Genetic Diversity and Population Structure Analysis of Dalbergia Odorifera Germplasm and Development of a Core Collection Using Microsatellite Markers

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and DNA Extraction
2.2. PCR and Capillary Electrophoresis
2.3. Population Structure
2.4. Genetic Diversity Statistics
2.5. Construction and Evaluation of the Core Collection
3. Results
3.1. Polymorphism of 19 Microsatellite Markers
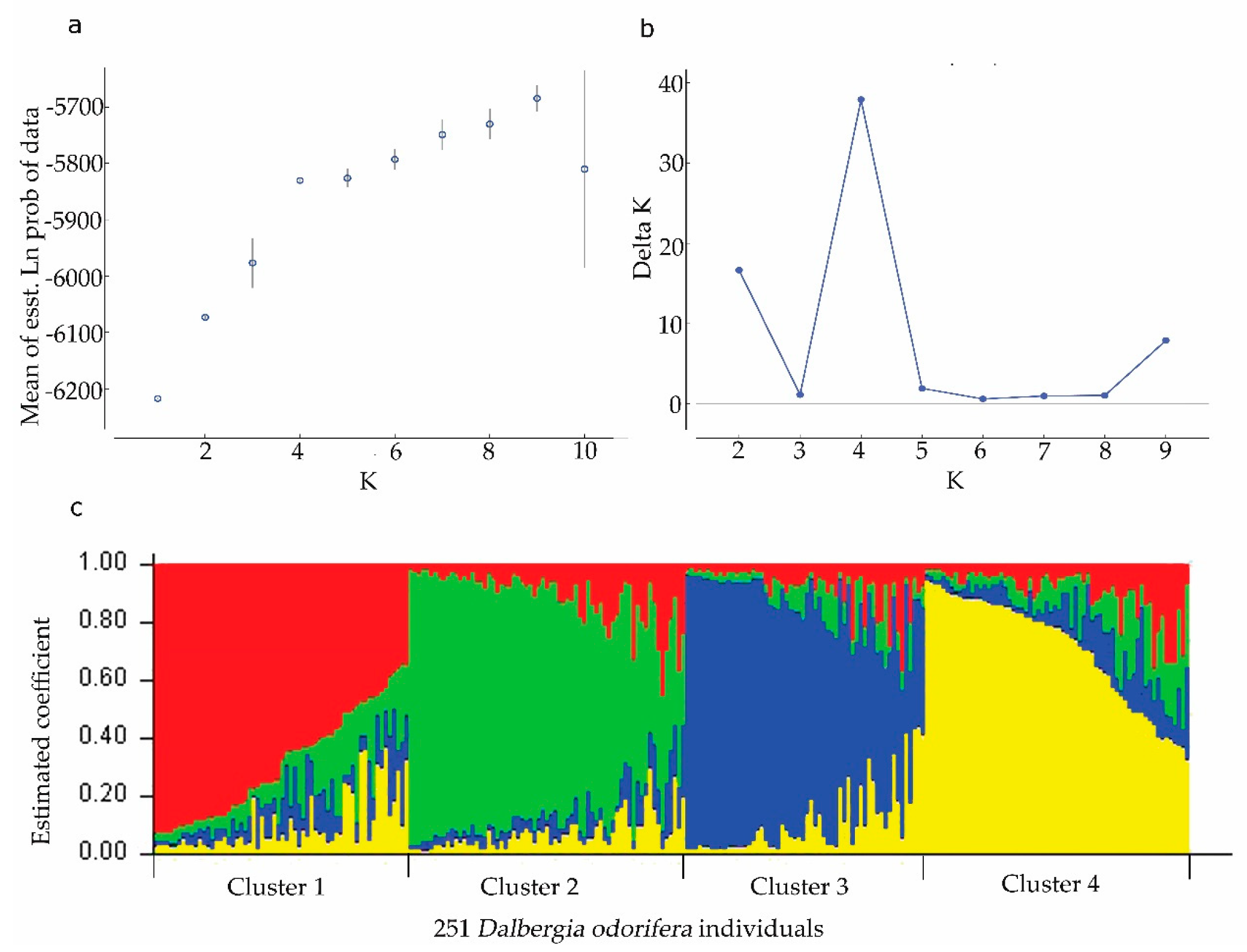
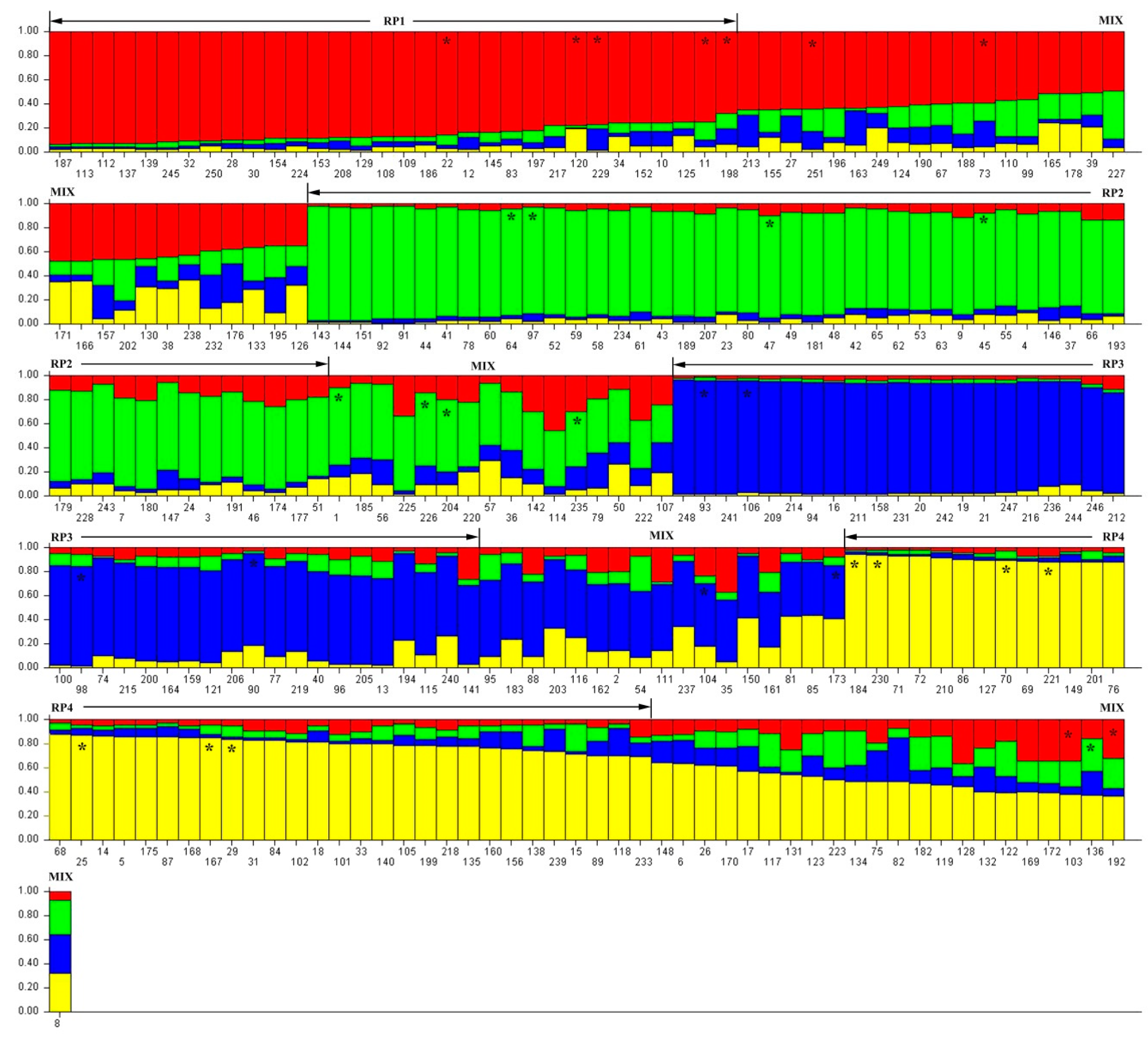
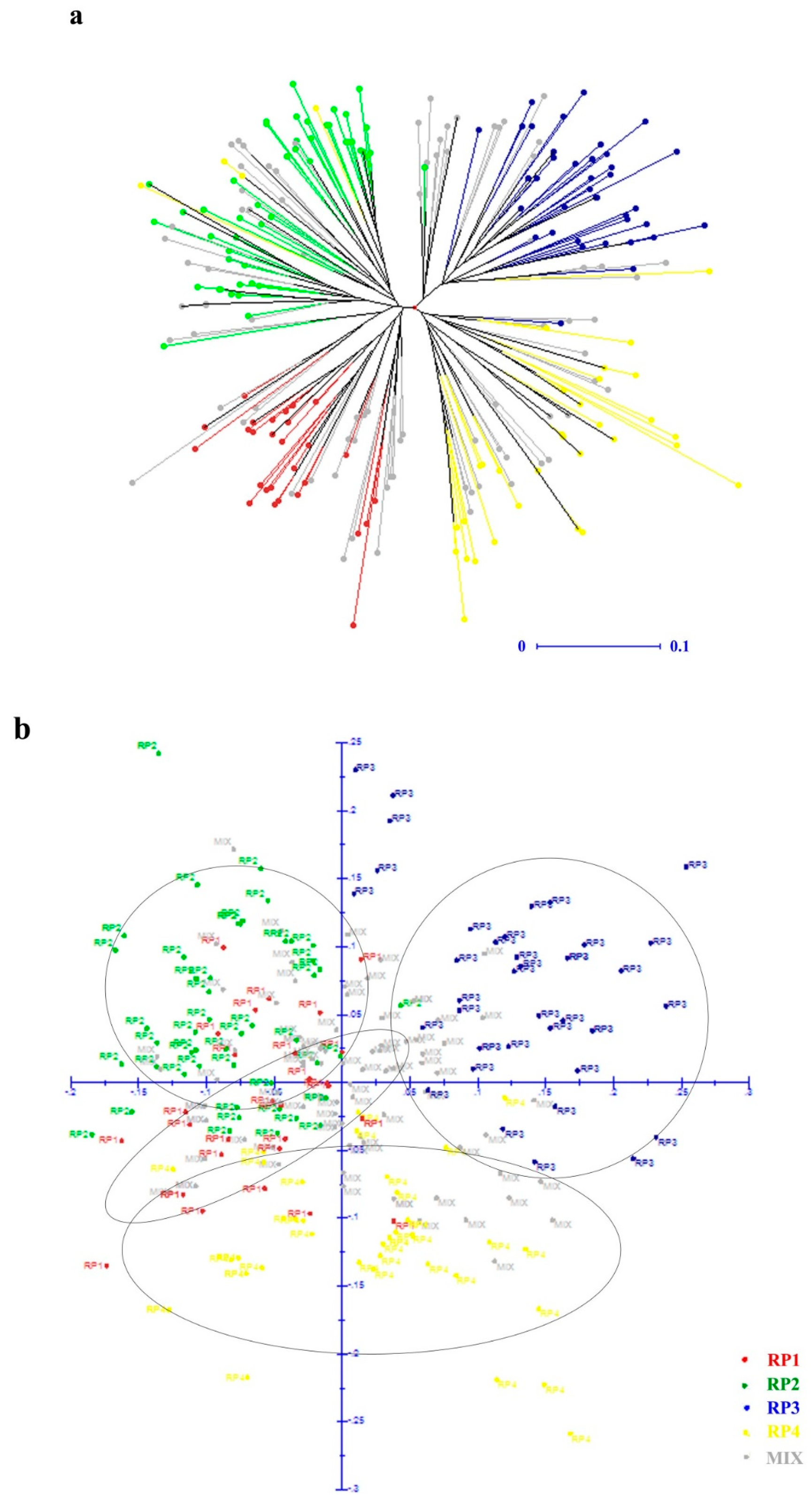
3.2. Population Structure of D. odorifera Germplasm
3.3. Genetic Diversity and Variation of D. odorifera Germplasm
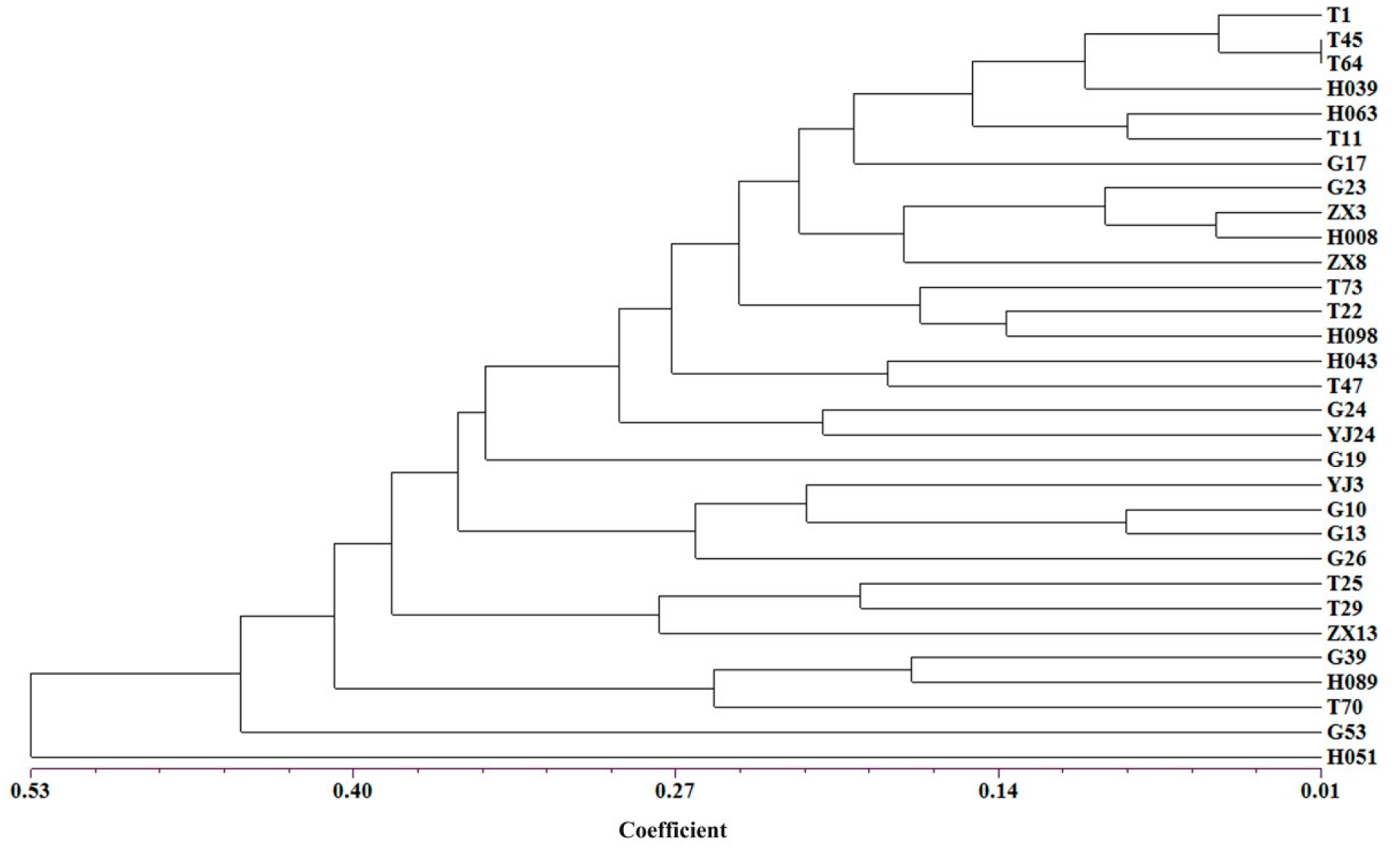
3.4. Core Collection Development of Dalbergia odorifera
4. Discussion
4.1. Genetic Diversity and Population Structure of D. odorifera Germplasm
4.2. The Core Collection of D. odorifera Germplasm
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fan, Z.M.; Wang, D.Y.; Yang, J.M.; Lin, Z.X.; Lin, Y.X.; Yang, A.L.; Fan, H.; Cao, M.; Yuan, S.Y.; Liu, Z.J.; et al. Dalbergia odorifera extract promotes angiogenesis through upregulation of VEGFRs and PI3K/MAPK signaling pathways. J. Ethnopharmacol. 2017, 204, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Li, B.; Keo, S.; Kim, K.S.; Jeong, G.S.; Oh, H.; Kim, Y.C. Inhibitory effect of 9-hydroxy-6,7-dimethoxydalbergiquinol from Dalbergia odorifera on the NF-kappaB-related neuroinflammatory response in lipopolysaccharide-stimulated mouse BV2 microglial cells is mediated by heme oxygenase-1. Int. Immunopharmacol. 2013, 17, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.S.; Park, J.A.; Hwang, J.S.; Ham, S.A.; Yoo, T.; Lee, W.J.; Paek, K.S.; Shin, H.C.; Lee, C.H.; Seo, H.G. A Dalbergia odorifera extract improves the survival of endotoxemia model mice by inhibiting HMGB1 release. BMC Complement. Altern. Med. 2017, 17, 212. [Google Scholar] [CrossRef] [PubMed]
- The, S.N. A review on the medicinal plant Dalbergia odorifera species: Phytochemistry and biological activity. Evid.-Based Complement. Altern. Med. 2017, 2017, 1–27. [Google Scholar] [CrossRef]
- Lee, D.S.; Kim, K.S.; Ko, W.; Li, B.; Keo, S.; Jeong, G.S.; Oh, H.; Kim, Y.C. The neoflavonoid latifolin isolated from MeOH extract of Dalbergia odorifera attenuates inflammatory responses by inhibiting NF-kappaB activation via Nrf2-mediated heme oxygenase-1 expression. Phytother. Res. 2014, 28, 1216–1223. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Dong, W.H.; Zuo, W.J.; Wang, H.; Zhong, H.M.; Mei, W.L.; Dai, H.F. Three new phenolic compounds from Dalbergia odorifera. J. Asian Nat. Prod. Res. 2014, 16, 1109–1118. [Google Scholar] [CrossRef]
- Meng, H.; Chen, D.L.; Yang, Y.; Liu, Y.Y.; Wei, J.H. Sesquiterpenoids with cytotoxicity from heartwood of Dalbergia odorifera. J. Asian Nat. Prod. Res. 2018, 1–7. [Google Scholar] [CrossRef]
- Tao, Y.; Wang, Y. Bioactive sesquiterpenes isolated from the essential oil of Dalbergia odorifera T. Chen. Fitoterapia 2010, 81, 393–396. [Google Scholar] [CrossRef]
- Wang, H.; Dong, W.H.; Zuo, W.J.; Liu, S.; Zhong, H.M.; Mei, W.L.; Dai, H.F. Five new sesquiterpenoids from Dalbergia odorifera. Fitoterapia 2014, 95, 16–21. [Google Scholar] [CrossRef] [PubMed]
- So, T.; Dell, B. Conservation and utilization of threatened hardwood species through reforestation—An example of Afzelia xylocarpa (Kruz.) craib and Dalbergia cochinchinensis Pierre in Cambodia. Pac. Conserv. Biol. 2010, 16, 101–116. [Google Scholar] [CrossRef]
- Yang, Q.X.; Feng, J.D.; Wei, J.H.; Li, R.T.; He, M.J. Genetic diversity of China’ s endangered medicinal plant Dalbergia odorifera. World Sci. Technol. Mod. Tradit. Chin. Med. Mater. Medica 2007, 9, 73–79. [Google Scholar]
- World Conservation Monitoring Centre. Dalbergia odorifera. The IUCN Red List of Threatened Species. 1998: e.T32398A9698077. Available online: http://www.iucnredlist.org/details/32398/0 (accessed on 18 July 2018).
- Liu, F.-M.; Hong, Z.; Yang, Z.-J.; Zhang, N.-N.; Liu, X.-J.; Xu, D.-P. De novo transcriptome analysis of Dalbergia odorifera and transferability of SSR markers developed from the transcriptome. Forests 2019, 10, 98. [Google Scholar] [CrossRef]
- Liu, X.; Xu, D.; Yang, Z.; Zhang, N. Geographic variations in seed germination of Dalbergia odorifera T. Chen in response to temperature. Ind. Crop Prod. 2017, 102, 45–50. [Google Scholar] [CrossRef]
- You, F.M.; Jia, G.; Xiao, J.; Duguid, S.D.; Rashid, K.Y.; Booker, H.M.; Cloutier, S. Genetic Variability of 27 Traits in a Core Collection of Flax (Linum usitatissimum L.). Front. Plant Sci. 2017, 8, 1636. [Google Scholar] [CrossRef]
- Xu, D.P.; Yang, Z.J.; Liang, K.N.; Zhang, N.N.; Zeng, J. Investigation on cold injuries of five valuable tree species in southern China. Sci. Silvae Sin. 2008, 44, 1–2. [Google Scholar]
- Yang, Z.J.; Xu, D.P.; Zeng, J.; Guo, J.Y.; Lin, Q.J.; Yao, Q.D. Investigation on cold injuries for Pterocarpus macrocarpus Kurz in southern China. Sci. Silvae Sin. 2008, 44, 123–127. [Google Scholar]
- Ma, Y.H.; Jia, R.F.; Zeng, J.; Xu, D.P.; Zhang, N.N.; Liu, X.J. Early growth evaluation on six-year-old Dalbergia odorifera T. Chen families. J. Cent. South Univ. For. Technol. 2017, 37, 42–47. [Google Scholar]
- Frankel, O.H. Genetic Perspectives of Germplasm Conservation. Available online: http://books.google.fr/books?hl=en&lr=&id=0yRUkeJFwAMC&oi=fnd&pg=PA161&dq=genetic+erosion+frankel&ots=XXbSegnxMS&sig=MbO4m1MOa5npb7zEbebw1XuGiRY) (accessed on 30 January 2019).
- Lee, H.-Y.; Ro, N.-Y.; Jeong, H.-J.; Kwon, J.-K.; Jo, J.; Ha, Y.; Jung, A.; Han, J.-W.; Venkatesh, J.; Kang, B.-C. Genetic diversity and population structure analysis to construct a core collection from a large Capsicum germplasm. BMC Genet. 2016, 17. [Google Scholar] [CrossRef]
- Mariano, L.C.; Zchonski, F.L.; da Silva, C.M.; Da-Silva, P.R. Genetic variability in a Brazilian apple germplasm collection with low chilling requirements. Peer J. 2019, 6, e6265. [Google Scholar] [CrossRef]
- Liang, W.; Dondini, L.; De Franceschi, P.; Paris, R.; Sansavini, S.; Tartarini, S. Genetic diversity, population structure and construction of a core collection of apple cultivars from Italian germplasm. Plant Mol. Biol. Rep. 2015, 33, 458–473. [Google Scholar] [CrossRef]
- Guzman, L.F.; Machida-Hirano, R.; Borrayo, E.; Cortes-Cruz, M.; Espindola-Barquera, M.D.; Heredia Garcia, E. Genetic structure and selection of a core collection for long term conservation of avocado in Mexico. Front. Plant Sci. 2017, 8, 243. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Lorenzo, S.; Ramos-Cabrer, A.M.; Barreneche, T.; Mattioni, C.; Villani, F.; Díaz-Hernández, M.B.; Martín, L.M.; Martín, Á. Database of European chestnut cultivars and definition of a core collection using simple sequence repeats. Tree Genet. Genomes 2017, 13, 114. [Google Scholar] [CrossRef]
- Ditta, A.; Zhou, Z.; Cai, X.; Wang, X.; Okubazghi, K.W.; Shehzad, M.; Xu, Y.; Hou, Y.; Sajid Iqbal, M.; Khan, M.K.R.; et al. Assessment of genetic diversity, population structure, and evolutionary relationship of uncharacterized genes in a novel germplasm collection of diploid and allotetraploid Gossypium accessions using EST and genomic SSR markers. Int. J. Mol. Sci. 2018, 19, 2401. [Google Scholar] [CrossRef]
- Gomes, S.; Martins-Lopes, P.; Lopes, J.; Guedes-Pinto, H. Assessing genetic diversity in Olea europaea L. using ISSR and SSR markers. Plant Mol. Biol. Rep. 2009, 27, 365–373. [Google Scholar] [CrossRef]
- Boraks, A.; Broders, K.D. Population genetic diversity of the rare hardwood butternut (Juglans cinerea) in the northeastern USA. Tree. Genet. Genomes 2016, 12. [Google Scholar] [CrossRef]
- Nybom, H. Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in plants. Mol. Ecol. 2004, 13, 1143–1155. [Google Scholar] [CrossRef]
- Kong, Q.; Chen, J.; Yong, L.; Ma, Y.; Peng, L.; Wu, S.; Yuan, H.; Bie, Z. Genetic diversity of Cucurbita rootstock germplasm as assessed using simple sequence repeat markers. Sci. Hortic.-Amsterdam 2014, 175, 150–155. [Google Scholar] [CrossRef]
- Kim, K.W.; Chung, H.K.; Cho, G.T.; Ma, K.H.; Chandrabalan, D.; Gwag, J.G.; Kim, T.S.; Cho, E.G.; Park, Y.J. PowerCore: A program applying the advanced M strategy with a heuristic search for establishing core sets. Bioinformatics 2007, 23, 2155–2162. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2010, 14, 2611–2620. [Google Scholar] [CrossRef]
- Porras-Hurtado, L.; Ruiz, Y.; Santos, C.; Phillips, C.; Carracedo, A.; Lareu, M.V. An overview of STRUCTURE: Applications, parameter settings, and supporting software. Front. Genet. 2013, 4, 98. [Google Scholar] [CrossRef]
- Earl, D.A.; Vonholdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Jonathan, M.; Mattias, J.; Rosenberg, N.A.; Itay, M. Clumpak: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef]
- Xu, Q.; Zeng, X.; Lin, B.; Li, Z.; Yuan, H.; Wang, Y.; Tashi, N. A microsatellite diversity analysis and the development of core-set germplasm in a large hulless barley (Hordeum vulgare L.) collection. BMC Genet. 2017, 18, 102. [Google Scholar] [CrossRef] [PubMed]
- Perrier, X.; Jacquemoud-Collet, J.P. DARwin Software. 2006. Available online: http://darwin.cirad.fr/darwin (accessed on 30 January 2019).
- Perrier, X.; Flori, A.; Bonnot, F. Data analysis methods. In Genetic Diversity of Cultivated Tropical Plants; Hamon, P., Seguin, M., Perrier, X., Glaszmann, J.C., Eds.; Enfield, Science Publishers: Montpellier, France, 2003; pp. 43–76. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. Micro-checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Popgene, Version 1.32; the User-Friendly Shareware for Population Genetic Analysismolecular Biology and Biotechnology Center, University of AlbertaEdmonton. Available online: http://www.ualberta.ca/~fyeh (accessed on 23 November 2017).
- Nagy, S.; Poczai, P.; Cernák, I.; Gorji, A.M.; Hegedűs, G.; Taller, J. PICcalc: An online program to calculate polymorphic information content for molecular genetic studies. Biochem. Genet. 2012, 50, 670–672. [Google Scholar] [CrossRef]
- Odong, T.L.; Jansen, J.; Van Eeuwijk, F.A.; Van Hintum, T.J.L. Quality of core collections for effective utilisation of genetic resources review, discussion and interpretation. Theor. Appl. Genet. 2013, 126, 289–305. [Google Scholar] [CrossRef]
- Jansen, J.; van Hintum, T. Genetic distance sampling: A novel sampling method for obtaining core collections using genetic distances with an application to cultivated lettuce. Theor. Appl. Genet. 2007, 114, 421–428. [Google Scholar] [CrossRef]
- Rohlf, F.J. NTSYS-pc: Microcomputer Programs for Numerical Taxonomy and Multivariate Analysis. Am. Stat. 1987, 41, 330. [Google Scholar] [CrossRef]
- Liu, F.-M.; Hong, Z.; Xu, D.-P.; Jia, H.-Y.; Zhang, N.-N.; Liu, X.-J.; Yang, Z.-J.; Lu, M.-Z. Genetic diversity of endangered endemic Dalbergia odorifera revealed by SSR markers. Forests 2019, 10, 225. [Google Scholar] [CrossRef]
- White, T.L.; Adams, W.T.; Neale, D.B. Forest Genetics; CABI Publishing: Boston, MA, USA, 2007; pp. 149–186. [Google Scholar]
- Ferrer, M.M.; Eguiarte, L.E.; Montana, C. Genetic structure and outcrossing rates in Flourensia cernua (Asteraceae) growing at different densities in the South-western Chihuahuan Desert. Ann. Bot. 2004, 94, 419–426. [Google Scholar] [CrossRef]
- Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogel, J.; Tingey, S.; Rafalski, A. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- Yang, X.; Song, J.; Todd, J.; Peng, Z.; Paudel, D.; Luo, Z.; Ma, X.; You, Q.; Hanson, E.; Zhao, Z.; et al. Target enrichment sequencing of 307 germplasm accessions identified ancestry of ancient and modern hybrids and signatures of adaptation and selection in sugarcane (Saccharum spp.), a ‘sweet’ crop with ‘bitter’ genomes. Plant Biotechnol. J. 2019, 17, 488–498. [Google Scholar] [CrossRef]
- Ambreen, H.; Kumar, S.; Kumar, A.; Agarwal, M.; Jagannath, A.; Goel, S. Association mapping for important agronomic traits in Safflower (Carthamus tinctorius L.) core collection using microsatellite markers. Front. Plant Sci. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, M.; Sharma, A. Assessment of genetic diversity in Dalbergia sissoo clones through RAPD profiling. J. For. Res. 2011, 22, 393. [Google Scholar] [CrossRef]
- Andrianoelina, O.; Rakotondraoelina, H.; Ramamonjisoa, L.; Maley, J.; Danthu, P.; Bouvet, J.-M. Genetic diversity of Dalbergia monticola (Fabaceae) an endangered tree species in the fragmented oriental forest of Madagascar. Biodivers. Conserv. 2006, 15, 1109–1128. [Google Scholar] [CrossRef]
- Buzatti, R.S.D.O.; Ribeiro, R.A.; Filho, J.P.D.L.; Lovato, M.B. Fine-scale spatial genetic structure of Dalbergia nigra (Fabaceae), a threatened and endemic tree of the Brazilian Atlantic forest. Genet. Mol. Biol. 2012, 35, 838–846. [Google Scholar] [CrossRef] [PubMed]
- Hartvig, I.; So, T.; Changtragoon, S.; Tran, H.T.; Bouamanivong, S.; Theilade, I.; Kjær, E.D.; Nielsen, L.R. Population genetic structure of the endemic rosewoods Dalbergia cochinchinensis and D. oliveri at a regional scale reflects the Indochinese landscape and life-history traits. Ecol. Evol. 2018, 8, 530–545. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.S.; Chung, M.G. Genetic variation and population structure in Korean endemic species: IV. Hemerocallis hakuunensis (Liliaceae). J. Plant Res. 1997, 110, 209–217. [Google Scholar] [CrossRef]
- Zhong, T.; Zhao, G.; Lou, Y.; Lin, X.; Guo, X. Genetic diversity analysis of Sinojackia microcarpa, a rare tree species endemic in China, based on simple sequence repeat markers. J. For. Res. 2018. [Google Scholar] [CrossRef]
- Gichira, A.W.; Li, Z.-Z.; Saina, J.K.; Hu, G.-W.; Gituru, R.W.; Wang, Q.-F.; Chen, J.-M. Demographic history and population genetic structure of Hagenia abyssinica (Rosaceae), a tropical tree endemic to the Ethiopian highlands and eastern African mountains. Tree Genet. Genomes 2017, 13, 72. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Godt, M.J.W.; Sherman-Broyles, S.L. Factors influencing levels of genetic diversity in woody plant species. New For. 1992, 6, 95–124. [Google Scholar] [CrossRef]
- Wang, B.Y.; Shi, L.; Ruan, Z.Y.; Deng, J. Genetic diversity and differentiation in Dalbergia sissoo (Fabaceae) as revealed by RAPD. Genet. Mol. Res. 2011, 10, 114. [Google Scholar] [CrossRef]
- Li, X.; Li, M.; Hou, L.; Zhang, Z.; Pang, X.; Li, Y. De novo transcriptome assembly and population genetic analyses for an endangered chinese endemic Acer miaotaiense (Aceraceae). Genes 2018, 9, 378. [Google Scholar] [CrossRef]
- Li, J.; Liu, X.-B.; Zhao, Z.-W.; Yang, Z.L. Genetic diversity, core collection and breeding history of Pleurotus ostreatus in China. Mycoscience 2019, 60, 14–24. [Google Scholar] [CrossRef]
- Yu, J.Z.; Fang, D.D.; Kohel, R.J.; Ulloa, M.; Hinze, L.L.; Percy, R.G.; Zhang, J.; Chee, P.; Scheffler, B.E.; Jones, D.C. Development of a core set of SSR markers for the characterization of Gossypium germplasm. Euphytica 2012, 187, 203–213. [Google Scholar] [CrossRef]
- Bengtsson, B.O.; Weibull, P.; Ghatnekar, L. The Loss of Alleles by Sampling: A study of the common outbreeding grass Festuca ovina over three geographic scales. Hereditas 1995, 122, 221–238. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Size | Na | Ne | Ho | He | GD | PIC | I | Nm | F | FNA |
|---|---|---|---|---|---|---|---|---|---|---|---|
| S01 | 251 | 4 | 2 | 0.46 | 0.50 | 0.5 | 0.43 | 0.87 | 1.93 | 0.08 | 0.03 no |
| S02 | 251 | 3 | 1.06 | 0.05 | 0.06 | 0.06 | 0.06 | 0.15 | 5.63 | 0.11 | 0.0 no |
| S03 | 249 | 3 | 1.98 | 0.38 | 0.50 | 0.49 | 0.37 | 0.70 | 2.53 | 0.23 | 0.08 * |
| S04 | 251 | 4 | 1.64 | 0.26 | 0.39 | 0.39 | 0.36 | 0.72 | 0.44 | 0.35 | 0.1 * |
| S07 | 251 | 2 | 1.87 | 0.44 | 0.47 | 0.47 | 0.36 | 0.66 | 3.22 | 0.05 | 0.02 no |
| S08 | 251 | 4 | 1.46 | 0.29 | 0.31 | 0.31 | 0.29 | 0.62 | 5.67 | 0.06 | 0.01 no |
| S09 | 251 | 4 | 2.39 | 0.45 | 0.58 | 0.58 | 0.53 | 1.08 | 2.60 | 0.23 | 0.08 * |
| S10 | 250 | 4 | 1.92 | 0.46 | 0.48 | 0.48 | 0.42 | 0.83 | 3.37 | 0.04 | 0.01 no |
| S11 | 251 | 2 | 1.62 | 0.36 | 0.38 | 0.38 | 0.31 | 0.57 | 2.49 | 0.05 | 0.01 no |
| S12 | 251 | 2 | 1.42 | 0.25 | 0.30 | 0.30 | 0.25 | 0.47 | 2.73 | 0.15 | 0.04 no |
| S21 | 250 | 7 | 3.48 | 0.69 | 0.71 | 0.71 | 0.66 | 1.34 | 2.45 | 0.03 | 0.01 no |
| S22 | 251 | 6 | 1.28 | 0.19 | 0.22 | 0.22 | 0.21 | 0.53 | 9.45 | 0.13 | 0.02 * |
| S23 | 251 | 6 | 1.04 | 0.02 | 0.04 | 0.04 | 0.04 | 0.12 | 33.31 | 0.33 | 0.01 * |
| S24 | 251 | 6 | 2.33 | 0.49 | 0.57 | 0.57 | 0.50 | 0.99 | 1.23 | 0.15 | 0.05 * |
| S26 | 251 | 3 | 1.03 | 0.02 | 0.03 | 0.03 | 0.03 | 0.08 | 9.53 | 0.28 | 0.01 * |
| S27 | 251 | 4 | 1.3 | 0.21 | 0.23 | 0.23 | 0.20 | 0.41 | 6.58 | 0.08 | 0.01 no |
| S28 | 251 | 2 | 1.7 | 0.29 | 0.41 | 0.41 | 0.33 | 0.6 | 2.18 | 0.29 | 0.08 * |
| S29 | 251 | 6 | 1.91 | 0.44 | 0.48 | 0.48 | 0.39 | 0.78 | 1.07 | 0.07 | 0.02 no |
| S30 | 251 | 5 | 1.92 | 0.43 | 0.48 | 0.48 | 0.40 | 0.79 | 1.68 | 0.11 | 0.04 no |
| Mean | 251 | 4.05 | 1.76 | 0.33 | 0.38 | 0.38 | 0.32 | 0.65 | 5.16 | 0.15 | - |
| Population | Size | Wild | Alleles | Na | Ne | Np | Ho | He | GD | PPB % |
|---|---|---|---|---|---|---|---|---|---|---|
| MIX | 86 | 13 | 66 | 3.47 | 1.74 | 9 | 0.34 | 0.36 | 0.36 | 100.00 |
| RP1 | 32 | 5 | 48 | 2.53 | 1.63 | 2 | 0.34 | 0.33 | 0.32 | 100.00 |
| RP2 | 51 | 3 | 48 | 2.53 | 1.62 | 1 | 0.33 | 0.32 | 0.32 | 94.74 |
| RP3 | 41 | 15 | 53 | 2.79 | 1.55 | 2 | 0.27 | 0.30 | 0.30 | 89.47 |
| RP4 | 41 | 6 | 51 | 2.68 | 1.84 | 3 | 0.33 | 0.38 | 0.37 | 94.74 |
| Mean | 50.2 | 8.4 | 53.2 | 2.80 | 1.68 | - | 0.32 | 0.34 | 0.33 | 95.79 |
| Total a | 251 | 42 | 77 | 4.05 | 1.76 | - | 0.33 | 0.38 | 0.38 | 100 |
| Source | d.f. | Sum of Square | Mean of Square | Variance Components | Percentage of Variation |
|---|---|---|---|---|---|
| Among populations | 4 | 171.788 | 42.947 | 0.408 | 11% *** |
| Within populations | 497 | 1621.023 | 6.527 | 3.264 | 89% |
| Among Individuals | 246 | 844.523 | 3.433 | 0.170 | 5% *** |
| Within Individuals | 251 | 776.500 | 3.094 | 3.094 | 84% |
| Total | 501 | 1792.811 | 3.671 | 100% |
| Population | MIX | RP1 | RP2 | RP3 | RP4 |
|---|---|---|---|---|---|
| MIX | |||||
| RP1 | 0.032 | ||||
| RP2 | 0.031 | 0.065 | |||
| RP3 | 0.041 | 0.091 | 0.095 | ||
| RP4 | 0.032 | 0.071 | 0.064 | 0.087 |
| Population | Size | Na a | Ne a | Ho a | He a | GD a |
|---|---|---|---|---|---|---|
| Core collection | 31 | 4.05 | 1.96 | 0.34 | 0.44 | 0.44 |
| Whole database | 251 | 4.05 | 1.76 | 0.33 | 0.38 | 0.38 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, F.-M.; Zhang, N.-N.; Liu, X.-J.; Yang, Z.-J.; Jia, H.-Y.; Xu, D.-P. Genetic Diversity and Population Structure Analysis of Dalbergia Odorifera Germplasm and Development of a Core Collection Using Microsatellite Markers. Genes 2019, 10, 281. https://doi.org/10.3390/genes10040281
Liu F-M, Zhang N-N, Liu X-J, Yang Z-J, Jia H-Y, Xu D-P. Genetic Diversity and Population Structure Analysis of Dalbergia Odorifera Germplasm and Development of a Core Collection Using Microsatellite Markers. Genes. 2019; 10(4):281. https://doi.org/10.3390/genes10040281
Chicago/Turabian StyleLiu, Fu-Mei, Ning-Nan Zhang, Xiao-Jin Liu, Zeng-Jiang Yang, Hong-Yan Jia, and Da-Ping Xu. 2019. "Genetic Diversity and Population Structure Analysis of Dalbergia Odorifera Germplasm and Development of a Core Collection Using Microsatellite Markers" Genes 10, no. 4: 281. https://doi.org/10.3390/genes10040281
APA StyleLiu, F. -M., Zhang, N. -N., Liu, X. -J., Yang, Z. -J., Jia, H. -Y., & Xu, D. -P. (2019). Genetic Diversity and Population Structure Analysis of Dalbergia Odorifera Germplasm and Development of a Core Collection Using Microsatellite Markers. Genes, 10(4), 281. https://doi.org/10.3390/genes10040281