ORTHOSCOPE Analysis Reveals the Presence of the Cellulose Synthase Gene in All Tunicate Genomes but Not in Other Animal Genomes


Abstract
:1. Introduction
2. Methods
2.1. Data Collection
2.2. Molecular Phylogenetic Analysis
3. Results and Discussion
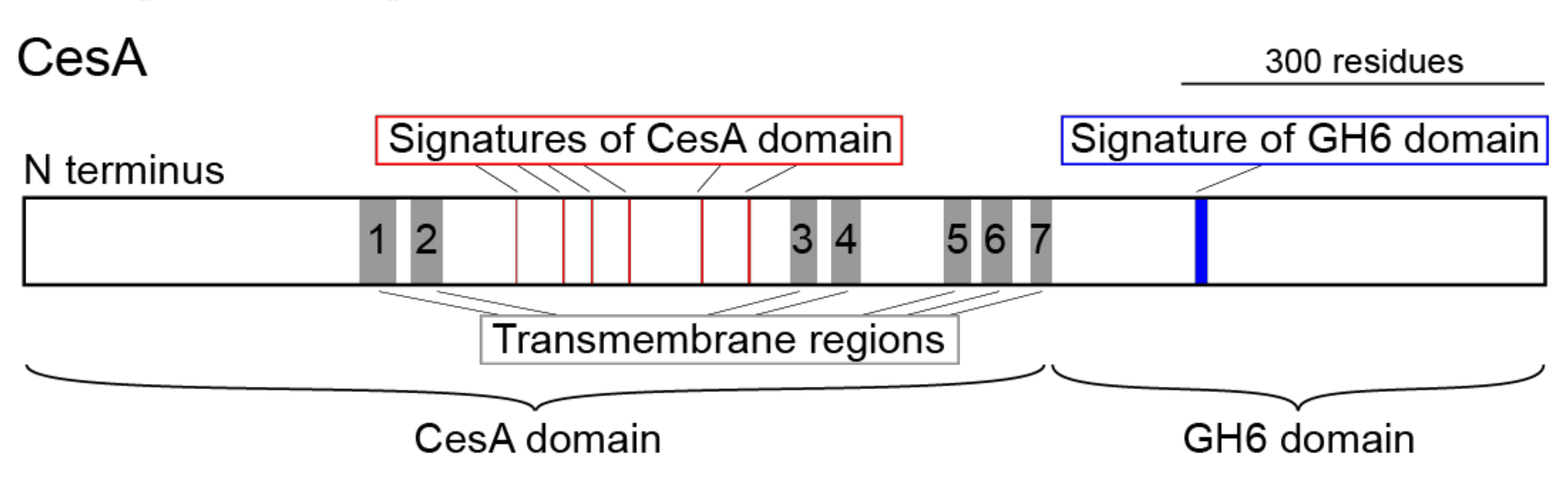
3.1. CesA is Present in the Genomes of All Tunicates but Not in the Genomes of the Other Metazoans Examined
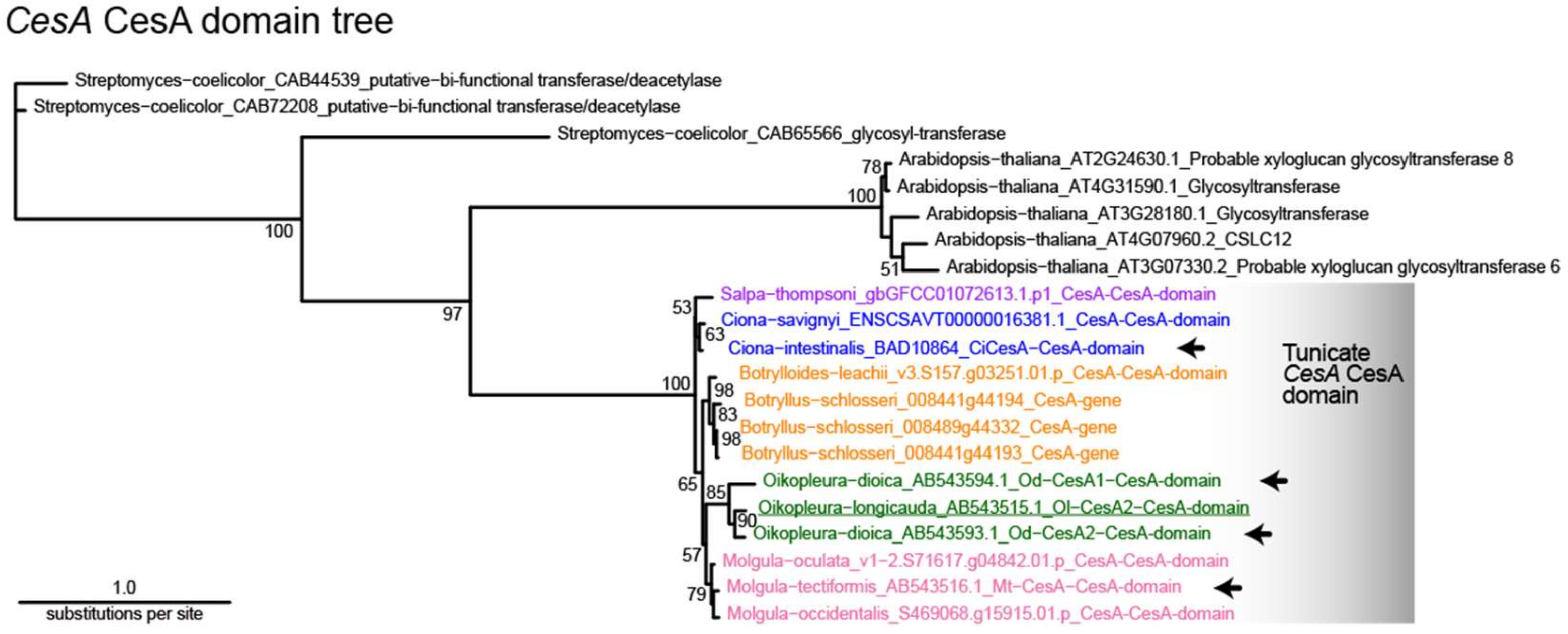
3.2. Molecular Phylogenetic Analysis with the CesA Domain
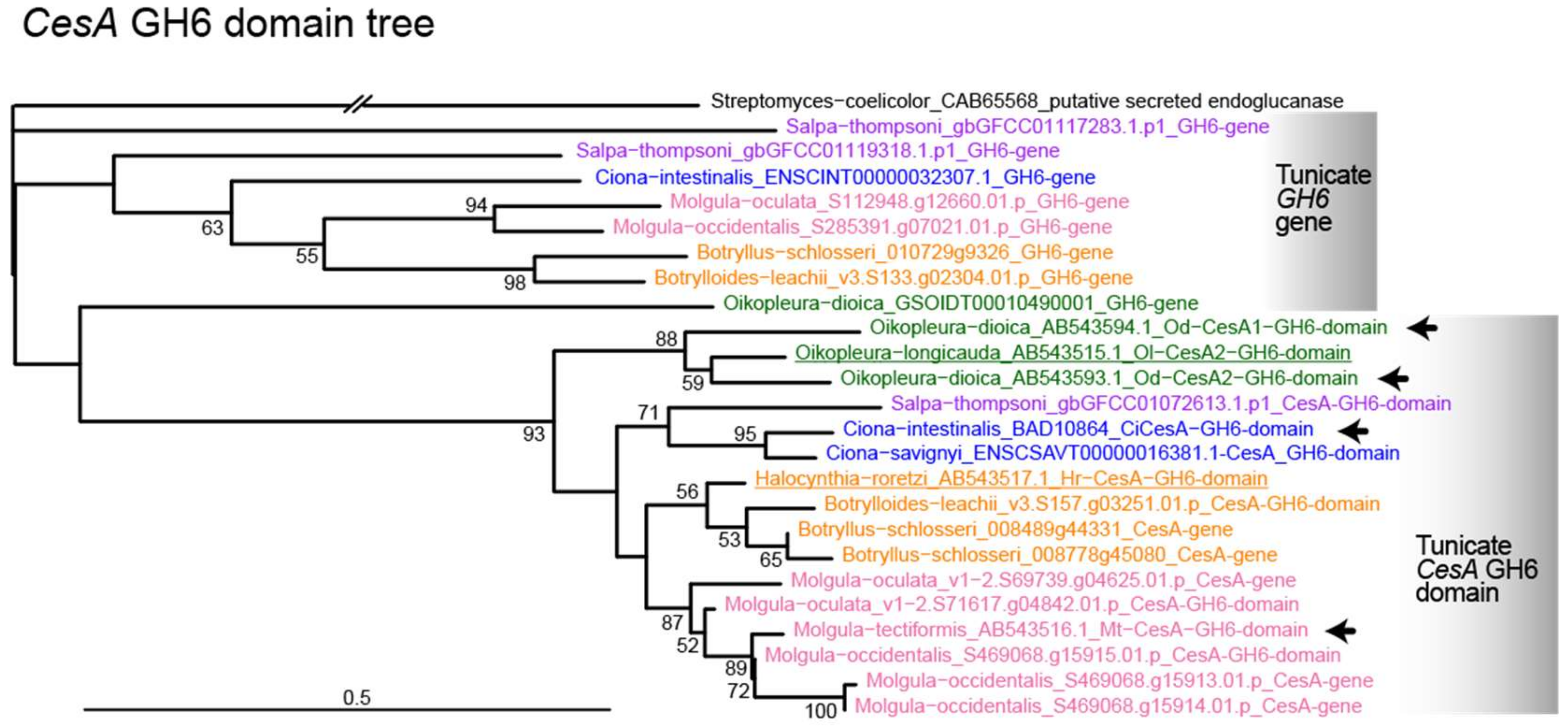
3.3. Molecular Phylogenetic Analysis with the GH6 Domain
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cresswell, R. Aristotle’s History of Animals. In Ten Books; Translated by Richard Creswell; George Bell & Sons: London, UK, 1862. [Google Scholar]
- Lamarck, J.B. Histoire Naturelle des Animaux sans Vertebres, vol. III: Tuniciers; Déterville: Paris, France, 1816. [Google Scholar]
- Haeckel, E. Generelle Morphologie der Organismen: Allgemeine Grundzüge der Organischen Formen-Wissenschaft, Mechanisch Begründet Durch die von Charles Darwin Reformirte Descendenz-Theorie; G. Reimer: Berlin, Germany, 1866. [Google Scholar]
- Balfour, F.M. A Treatise on Comparative Embryology; Macmillan and, Co.: London, UK, 1880. [Google Scholar]
- Lankester, E.R. Notes on the Embryology and classification of the Animal kingdom: Comprising a revision of speculations relative to the origin and significance of the germ-layers. Q. J. Microsc. Sci. 1877, 68, 399–454. [Google Scholar]
- Schmidt, C. Zur vergleichenden Physiologie der wirbellosen Thiere. Justus Liebigs Ann. Chem. 1845, 54, 284–330. [Google Scholar] [CrossRef]
- Belton, P.S.; Tanner, S.F.; Cartier, N.; Chanzy, H. High-resolution solid-state C-13 nuclear magnetic-resonance spectroscopy of tunicin, an animal cellulose. Macromolecules 1989, 22, 1615–1617. [Google Scholar] [CrossRef]
- Matthysse, A.G.; Deschet, K.; Williams, M.; Marry, M.; White, A.R.; Smith, W.C. A functional cellulose synthase from ascidian epidermis. Proc. Natl. Acad. Sci. USA 2004, 101, 986–991. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Yamada, L.; Satou, Y.; Azuma, J.; Satoh, N. The evolutionary origin of animal cellulose synthase. Dev. Genes Evol. 2004, 214, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Nishino, A.; Horikawa, Y.; Hirose, E.; Sugiyama, J.; Satoh, N. The crystalline phase of cellulose changes under developmental control in a marine chordate. Cell. Mol. Life Sci. 2011, 68, 1623–1631. [Google Scholar] [CrossRef]
- Sagane, Y.; Zech, K.; Bouquet, J.M.; Schmid, M.; Bal, U.; Thompson, E.M. Functional specialization of cellulose synthase genes of prokaryotic origin in chordate larvaceans. Development 2010, 137, 1483–1492. [Google Scholar] [CrossRef]
- Sasakura, Y.; Nakashima, K.; Awazu, S.; Matsuoka, T.; Nakayama, A.; Azuma, J.; Satoh, N. Transposon-mediated insertional mutagenesis revealed the functions of animal cellulose synthase in the ascidian Ciona intestinalis. Proc. Natl. Acad. Sci. USA 2005, 102, 15134–15139. [Google Scholar] [CrossRef] [PubMed]
- Satoh, N. Chordate Origins and Evolution: The Molecular Evolutionary Road to Vertebrates; Elsevier: Boston, MA, USA, 2016. [Google Scholar]
- Inoue, J.; Satoh, N. ORTHOSCOPE: An automatic web tool for phylogenetically inferring bilaterian orthogroups with user-selected taxa. Mol. Biol. Evol. 2019, 36, 621–631. [Google Scholar] [CrossRef]
- Blanchoud, S.; Rutherford, K.; Zondag, L.; Gemmell, N.J.; Wilson, M.J. De novo draft assembly of the Botrylloides leachii genome provides further insight into tunicate evolution. Sci. Rep. 2018, 8, 5518. [Google Scholar] [CrossRef] [PubMed]
- Voskoboynik, A.; Neff, N.F.; Sahoo, D.; Newman, A.M.; Pushkarev, D.; Koh, W.; Passarelli, B.; Fan, H.C.; Mantalas, G.L.; Palmeri, K.J.; et al. The genome sequence of the colonial chordate, Botryllus schlosseri. Elife 2013, 2, e00569. [Google Scholar] [CrossRef]
- Dehal, P.; Satou, Y.; Campbell, R.K.; Chapman, J.; Degnan, B.; De Tomaso, A.; Davidson, B.; Di Gregorio, A.; Gelpke, M.; Goodstein, D.M.; et al. The draft genome of Ciona intestinalis: Insights into chordate and vertebrate origins. Science 2002, 298, 2157–2167. [Google Scholar] [CrossRef]
- Small, K.S.; Brudno, M.; Hill, M.M.; Sidow, A. A haplome alignment and reference sequence of the highly polymorphic Ciona savignyi genome. Genome Biol. 2007, 8, R41. [Google Scholar] [CrossRef] [PubMed]
- Stolfi, A.; Lowe, E.K.; Racioppi, C.; Ristoratore, F.; Brown, C.T.; Swalla, B.J.; Christiaen, L. Divergent mechanisms regulate conserved cardiopharyngeal development and gene expression in distantly related ascidians. Elife 2014, 3, e03728. [Google Scholar] [CrossRef]
- Danks, G.; Campsteijn, C.; Parida, M.; Butcher, S.; Doddapaneni, H.; Fu, B.; Petrin, R.; Metpally, R.; Lenhard, B.; Wincker, P.; et al. OikoBase: A genomics and developmental transcriptomics resource for the urochordate Oikopleura dioica. Nucleic Acids Res. 2013, 41, D845–D853. [Google Scholar] [CrossRef] [PubMed]
- Jue, N.K.; Batta-Lona, P.G.; Trusiak, S.; Obergfell, C.; Bucklin, A.; O’Neill, M.J.; O’Neill, R.J. Rapid evolutionary rates and unique genomic signatures discovered in the first reference genome for the southern ocean salp, Salpa thompsoni (Urochordata, Thaliacea). Genome Biol. Evol. 2016, 8, 3171–3186. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Capella-Gutierrez, S.; Silla-Martinez, J.M.; Gabaldon, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Suyama, M.; Torrents, D.; Bork, P. PAL2NAL: Robust conversion of protein sequence alignments into the corresponding codon alignments. Nucleic Acids Res. 2006, 34, W609–W612. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. Estimating the pattern of nucleotide substitution. J. Mol. Evol. 1994, 39, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. Maximum likelihood phylogenetic estimation from DNA sequences with variable rates over sites: Approximate method. J. Mol. Evol. 1994, 39, 306–314. [Google Scholar] [CrossRef]
- Danchin, E.G.J.; Rosso, M.N.; Vieira, P.; de Almeida-Engler, J.; Coutinho, P.M.; Henrissat, B.; Abad, P. Multiple lateral gene transfers and duplications have promoted plant parasitism ability in nematodes. Proc. Natl. Acad. Sci. USA 2010, 107, 17651–17656. [Google Scholar] [CrossRef] [PubMed]
- Mayer, W.E.; Schuster, L.N.; Bartelmes, G.; Dieterich, C.; Sommer, R.J. Horizontal gene transfer of microbial cellulases into nematode genomes is associated with functional assimilation and gene turnover. BMC Evol. Biol. 2011, 11. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.A.; Pease, J.B. Heterogeneous molecular processes among the causes of how sequence similarity scores can fail to recapitulate phylogeny. Brief. Bioinform. 2017, 18, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Delsuc, F.; Philippe, H.; Tsagkogeorga, G.; Simion, P.; Tilak, M.K.; Turon, X.; Lopez-Legentil, S.; Piette, J.; Lemaire, P.; Douzery, E.J.P. A phylogenomic framework and timescale for comparative studies of tunicates. BMC Biol. 2018, 16, 39. [Google Scholar] [CrossRef] [PubMed]
- Holland, L.Z. Tunicates. Curr. Biol. 2016, 26, R146–R152. [Google Scholar] [CrossRef] [PubMed]
- Satoh, N. Developmental Genomics of Ascidians; Wiley-Blackwell: Hoboken, NJ, USA, 2014. [Google Scholar]
- Satoh, N.; Rokhsar, D.; Nishikawa, T. Chordate evolution and the three-phylum system. Proc. R. Soc. B 2014, 281. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| # of BLAST Hits 1 | # of Related Genes/Orthologs 1 | ||||
|---|---|---|---|---|---|
| Classification | Species | CesA | GH6 | CesA | GH6 |
| Bacteria | Streptomyces coelicolor | 3 | 1 | 3 | 1 |
| Viridiplantae | Arabidopsis thaliana | 5 | 0 | 5 | 0 |
| Fungi | Saccharomyces cerevisiae | 0 | 0 | 0 | 0 |
| Ichthyosporea | Capsaspora owczarzaki | 0 | 0 | 0 | 0 |
| Choanoflagellida | Salpingoeca rosetta | 0 | 0 | 0 | 0 |
| Monosiga brevicollis | 1 | 0 | 0 | 0 | |
| Metazoa | |||||
| Porifera | Amphimedon queenslandica | 0 | 0 | 0 | 0 |
| Placozoa | Trichoplax adhaerens | 0 | 0 | 0 | 0 |
| Cnidaria | Thelohanellus kitauei | 0 | 0 | 0 | 0 |
| Hydra vulgaris | 0 | 0 | 0 | 0 | |
| Acropora digitifera | 0 | 0 | 0 | 0 | |
| Ctenophora | Mnemiopsis leidyi | 1 | 0 | 0 | 0 |
| Bilateria | |||||
| Protostomia | |||||
| Platyhelminthes | Schistosoma mansoni | 0 | 0 | 0 | 0 |
| Annelida | Capitella teleta | 0 | 0 | 0 | 0 |
| Nemertea | Notospermus geniculatus | 0 | 0 | 0 | 0 |
| Brachiopoda | Phoronis australis | 0 | 0 | 0 | 0 |
| Cephalopoda | Octopus bimaculoides | 1 | 0 | 0 | 0 |
| Gastropoda | Biomphalaria glabrata | 0 | 0 | 0 | 0 |
| Bivalvia | Crassostrea gigas | 0 | 0 | 0 | 0 |
| Pinctada fucata | 0 | 0 | 0 | 0 | |
| Priapulus caudatus | 0 | 0 | 0 | 0 | |
| Nematoda | Trichinella spiralis | 0 | 0 | 0 | 0 |
| Caenorhabditis elegans | 0 | 0 | 0 | 0 | |
| Merostomata | Limulus polyphemus | 0 | 0 | 0 | 0 |
| Chilopoda | Strigamia maritima | 0 | 0 | 0 | 0 |
| Malacostraca | Hyalella azteca | 0 | 0 | 0 | 0 |
| Insecta | Pediculus humanus | 0 | 0 | 0 | 0 |
| Zootermopsis nevadensis | 0 | 0 | 0 | 0 | |
| Rhodnius prolixus | 0 | 0 | 0 | 0 | |
| Nasonia vitripennis | 0 | 0 | 0 | 0 | |
| Dendroctonus ponderosae | 0 | 0 | 0 | 0 | |
| Bombyx mori | 0 | 0 | 0 | 0 | |
| Drosophila melanogaster | 0 | 0 | 0 | 0 | |
| Deuterostomia | |||||
| Echinodermata | Strongylocentrotus purpuratus | 0 | 0 | 0 | 0 |
| Acanthaster planci | 0 | 0 | 0 | 0 | |
| Hemichordata | Saccoglossus kowalevskii | 0 | 0 | 0 | 0 |
| Ptychodera flava | 0 | 0 | 0 | 0 | |
| Cephalochordata | Branchiostoma belcheri | 0 | 0 | 0 | 0 |
| Branchiostoma floridae | 0 | 0 | 0 | 0 | |
| Vertebrata | Gallus gallus | 0 | 0 | 0 | 0 |
| Homo sapiens | 0 | 0 | 0 | 0 | |
| Tunicata | |||||
| Appendicularia | Oikopleura dioica | 2 | 3 | 2 | 2 |
| Thaliacea | Salpa thompsoni | 1 | 3 | 1 | 1 |
| Stolidobranchia | Botrylloides leachii | 1 | 2 | 1 | 1 |
| Botryllus schlosseri | 3 | 3 | 3 | 2 | |
| Molgula occidentalis | 1 | 4 | 1 | 3 | |
| Molgula oculate | 1 | 3 | 1 | 2 | |
| Enterogona | Ciona savignyi | 1 | 1 | 1 | 1 |
| Ciona intestinalis | 1 | 2 | 1 | 1 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inoue, J.; Nakashima, K.; Satoh, N. ORTHOSCOPE Analysis Reveals the Presence of the Cellulose Synthase Gene in All Tunicate Genomes but Not in Other Animal Genomes. Genes 2019, 10, 294. https://doi.org/10.3390/genes10040294
Inoue J, Nakashima K, Satoh N. ORTHOSCOPE Analysis Reveals the Presence of the Cellulose Synthase Gene in All Tunicate Genomes but Not in Other Animal Genomes. Genes. 2019; 10(4):294. https://doi.org/10.3390/genes10040294
Chicago/Turabian StyleInoue, Jun, Keisuke Nakashima, and Noriyuki Satoh. 2019. "ORTHOSCOPE Analysis Reveals the Presence of the Cellulose Synthase Gene in All Tunicate Genomes but Not in Other Animal Genomes" Genes 10, no. 4: 294. https://doi.org/10.3390/genes10040294
APA StyleInoue, J., Nakashima, K., & Satoh, N. (2019). ORTHOSCOPE Analysis Reveals the Presence of the Cellulose Synthase Gene in All Tunicate Genomes but Not in Other Animal Genomes. Genes, 10(4), 294. https://doi.org/10.3390/genes10040294