Finding the Unicorn, a New Mouse Model of Midfacial Clefting


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Unicorn Mouse Line Recovery and Husbandry
2.2. Embryo Collection
2.3. Skeletal Preparations
2.4. Pseudo-SEM and Morphometric Analysis
2.5. Histology Analysis
2.6. In Situ Hybridization
2.7. Proliferation and Apoptotic Studies
2.8. Imaging
3. Results
3.1. Unicorn Mice Develop a Midfacial Cleft with a Bifurcated Nasal Septum
3.2. Genetics of the Unicorn Mouse Line
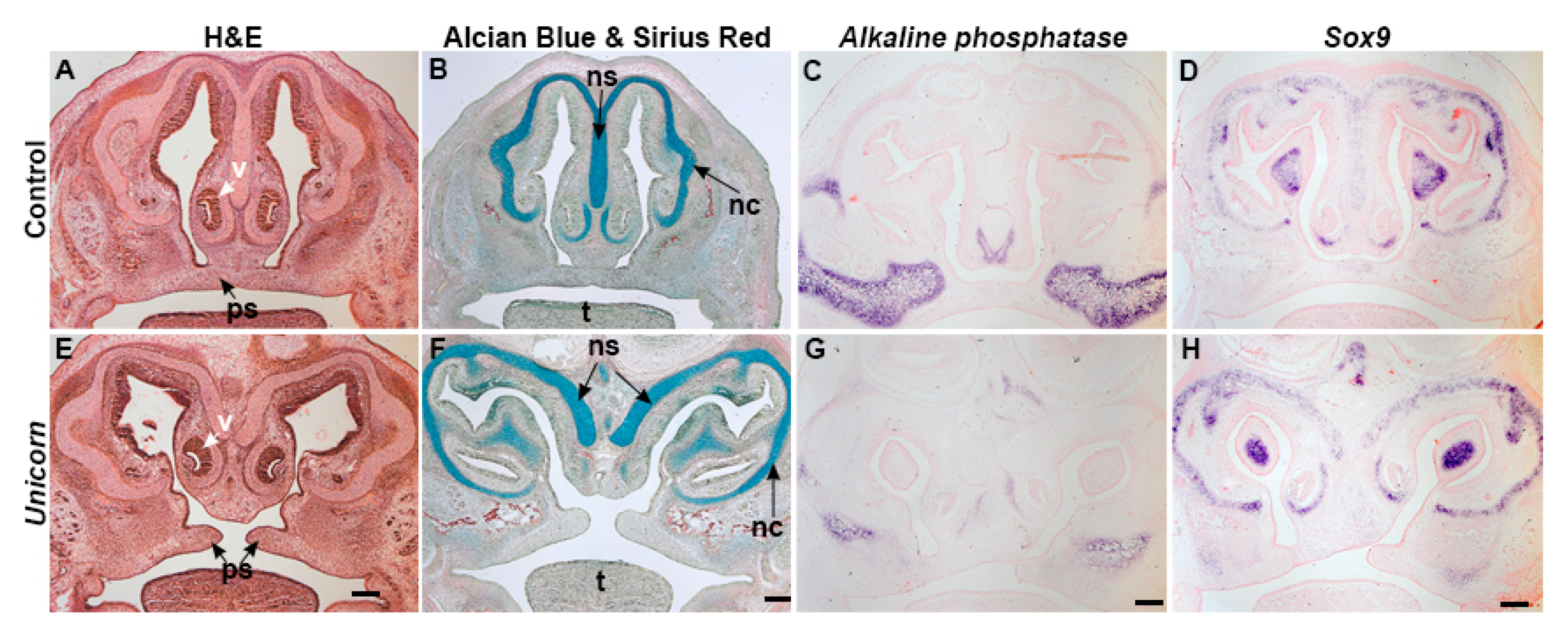
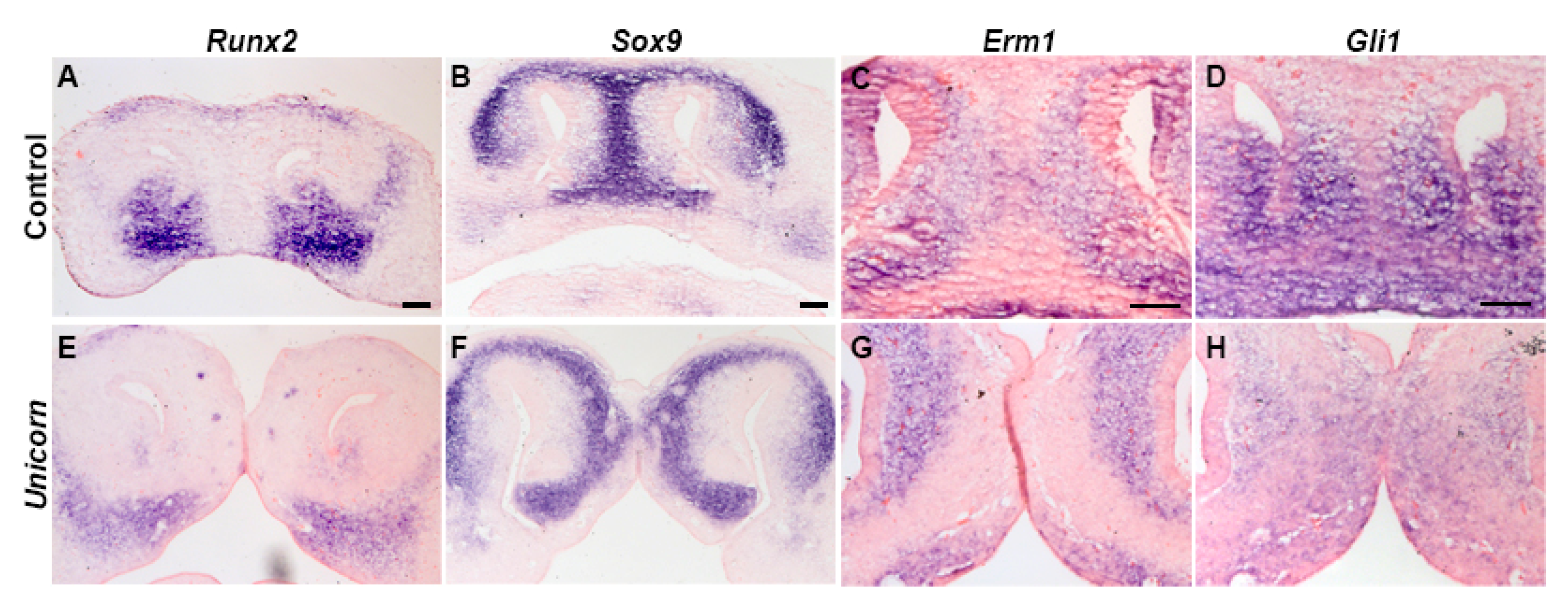
3.3. Unicorn Nasal Septum and Surrounding Tissues Differentiate Normally
3.4. Unicorn Patterning is Normal at E12.5
3.5. The Critical Period for Midfacial Clefting Is between E10.5 and E11.5
3.6. Unicorn Embryos Have Decreased Proliferation in Caudal MNPs
3.7. Unicorn Frontonasal Ectodermal Zone Is Shifted Laterally
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dixon, M.J.; Marazita, M.L.; Beaty, T.H.; Murray, J.C. Cleft lip and palate: Understanding genetic and environmental influences. Nat. Rev. Genet. 2011, 12, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolker, A.R.; Sailon, A.M.; Meara, J.G.; Holmes, A.D. Midline cleft lip and bifid nose deformity: Description, classification, and treatment. J. Craniofac. Surg. 2015, 26, 2304–2308. [Google Scholar] [CrossRef] [PubMed]
- Bush, J.O.; Jiang, R. Palatogenesis: Morphogenetic and molecular mechanisms of secondary palate development. Development 2012, 139, 231–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, R.; Bush, J.O.; Lidral, A.C. Development of the upper lip: Morphogenetic and molecular mechanisms. Dev. Dyn. 2006, 235, 1152–1166. [Google Scholar] [CrossRef] [PubMed]
- Szabo-Rogers, H.L.; Smithers, L.E.; Yakob, W.; Liu, K.J. New directions in craniofacial morphogenesis. Dev. Biol. 2010, 341, 84–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.; Marcucio, R.S.; Helms, J. A zone of frontonasal ectoderm regulates patterning adn growth in the face. Development 2003, 130, 1749–1758. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.; Marcucio, R.S. Unique organization of the frontonasal ectodermal zone in birds and mammals. Dev. Biol. 2009, 325, 200–210. [Google Scholar] [CrossRef] [Green Version]
- Szabo-Rogers, H.L.; Geetha-Loganathan, P.; Whiting, C.J.; Nimmagadda, S.; Fu, K.; Richman, J.M. Novel skeletogenic patterning roles for the olfactory pit. Development 2009, 136, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Helms, J.A.; Schneider, R.A. Cranial skeletal biology. Nature 2003, 423, 326–331. [Google Scholar] [CrossRef]
- Brugmann, S.A.; Allen, N.C.; James, A.W.; Mekonnen, Z.; Madan, E.; Helms, J.A. A primary cilia-dependent etiology for midline facial disorders. Hum. Mol. Genet. 2010, 19, 1577–1592. [Google Scholar] [CrossRef] [Green Version]
- Brugmann, S.A.; Cordero, D.R.; Helms, J.A. Craniofacial ciliopathies: A new classification for craniofacial disorders. Am. J. Med. Genet. A 2010, 152, 2995–3006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordero, D.; Marcucio, R.; Hu, D.; Gaffield, W.; Tapadia, M.; Helms, J.A. Temporal perturbations in sonic hedgehog signaling elicit the spectrum of holoprosencephaly phenotypes. J. Clin. Investig. 2004, 114, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Marcucio, R.S.; Cordero, D.R.; Hu, D.; Helms, J.A. Molecular interactions coordinating the development of the forebrain and face. Dev. Biol. 2005, 284, 48–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghyselinck, N.B.; Duester, G. Retinoic acid signaling pathways. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Hui, J.N.; Fu, K.K.; Richman, J.M. Control of retinoic acid synthesis and FGF expression in the nasal pit is required to pattern the craniofacial skeleton. Dev. Biol. 2004, 276, 313–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helms, J.A.; Kim, C.H.; Hu, D.; Minkoff, R.; Thaller, C.; Eichele, G. Sonic hedgehog participates in craniofacial morphogenesis and is down-regulated by teratogenic doses of retinoic acid. Dev. Biol. 1997, 187, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Niederreither, K.; Subbarayan, V.; Dolle, P.; Chambon, P. Embryonic retinoic acid synthesis is essential for early mouse post-implantation development. Nat. Genet. 1999, 21, 444–448. [Google Scholar] [CrossRef]
- Jaehning, J.A. The Paf1 complex: Platform or player in RNA polymerase II transcription? Biochim. Biophys. Acta 2010, 1799, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Van Oss, S.B.; Cucinotta, C.E.; Arndt, K.M. Emerging Insights into the Roles of the Paf1 Complex in Gene Regulation. Trends Biochem. Sci. 2017, 42, 788–798. [Google Scholar] [CrossRef]
- Akanuma, T.; Koshida, S.; Kawamura, A.; Kishimoto, Y.; Takada, S. Paf1 complex homologues are required for Notch-regulated transcription during somite segmentation. EMBO Rep. 2007, 8, 858–863. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, C.T.; Langenbacher, A.; Hsieh, M.; Chen, J.N. The PAF1 complex component Leo1 is essential for cardiac and neural crest development in zebrafish. Dev. Biol. 2010, 341, 167–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Leatherbury, L.; Rosenthal, J.; Yu, Q.; Pappas, M.; Wessels, A.; Lucas, J.; Siegfried, B.; Chatterjee, B.; Svenson, K. Cardiovascular phenotyping of fetal mice by noninvasive high-frequency ultrasound facilitates recovery of ENU-induced mutations causing congenital cardiac and extracardiac defects. Physiol. Genom. 2006, 24, 23–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Klena, N.T.; Gabriel, G.C.; Liu, X.; Kim, A.J.; Lemke, K.; Chen, Y.; Chatterjee, B.; Devine, W.; Damerla, R.R. Global genetic analysis in mice unveils central role for cilia in congenital heart disease. Nature 2015, 521, 520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo-Rogers, H.; Yakob, W.; Liu, K.J. Frontal Bone Insufficiency in Gsk3β Mutant Mice. PLoS ONE 2016, 11, e0149604. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Lantz, B.; Cusack, B.J.; Szabo-Rogers, H.L. Prickle1 regulates differentiation of frontal bone osteoblasts. Sci. Rep. 2018, 8, 18021. [Google Scholar] [CrossRef]
- Wan, Y.; Rogers, M.B.; Szabo-Rogers, H.L. A six-gene expression toolbox for the glands, epithelium and chondrocytes in the mouse nasal cavity. Gene Expr. Patterns Gep. 2017, 27, 46–55. [Google Scholar] [CrossRef]
- Ribes, V.; Wang, Z.; Dolle, P.; Niederreither, K. Retinaldehyde dehydrogenase 2 (RALDH2)-mediated retinoic acid synthesis regulates early mouse embryonic forebrain development by controlling FGF and sonic hedgehog signaling. Development 2006, 133, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Schmahl, J.; Raymond, C.S.; Soriano, P. PDGF signaling specificity is mediated through multiple immediate early genes. Nat. Genet. 2007, 39, 52–60. [Google Scholar] [CrossRef]
- Hendzel, M.J.; Wei, Y.; Mancini, M.A.; Van Hooser, A.; Ranalli, T.; Brinkley, B.; Bazett-Jones, D.P.; Allis, C.D. Mitosis-specific phosphorylation of histone H3 initiates primarily within pericentromeric heterochromatin during G2 and spreads in an ordered fashion coincident with mitotic chromosome condensation. Chromosoma 1997, 106, 348–360. [Google Scholar] [CrossRef]
- Juan, G.; Traganos, F.; James, W.M.; Ray, J.M.; Roberge, M.; Sauve, D.M.; Anderson, H.; Darzynkiewicz, Z. Histone H3 phosphorylation and expression of cyclins A and B1 measured in individual cells during their progression through G2 and mitosis. Cytom. J. Int. Soc. Anal. Cytol. 1998, 32, 71–77. [Google Scholar] [CrossRef]
- Cuervo, R.; Covarrubias, L. Death is the major fate of medial edge epithelial cells and the cause of basal lamina degradation during palatogenesis. Development 2004, 131, 15–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi, K.; Sato, N.; Uchiyama, Y. Apoptosis and heterophagy of medial edge epithelial cells of the secondary palatine shelves during fusion. Arch. Histol. Cytol. 1995, 58, 191–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, C.; Nakamura, N.; Okamoto, Y.; Osawa, M.; Shiota, K. Cytochemical identification of programmed cell death in the fusing fetal mouse palate by specific labelling of DNA fragmentation. Anat. Embryol. 1994, 190, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Martınez-Alvarez, C.; Tudela, C.; Perez-Miguelsanz, J.; O’kane, S.; Puerta, J.; Ferguson, M. Medial edge epithelial cell fate during palatal fusion. Dev. Biol. 2000, 220, 343–357. [Google Scholar] [CrossRef] [PubMed]
- Urata, M.M.; Kawamoto, H.K. Median clefts of the upper lip: A review and surgical management of a minor manifestation. J. Craniofac. Surg. 2003, 14, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Koh, K.S.; Do Yeon Kim, T.S.O. Clinical features and management of a median cleft lip. Arch. Plast. Surg. 2016, 43, 242. [Google Scholar] [CrossRef]
- Scott, J. The cartilage of the nasal septum. Br. Dent. J. 1953, 95, 37–40. [Google Scholar]
- Kvinnsland, S. Partial resection of the cartilaginous nasal septum in rats; its influence on growth. Angle Orthod. 1974, 44, 135–140. [Google Scholar]
- Sarnat, B.G.; Wexler, M.R. Growth of the face and jaws after resection of the septal cartilage in the rabbit. Dev. Dyn. 1966, 118, 755–767. [Google Scholar] [CrossRef]
- Nakamura, E.; Nguyen, M.T.; Mackem, S. Kinetics of tamoxifen-regulated Cre activity in mice using a cartilage-specific CreER(T) to assay temporal activity windows along the proximodistal limb skeleton. Dev. Dyn. 2006, 235, 2603–2612. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lantz, B.; White, C.; Liu, X.; Wan, Y.; Gabriel, G.; Lo, C.W.Y.; Szabo-Rogers, H.L. Finding the Unicorn, a New Mouse Model of Midfacial Clefting. Genes 2020, 11, 83. https://doi.org/10.3390/genes11010083
Lantz B, White C, Liu X, Wan Y, Gabriel G, Lo CWY, Szabo-Rogers HL. Finding the Unicorn, a New Mouse Model of Midfacial Clefting. Genes. 2020; 11(1):83. https://doi.org/10.3390/genes11010083
Chicago/Turabian StyleLantz, Brandi, Casey White, Xinyun Liu, Yong Wan, George Gabriel, Cecilia W. Y. Lo, and Heather L. Szabo-Rogers. 2020. "Finding the Unicorn, a New Mouse Model of Midfacial Clefting" Genes 11, no. 1: 83. https://doi.org/10.3390/genes11010083