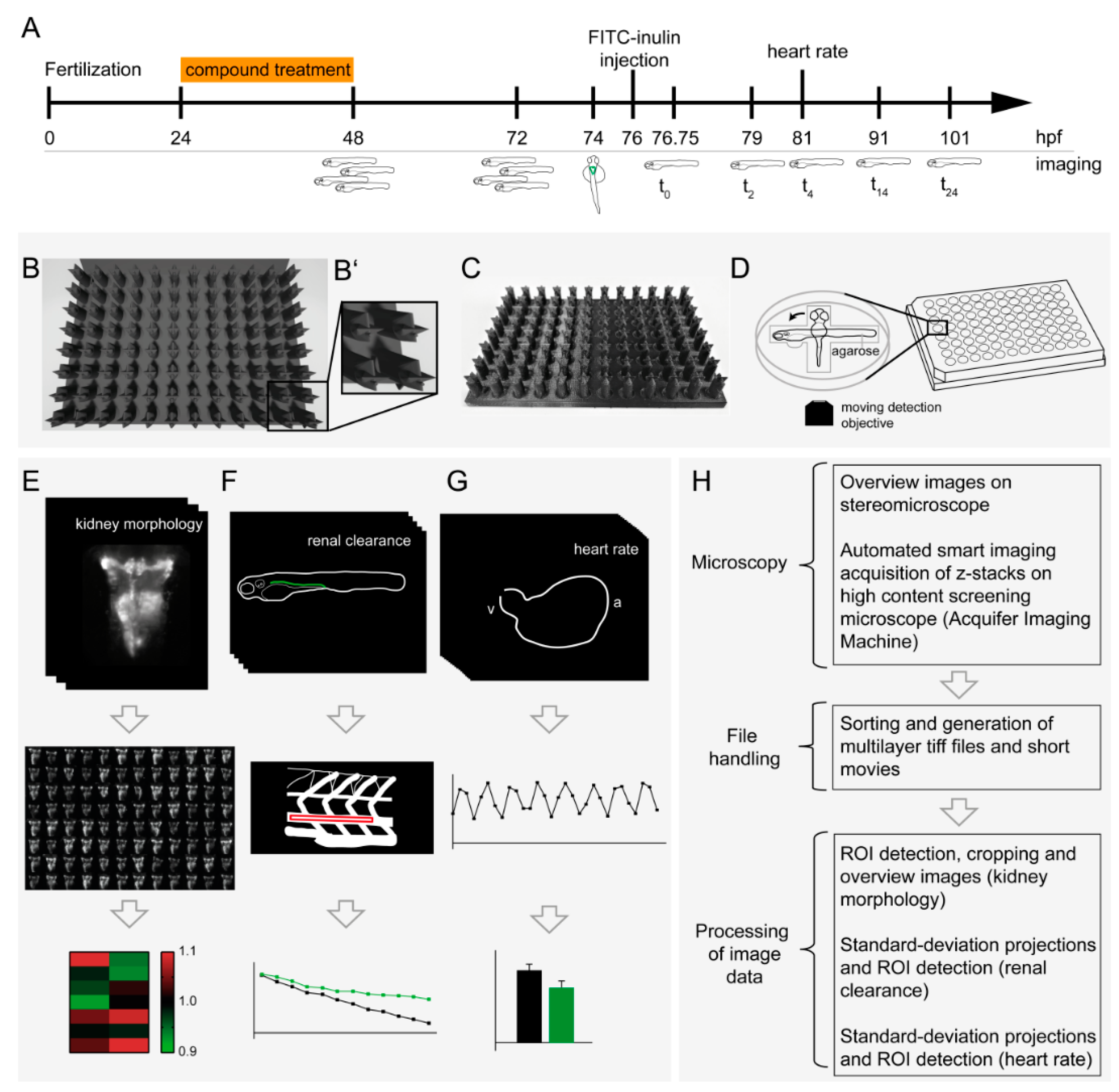
3.1. Multiparametric In Vivo Pipeline for the Assessment of Renal Morphology, Renal Clearance and Heart Rate
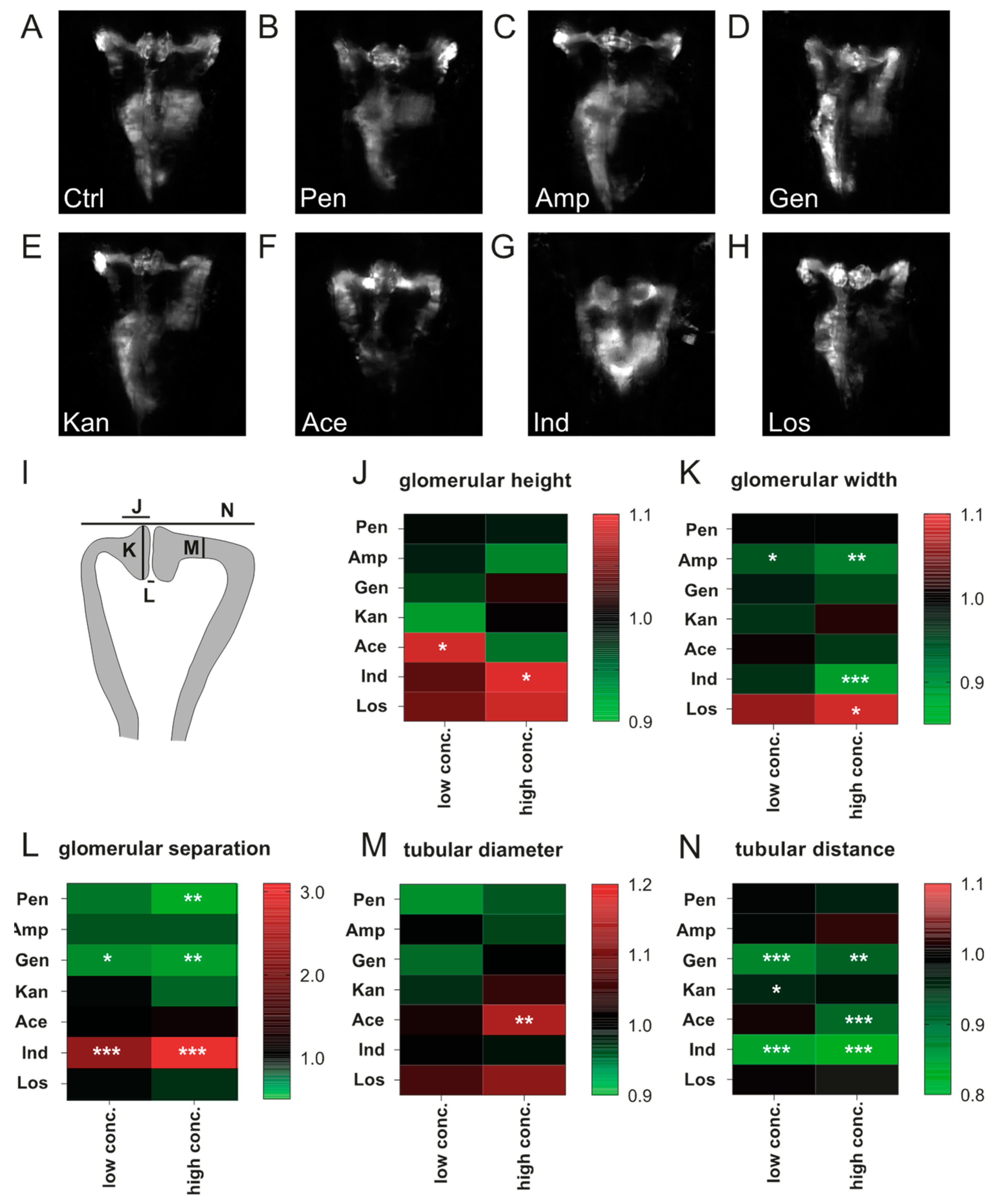
3.2. Impact of Compound Treatment on Pronephros Development
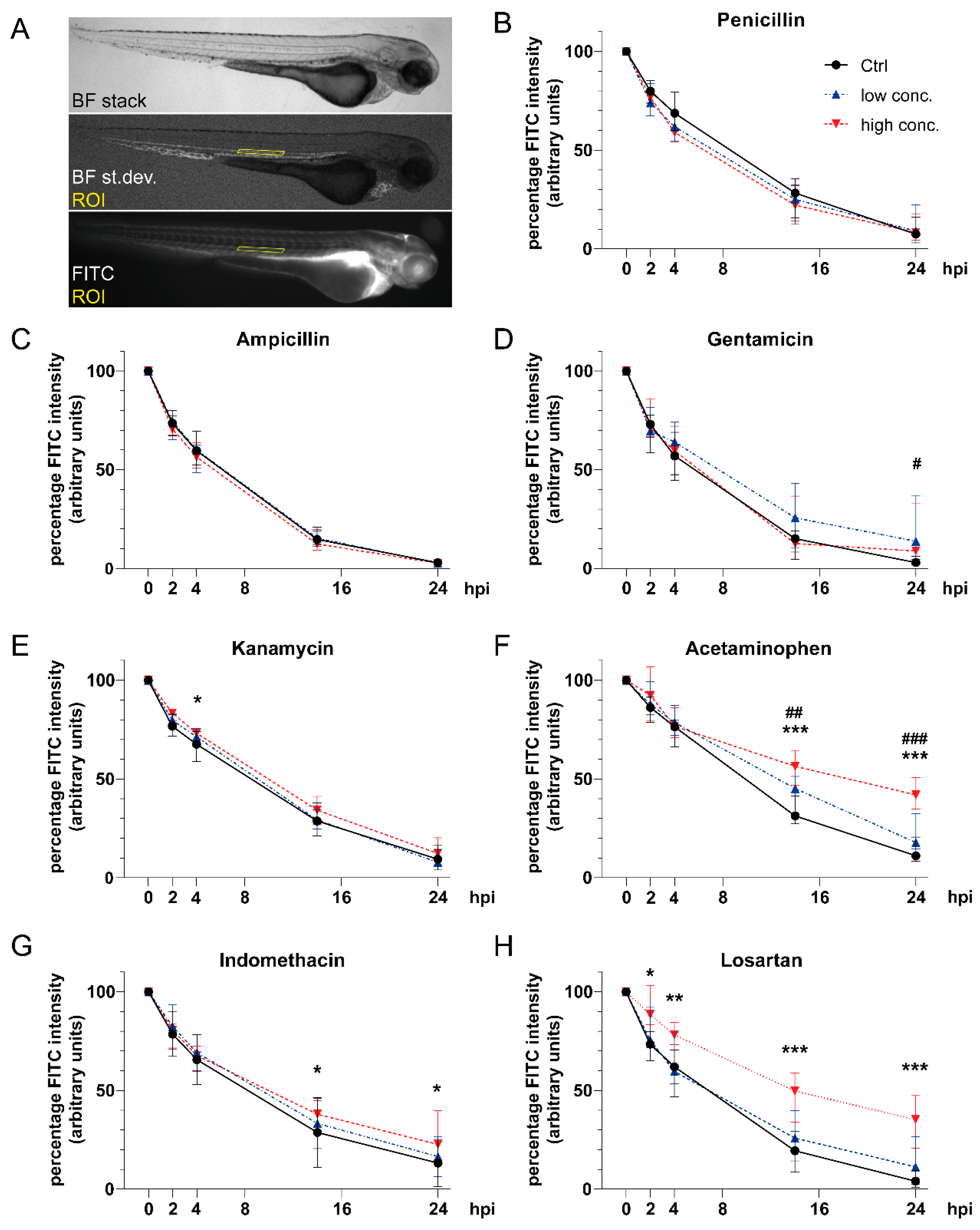
3.3. Impact of Tested Compounds on Pronephric Clearance
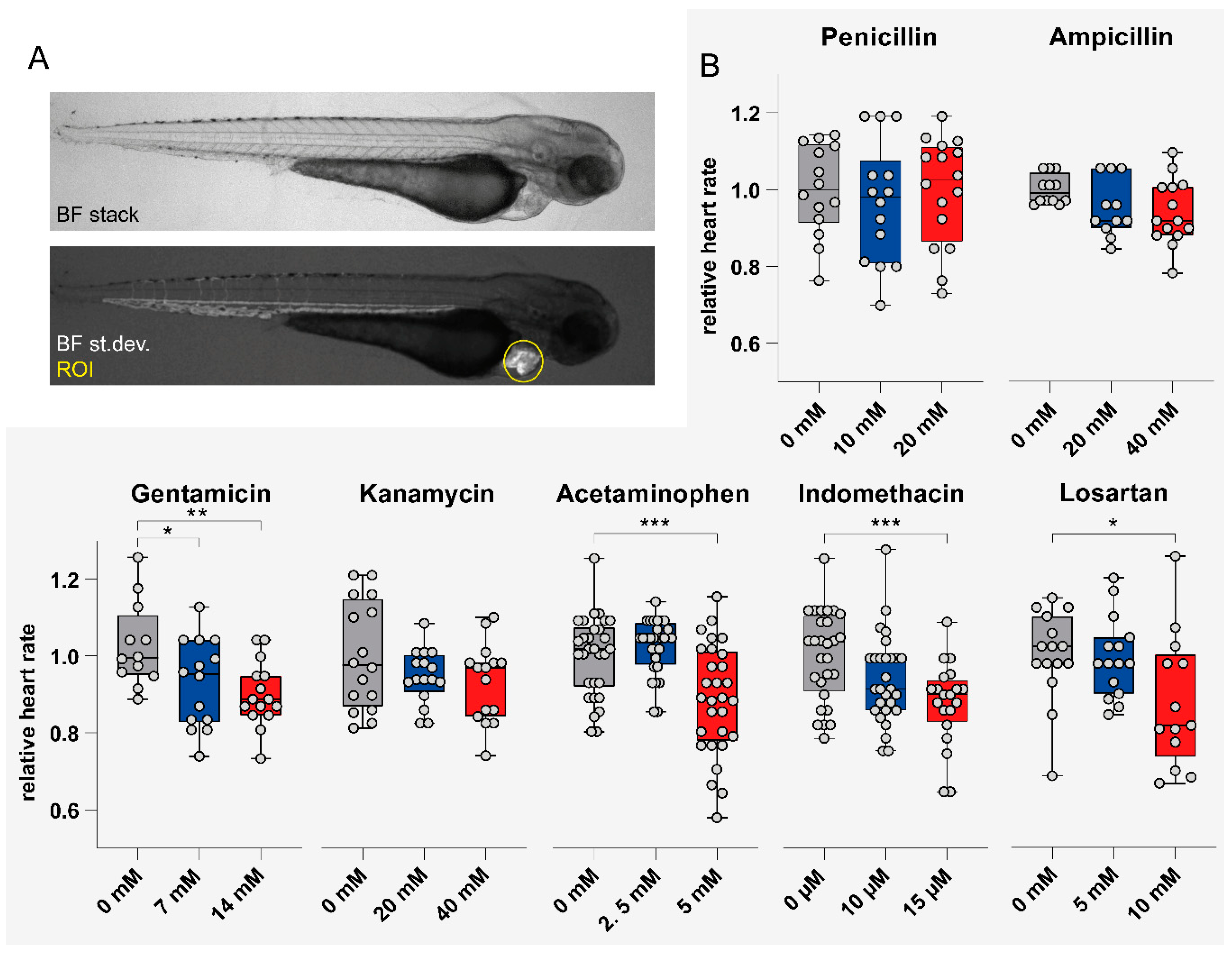
3.4. Impact of Tested Compounds on Larval Heart Rate
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rovida, C.; Hartung, T. Re-evaluation of animal numbers and costs for in vivo tests to accomplish REACH legislation requirements for chemicals - a report by the transatlantic think tank for toxicology (t(4)). ALTEX 2009, 26, 187–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, E.S.; Roberts, R.A. Collaboration and competition: Ethics in toxicology. Toxicol Res. (Camb) 2018, 7, 576–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacArthur Clark, J. The 3Rs in research: A contemporary approach to replacement, reduction and refinement. Br. J. Nutr. 2018, 120, S1–S7. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.M. The development of the three Rs concept. Altern Lab. Anim 1995, 23, 298–304. [Google Scholar] [PubMed]
- Cassar, S.; Adatto, I.; Freeman, J.L.; Gamse, J.T.; Iturria, I.; Lawrence, C.; Muriana, A.; Peterson, R.T.; Van Cruchten, S.; Zon, L.I. Use of Zebrafish in Drug Discovery Toxicology. Chem. Res. Toxicol. 2019, 33, 95–118. [Google Scholar] [CrossRef] [Green Version]
- Gehrig, J.; Pandey, G.; Westhoff, J.H. Zebrafish as a Model for Drug Screening in Genetic Kidney Diseases. Front. Pediatr. 2018, 6, 183. [Google Scholar] [CrossRef]
- MacRae, C.A.; Peterson, R.T. Zebrafish as tools for drug discovery. Nat. Rev. Drug Discov. 2015, 14, 721–731. [Google Scholar] [CrossRef]
- Horzmann, K.A.; Freeman, J.L. Making Waves: New Developments in Toxicology With the Zebrafish. Toxicol Sci 2018, 163, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Drummond, I.A.; Davidson, A.J. Zebrafish kidney development. Methods Cell Biol. 2010, 100, 233–260. [Google Scholar]
- Wingert, R.A.; Selleck, R.; Yu, J.; Song, H.D.; Chen, Z.; Song, A.; Zhou, Y.; Thisse, B.; Thisse, C.; McMahon, A.P.; et al. The cdx genes and retinoic acid control the positioning and segmentation of the zebrafish pronephros. PLoS Genet. 2007, 3, 1922–1938. [Google Scholar] [CrossRef] [Green Version]
- Kramer-Zucker, A.G.; Olale, F.; Haycraft, C.J.; Yoder, B.K.; Schier, A.F.; Drummond, I.A. Cilia-driven fluid flow in the zebrafish pronephros, brain and Kupffer’s vesicle is required for normal organogenesis. Development 2005, 132, 1907–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, E.E.; Wingert, R.A. Zebrafish as a Model of Kidney Disease. Results Probl Cell Differ. 2017, 60, 55–75. [Google Scholar] [PubMed]
- Outtandy, P.; Russell, C.; Kleta, R.; Bockenhauer, D. Zebrafish as a model for kidney function and disease. Pediatr. Nephrol. 2019, 34, 751–762. [Google Scholar] [CrossRef] [Green Version]
- Pandey, G.; Westhoff, J.H.; Schaefer, F.; Gehrig, J. A Smart Imaging Workflow for Organ-Specific Screening in a Cystic Kidney Zebrafish Disease Model. Int. J. Mol. Sci. 2019, 20, 1290. [Google Scholar] [CrossRef] [Green Version]
- Westhoff, J.H.; Giselbrecht, S.; Schmidts, M.; Schindler, S.; Beales, P.L.; Tonshoff, B.; Liebel, U.; Gehrig, J. Development of an automated imaging pipeline for the analysis of the zebrafish larval kidney. PLoS ONE 2013, 8, e82137. [Google Scholar] [CrossRef] [Green Version]
- Hanke, N.; King, B.L.; Vaske, B.; Haller, H.; Schiffer, M. A Fluorescence-Based Assay for Proteinuria Screening in Larval Zebrafish (Danio rerio). Zebrafish 2015, 12, 372–376. [Google Scholar] [CrossRef] [Green Version]
- Hentschel, D.M.; Mengel, M.; Boehme, L.; Liebsch, F.; Albertin, C.; Bonventre, J.V.; Haller, H.; Schiffer, M. Rapid screening of glomerular slit diaphragm integrity in larval zebrafish. Am. J. Physiol. Ren. Physiol. 2007, 293, F1746–F1750. [Google Scholar] [CrossRef]
- Zhou, W.; Hildebrandt, F. Inducible Podocyte Injury and Proteinuria in Transgenic Zebrafish. J. Am. Soc. Nephrol. JASN 2012, 23, 1039–1047. [Google Scholar] [CrossRef]
- Hentschel, D.M.; Park, K.M.; Cilenti, L.; Zervos, A.S.; Drummond, I.; Bonventre, J.V. Acute renal failure in zebrafish: A novel system to study a complex disease. Am. J. Physiol. Ren. Physiol. 2005, 288, F923–F929. [Google Scholar] [CrossRef] [Green Version]
- Rider, S.A.; Tucker, C.S.; del-Pozo, J.; Rose, K.N.; MacRae, C.A.; Bailey, M.A.; Mullins, J.J. Techniques for the in vivo assessment of cardio-renal function in zebrafish (Danio rerio) larvae. J. Physiol. 2012, 590, 1803–1809. [Google Scholar] [CrossRef] [Green Version]
- Kotb, A.M.; Muller, T.; Xie, J.; Anand-Apte, B.; Endlich, K.; Endlich, N. Simultaneous assessment of glomerular filtration and barrier function in live zebrafish. Am. J. Physiol. Ren. Physiol. 2014, 307, F1427–F1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorgulho, R.; Jacinto, R.; Lopes, S.S.; Pereira, S.A.; Tranfield, E.M.; Martins, G.G.; Gualda, E.J.; Derks, R.J.E.; Correia, A.C.; Steenvoorden, E.; et al. Usefulness of zebrafish larvae to evaluate drug-induced functional and morphological renal tubular alterations. Arch. Toxicol. 2018, 92, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Perner, B.; Englert, C.; Bollig, F. The Wilms tumor genes wt1a and wt1b control different steps during formation of the zebrafish pronephros. Dev. Biol. 2007, 309, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Wittbrodt, J.N.; Liebel, U.; Gehrig, J. Generation of orientation tools for automated zebrafish screening assays using desktop 3D printing. BMC Biotechnol. 2014, 14, 36. [Google Scholar] [CrossRef] [Green Version]
- Gierten, J.; Pylatiuk, C.; Hammouda, O.T.; Schock, C.; Stegmaier, J.; Wittbrodt, J.; Gehrig, J.; Loosli, F. Automated high-throughput heartbeat quantification in medaka and zebrafish embryos under physiological conditions. Sci. Rep. 2020, 10, 2046. [Google Scholar] [CrossRef]
- Westhoff, J.H.; Steenbergen, P.J.; Thomas, L.S.V.; Heigwer, J.; Bruckner, T.; Cooper, L.; Tönshoff, B.; Hoffmann, G.F.; Gehrig, J. In vivo high-content screening in zebrafish for developmental nephrotoxicity of approved drugs. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Brady, C.A.; Rennekamp, A.J.; Peterson, R.T. Chemical Screening in Zebrafish. Methods Mol. Biol. 2016, 1451, 3–16. [Google Scholar]
- Sukardi, H.; Chng, H.T.; Chan, E.C.; Gong, Z.; Lam, S.H. Zebrafish for drug toxicity screening: bridging the in vitro cell-based models and in vivo mammalian models. Expert Opin. Drug Metab. Toxicol. 2011, 7, 579–589. [Google Scholar] [CrossRef]
- Cianciolo Cosentino, C.; Roman, B.L.; Drummond, I.A.; Hukriede, N.A. Intravenous microinjections of zebrafish larvae to study acute kidney injury. J. Vis. Exp. JoVE 2010. [Google Scholar] [CrossRef]
- Burns, C.G.; Milan, D.J.; Grande, E.J.; Rottbauer, W.; MacRae, C.A.; Fishman, M.C. High-throughput assay for small molecules that modulate zebrafish embryonic heart rate. Nat. Chem. Biol. 2005, 1, 263–264. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.K.; Lin, C.C.; Cheng, S.H. Noninvasive technique for measurement of heartbeat regularity in zebrafish (Danio rerio) embryos. BMC Biotechnol. 2009, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink, M.; Callol-Massot, C.; Chu, A.; Ruiz-Lozano, P.; Izpisua Belmonte, J.C.; Giles, W.; Bodmer, R.; Ocorr, K. A new method for detection and quantification of heartbeat parameters in Drosophila, zebrafish, and embryonic mouse hearts. Biotechniques 2009, 46, 101–113. [Google Scholar] [CrossRef]
- Kang, C.P.; Tu, H.C.; Fu, T.F.; Wu, J.M.; Chu, P.H.; Chang, D.T. An automatic method to calculate heart rate from zebrafish larval cardiac videos. BMC Bioinform. 2018, 19, 169. [Google Scholar] [CrossRef] [Green Version]
- Nepstad, R.; Davies, E.; Altin, D.; Nordtug, T.; Hansen, B.H. Automatic determination of heart rates from microscopy videos of early life stages of fish. J. Toxicol. Environ. Health A 2017, 80, 932–940. [Google Scholar] [CrossRef]
- Puybareau, E.; Genest, D.; Barbeau, E.; Leonard, M.; Talbot, H. An automated assay for the assessment of cardiac arrest in fish embryo. Comput. Biol. Med. 2017, 81, 32–44. [Google Scholar] [CrossRef] [Green Version]
- Pylatiuk, C.; Sanchez, D.; Mikut, R.; Alshut, R.; Reischl, M.; Hirth, S.; Rottbauer, W.; Just, S. Automatic zebrafish heartbeat detection and analysis for zebrafish embryos. Zebrafish 2014, 11, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Yozzo, K.L.; Isales, G.M.; Raftery, T.D.; Volz, D.C. High-content screening assay for identification of chemicals impacting cardiovascular function in zebrafish embryos. Environ. Sci. Technol. 2013, 47, 11302–11310. [Google Scholar] [CrossRef]
- Kent, A.L.; Maxwell, L.E.; Koina, M.E.; Falk, M.C.; Willenborg, D.; Dahlstrom, J.E. Renal glomeruli and tubular injury following indomethacin, ibuprofen, and gentamicin exposure in a neonatal rat model. Pediatr. Res. 2007, 62, 307–312. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Salgado, C.; Lopez-Hernandez, F.J.; Lopez-Novoa, J.M. Glomerular nephrotoxicity of aminoglycosides. Toxicol. Appl. Pharmacol. 2007, 223, 86–98. [Google Scholar] [CrossRef]
- McWilliam, S.J.; Antoine, D.J.; Sabbisetti, V.; Turner, M.A.; Farragher, T.; Bonventre, J.V.; Park, B.K.; Smyth, R.L.; Pirmohamed, M. Mechanism-based urinary biomarkers to identify the potential for aminoglycoside-induced nephrotoxicity in premature neonates: a proof-of-concept study. PLoS ONE 2012, 7, e43809. [Google Scholar] [CrossRef]
- Koren, G.; Pastuszak, A.; Ito, S. Drugs in pregnancy. N. Engl. J. Med. 1998, 338, 1128–1137. [Google Scholar] [CrossRef]
- Neto, J.A.; Oliveira-Filho, R.M.; Simoes, M.J.; Soares, J.M., Jr.; Kulay, L., Jr. Long-term acetaminophen (paracetamol) treatment causes liver and kidney ultra-structural changes during rat pregnancy. Clin. Exp. Obstet Gynecol. 2004, 31, 221–224. [Google Scholar]
- Peng, H.C.; Wang, Y.H.; Wen, C.C.; Wang, W.H.; Cheng, C.C.; Chen, Y.H. Nephrotoxicity assessments of acetaminophen during zebrafish embryogenesis. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 151, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Balbi, A.P.; Marin, E.C.; Francescato, H.D.; Costa, R.S.; Coimbra, T.M. MAPK and angiotensin II receptor in kidney of newborn rats from losartan-treated dams. Pediatr. Nephrol. 2008, 23, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, L.F.; Francescato, H.D.C.; da Silva, C.G.A.; Costa, R.S.; Coimbra, T.M. Calcitriol reduces kidney development disorders in rats provoked by losartan administration during lactation. Sci. Rep. 2017, 7, 11472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kent, A.L.; Douglas-Denton, R.; Shadbolt, B.; Dahlstrom, J.E.; Maxwell, L.E.; Koina, M.E.; Falk, M.C.; Willenborg, D.; Bertram, J.F. Indomethacin, ibuprofen and gentamicin administered during late stages of glomerulogenesis do not reduce glomerular number at 14 days of age in the neonatal rat. Pediatr. Nephrol. 2009, 24, 1143–1149. [Google Scholar] [CrossRef]
- Poureetezadi, S.J.; Cheng, C.N.; Chambers, J.M.; Drummond, B.E.; Wingert, R.A. Prostaglandin signaling regulates nephron segment patterning of renal progenitors during zebrafish kidney development. Elife 2016, 5. [Google Scholar]
- Bueters, R.R.; Klaasen, A.; van den Heuvel, L.P.; Schreuder, M.F. Effect of NSAIDs and diuretics on nephrogenesis in an ex vivo embryogenic kidney model. Birth Defects Res. B Dev. Reprod Toxicol 2013, 98, 486–492. [Google Scholar] [CrossRef]
- Gloor, J.M.; Muchant, D.G.; Norling, L.L. Prenatal maternal indomethacin use resulting in prolonged neonatal renal insufficiency. J. Perinatol. 1993, 13, 425–427. [Google Scholar]
- Kaplan, B.S.; Restaino, I.; Raval, D.S.; Gottlieb, R.P.; Bernstein, J. Renal failure in the neonate associated with in utero exposure to non-steroidal anti-inflammatory agents. Pediatr. Nephrol. 1994, 8, 700–704. [Google Scholar] [CrossRef]
- van der Heijden, B.J.; Carlus, C.; Narcy, F.; Bavoux, F.; Delezoide, A.L.; Gubler, M.C. Persistent anuria, neonatal death, and renal microcystic lesions after prenatal exposure to indomethacin. Am. J. Obstet. Gynecol. 1994, 171, 617–623. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Low Conc. | High Conc. | Embryos Treated per Group | |
|---|---|---|---|
| Acetaminophen | 2.5 mM | 5 mM | 50 |
| Ampicillin | 20 mM | 40 mM | 30 |
| Gentamicin | 7 mM | 14 mM | 30 |
| Indomethacin | 10 µM | 15 µM | 50 |
| Kanamycin | 20 mM | 40 mM | 30 |
| Losartan | 5 mM | 10 mM | 30 |
| Penicillin | 10 mM | 20 mM | 30 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steenbergen, P.J.; Heigwer, J.; Pandey, G.; Tönshoff, B.; Gehrig, J.; Westhoff, J.H. A Multiparametric Assay Platform for Simultaneous In Vivo Assessment of Pronephric Morphology, Renal Function and Heart Rate in Larval Zebrafish. Cells 2020, 9, 1269. https://doi.org/10.3390/cells9051269
Steenbergen PJ, Heigwer J, Pandey G, Tönshoff B, Gehrig J, Westhoff JH. A Multiparametric Assay Platform for Simultaneous In Vivo Assessment of Pronephric Morphology, Renal Function and Heart Rate in Larval Zebrafish. Cells. 2020; 9(5):1269. https://doi.org/10.3390/cells9051269
Chicago/Turabian StyleSteenbergen, Petrus J., Jana Heigwer, Gunjan Pandey, Burkhard Tönshoff, Jochen Gehrig, and Jens H. Westhoff. 2020. "A Multiparametric Assay Platform for Simultaneous In Vivo Assessment of Pronephric Morphology, Renal Function and Heart Rate in Larval Zebrafish" Cells 9, no. 5: 1269. https://doi.org/10.3390/cells9051269
APA StyleSteenbergen, P. J., Heigwer, J., Pandey, G., Tönshoff, B., Gehrig, J., & Westhoff, J. H. (2020). A Multiparametric Assay Platform for Simultaneous In Vivo Assessment of Pronephric Morphology, Renal Function and Heart Rate in Larval Zebrafish. Cells, 9(5), 1269. https://doi.org/10.3390/cells9051269