A Single Nucleotide ADA Genetic Variant Is Associated to Central Inflammation and Clinical Presentation in MS: Implications for Cladribine Treatment

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. MS Patients
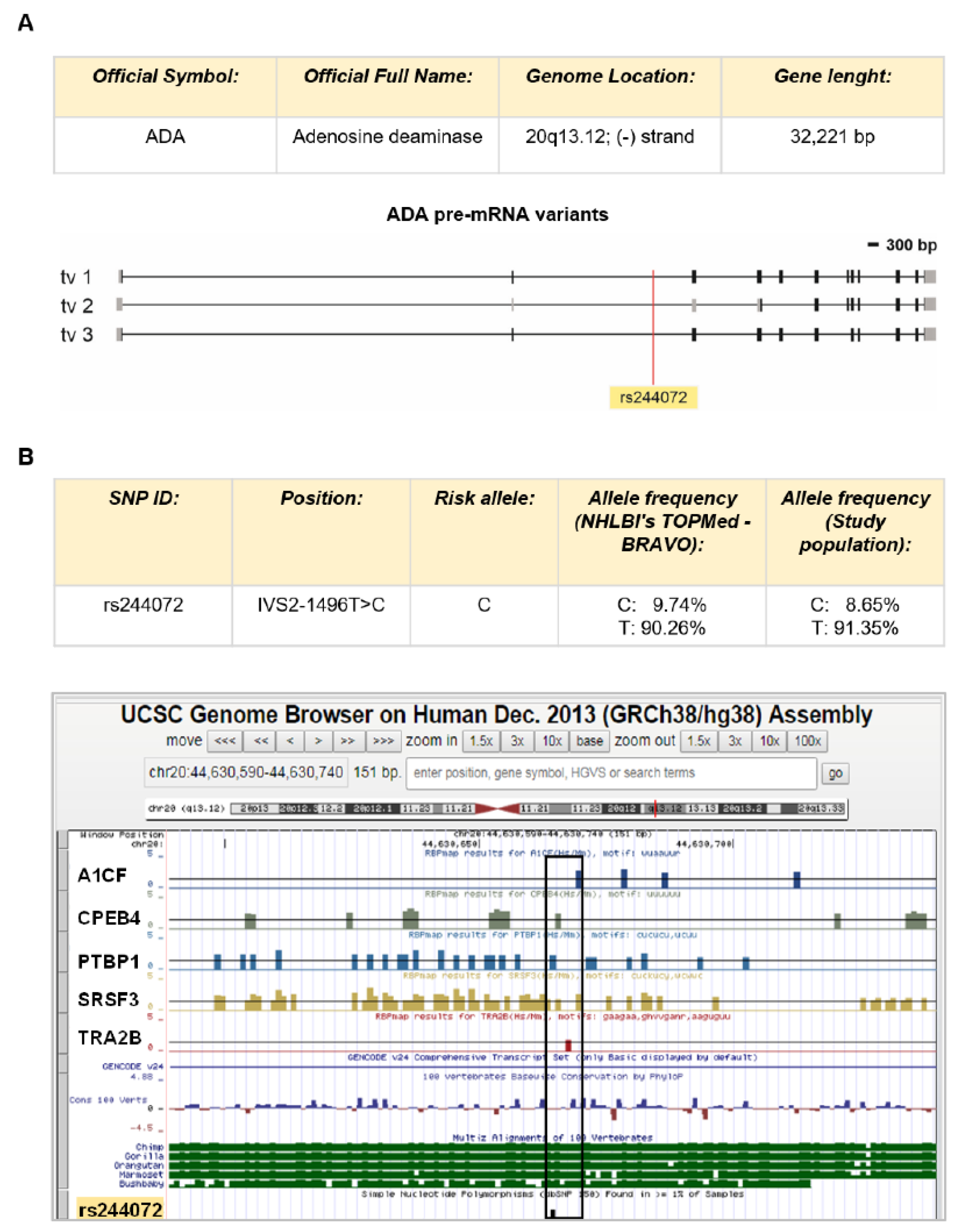
2.2. SNP rs244072 Analysis
2.3. CSF Collection and Analysis
2.4. Blood Samples Collection and WBC Count/Analysis
2.5. Statistical Analysis
3. Results
3.1. The ADA SNP rs244072 in Patients with MS
3.2. The ADA SNP rs244072 and Clinical Characteristics
3.3. The ADA SNP rs244072 and CNS Inflammation
3.4. ADA SNP rs244072 Peripheral Blood Lymphocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADA | Adenosine deaminase |
| CIS | Clinically isolated syndrome |
| CNS | Central nervous system |
| CSF | Cerebrospinal fluid |
| EAE | Experimental autoimmune encephalomyelitis |
| EDSS | Expanded disability status score |
| Gd | Gadolinium |
| Gd+ | Gadolinium-enhancing |
| IL | Interleukin |
| IQR | Interquartile range |
| LP | Lumbar puncture |
| MS | Multiple Sclerosis |
| NLR | Neutrophil/lymphocyte ratio |
| PP | Primary-progressive |
| PTBP1 | Polypyrimidine Tract Binding Protein 1 |
| RR | Relapsing-remitting |
| SP | Secondary-progressive |
| SRSF3 | Serine and arginine Rich Splicing Factor 3 |
| SNP | Single nucleotide polymorphism |
| SD | Standard deviation |
| TNF | Tumor necrosis factor |
| WBC | white blood cells |
| % | Frequency |
References
- Chitnis, T. The role of CD4 T cells in the pathogenesis of multiple sclerosis. Int. Rev. Neurobiol. 2007, 79, 43–72. [Google Scholar] [CrossRef] [PubMed]
- Arneth, B.M. Impact of B cells to the pathophysiology of multiple sclerosis. J. Neuroinflamm. 2019, 16, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandolesi, G.; Gentile, A.; Musella, A.; Fresegna, D.; De Vito, F.; Bullitta, S.; Sepman, H.; Marfia, G.A.; Centonze, D. Synaptopathy connects inflammation and neurodegeneration in multiple sclerosis. Nat. Rev. Neurol. 2015, 11, 711–724. [Google Scholar] [CrossRef] [PubMed]
- Gentile, A.; De Vito, F.; Fresegna, D.; Rizzo, F.R.; Bullitta, S.; Guadalupi, L.; Vanni, V.; Buttari, F.; Stampanoni Bassi, M.; Leuti, A.; et al. Peripheral T cells from multiple sclerosis patients trigger synaptotoxic alterations in central neurons. Neuropathol. Appl. Neurobiol. 2020, 46, 160–170. [Google Scholar] [CrossRef]
- Stampanoni Bassi, M.; Iezzi, E.; Landi, D.; Monteleone, F.; Gilio, L.; Simonelli, I.; Musella, A.; Mandolesi, G.; De Vito, F.; Furlan, R.; et al. Delayed treatment of MS is associated with high CSF levels of IL-6 and IL-8 and worse future disease course. J. Neurol. 2018, 265, 2540–2547. [Google Scholar] [CrossRef]
- Larochelle, C.; Alvarez, J.I.; Prat, A. How do immune cells overcome the blood-brain barrier in multiple sclerosis? FEBS Lett. 2011, 585, 3770–3780. [Google Scholar] [CrossRef] [Green Version]
- di Penta, A.; Moreno, B.; Reix, S.; Fernandez-Diez, B.; Villanueva, M.; Errea, O.; Escala, N.; Vandenbroeck, K.; Comella, J.X.; Villosladaet, P. Oxidative stress and proinflammatory cytokines contribute to demyelination and axonal damage in a cerebellar culture model of neuroinflammation. PLoS ONE 2013, 8, e54722. [Google Scholar] [CrossRef] [Green Version]
- Beutler, E. Cladribine (2-chlorodeoxyadenosine). Lancet 1992, 340, 952–956. [Google Scholar] [CrossRef]
- Laugel, B.; Borlat, F.; Galibert, L.; Vicari, A.; Weissert, R.; Chvatchko, Y.; Bruniquel, D. Cladribine inhibits cytokine secretion by T cells independently of deoxycytidine kinase activity. J. Neuroimmunol. 2011, 240–241, 52–57. [Google Scholar] [CrossRef]
- Dong, K.; Gao, Z.W.; Zhang, H.Z. The role of adenosinergic pathway in human autoimmune diseases. Immunol. Res. 2016, 64, 1133–1141. [Google Scholar] [CrossRef] [Green Version]
- Polachini, C.R.; Spanevello, R.M.; Casali, E.A.; Zanini, D.; Pereira, L.B.; Martins, C.C.; Baldissareli, J.; Cardoso, A.M.; Duarte, M.F.; da Costa, P.; et al. Alterations in the cholinesterase and adenosine deaminase activities and inflammation biomarker levels in patients with multiple sclerosis. Neuroscience 2014, 266, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Samuraki, M.; Sakai, K.; Odake, Y.; Yoshita, M.; Misaki, K.; Nakada, M.; Yamada, M. Multiple sclerosis showing elevation of adenosine deaminase levels in the cerebrospinal fluid. Mult. Scler. Relat. Disord. 2017, 13, 44–46. [Google Scholar] [CrossRef] [PubMed]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: 2010 Revisions to the McDonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UCSC Genome Browser (Santa Cruz, University of California). Available online: genome.ucsc.edu; genome-euro.ucsc.edu (accessed on 16 February 2018).
- RBPmap, Mapping Binding Sites of RNA Binding Proteins v 1.1. Available online: http://rbpmap.technion.ac.il (accessed on 16 February 2018).
- David Bioinformatics Resources 6.8, Laboratory of Human Retrovirology and Immunoinformatics (LHRI). Available online: https://david.ncifcrf.gov/content.jsp?file=about_us.html (accessed on 15 April 2018).
- PubMed. Available online: https://www.ncbi.nlm.nih.gov/pubmed/ (accessed on 31 May 2018).
- ten Dam, G.B.; Wieringa, B.; Poels, L.G. Alternative splicing of CD45 pre-mRNA is uniquely obedient to conditions in lymphoid cells. Biochim. Biophys. Acta 1999, 1446, 317–333. [Google Scholar] [CrossRef]
- La Porta, J.; Matus-Nicodemos, R.; Valentín-Acevedo, A.; Covey, L.R. The RNA-Binding Protein, Polypyrimidine Tract-Binding Protein 1 (PTBP1) Is a Key Regulator of CD4 T Cell Activation. PLoS ONE 2016, 11, e0158708. [Google Scholar] [CrossRef] [Green Version]
- Monzón-Casanova, E.; Screen, M.; Díaz-Muñoz, M.D.; Coulson, R.M.R.; Bell, S.E.; Lamers, G.; Solimena, M.; Smith, C.W.J.; Turner, M. The RNA-binding protein PTBP1 is necessary for B cell selection in germinal centers. Nat. Immunol. 2018, 19, 267–278. [Google Scholar] [CrossRef]
- Demirci, S.; Demirci, S.; Kutluhan, S.; Koyuncuoglu, H.R.; Yurekli, V.A. The clinical significance of the neutrophil-to-lymphocyte ratio in multiple sclerosis. Int. J. Neurosci. 2016, 126, 700–706. [Google Scholar] [CrossRef]
- Mayo, L.; Da Cunha, A.P.; Madi, A.; Beynon, V.; Yang, Z.; Alvarez, J.I.; Prat, A.; Sobel, R.A.; Kobzik, L.; Lassmann, H.; et al. IL-10-dependent Tr1 cells attenuate astrocyte activation and ameliorate chronic central nervous system inflammation. Brain 2016, 139, 1939–1957. [Google Scholar] [CrossRef] [Green Version]
- Stampanoni Bassi, M.; Iezzi, E.; Marfia, G.A.; Simonelli, I.; Musella, A.; Mandolesi, G.; Fresegna, D.; Pasqualetti, P.; Furlan, R.; Finardi, A.; et al. Platelet-derived growth factor predicts prolonged relapse-free period in multiple sclerosis. J. Neuroinflamm. 2018, 15, 108. [Google Scholar] [CrossRef]
- Centonze, D.; Muzio, L.; Rossi, S.; Cavasinni, F.; De Chiara, V.; Bergami, A.; Musella, A.; D’Amelio, M.; Cavallucci, V.; Martorana, A.; et al. Inflammation triggers synaptic alteration and degeneration in experimental autoimmune encephalomyelitis. J. Neurosci. 2009, 29, 3442–3452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, S.; Motta, C.; Studer, V.; Barbieri, F.; Buttari, F.; Bergami, A.; Sancesario, G.; Bernardini, S.; De Angelis, G.; Martino, G.; et al. Tumor necrosis factor is elevated in progressive multiple sclerosis and causes excitotoxic neurodegeneration. Mult. Scler. 2014, 20, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Szabo, M.C.; Butcher, E.C.; McIntyre, B.W.; Schall, T.J.; Bacon, K.B. RANTES stimulation of T lymphocyte adhesion and activation: Role for LFA-1 and ICAM-3. Eur. J. Immunol. 1997, 27, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.C.; Samanta, A.K.; Halder, J.B.; Hong, J.; Tejada-Simon, M.V.; Rivera, M.V.; Zhang, J.Z. Aberrant T cell migration toward RANTES and MIP-1 α in patients with multiple sclerosis. Overexpression of chemokine receptor CCR5. Brain 2000, 123, 1874–1882. [Google Scholar] [CrossRef]
- Iarlori, C.; Reale, M.; Lugaresi, A.; De Luca, G.; Bonanni, L.; Di Iorio, A.; Feliciani, C.; Conti, P.; Gambi, D. RANTES production and expression is reduced in relapsing-remitting multiple sclerosis patients treated with interferon-β-1b. J. Neuroimmunol. 2000, 107, 100–107. [Google Scholar] [CrossRef]
- Mori, F.; Nisticò, R.; Nicoletti, C.G.; Zagaglia, S.; Mandolesi, G.; Piccinin, S.; Martino, G.; Finardi, A.; Rossini, P.M.; Marfia, G.A.; et al. RANTES Correlates with Inflammatory Activity and Synaptic Excitability in Multiple Sclerosis. Mult. Scler. 2016, 22, 1405–1412. [Google Scholar] [CrossRef] [Green Version]
- van Veen, T.; Nielsen, J.; Berkhof, J.; Barkhof, F.; Kamphorst, W.; Bö, L.; Ravid, R.; Verweij, C.L.; Huitinga, I.; Polman, C.H.; et al. CCL5 and CCR5 genotypes modify clinical, radiological and pathological features of multiple sclerosis. J. Neuroimmunol. 2007, 190, 157–164. [Google Scholar] [CrossRef]
- Wiesemann, E.; Klatt, J.; Sönmez, D.; Heidenreich, F.; Windhagen, A. Glatiramer acetate (GA) induces IL-13/IL-5 secretion in naive T cells. J. Neuroimmunol. 2001, 119, 137–144. [Google Scholar] [CrossRef]
- van Boxel-Dezaire, A.H.; Hoff, S.C.; van Oosten, B.W.; Verweij, C.L.; Dräger, A.M.; Adèr, H.J.; van Houwelingen, J.C.; Barkhof, F.; Polman, C.H.; Nagelkerken, L. Decreased interleukin-10 and increased interleukin-12p40 mRNA are associated with disease activity and characterize different disease stages in multiple sclerosis. Ann. Neurol. 1999, 45, 695–703. [Google Scholar] [CrossRef]
- Martinez-Forero, I.; Garcia-Munoz, R.; Martinez-Pasamar, S.; Inoges, S.; Lopez-Diaz de Cerio, A.; Palacios, R.; Sepulcre, J.; Moreno, B.; Gonzalez, Z.; Fernandez-Diez, B.; et al. IL-10 suppressor activity and ex vivo Tr1 cell function are impaired in multiple sclerosis. Eur. J. Immunol. 2008, 38, 576–586. [Google Scholar] [CrossRef]
- Oliveira, I.B.N.; Gomes, R.S.; Gomides, L.F.; Dos Santos, J.C.; Diniz Carneiro, M.A.; Ribeiro-Dias, F.; Sisterolli Diniz, D. Interferon-β Treatment Differentially Alters TLR2 and TLR4-Dependent Cytokine Production in Multiple Sclerosis Patients. Neuroimmunomodulation 2019, 21, 1–7. [Google Scholar] [CrossRef]
- Wang, H.; Brown, J.; Garcia, C.A.; Tang, Y.; Benakanakere, M.R.; Greenway, T.; Alard, P.; Kinane, D.F.; Martin, M. The role of glycogen synthase kinase 3 in regulating IFN-β-mediated IL-10 production. J. Immunol. 2011, 186, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, G.; Soelberg Sorensen, P.; Cook, S.; Rammohan, K.W.; Rieckmann, P.; Comi, G.; Dangond, F.; Hicking, C.; Vermersch, P. Efficacy of Cladribine Tablets in high disease activity subgroups of patients with relapsing multiple sclerosis: A post hoc analysis of the CLARITY study. Mult. Scler. 2019, 25, 819–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartosik-Psujek, H.; Belniak, E.; Mitosek-Szewczyk, K.; Dobosz, B.; Stelmasiak, Z. Interleukin-8 and RANTES levels in patients with relapsing-remitting multiple sclerosis (RR-MS) treated with cladribine. Acta Neurol. Scand. 2004, 109, 390–392. [Google Scholar] [CrossRef] [PubMed]
- Freund, M.K.; Burch, K.S.; Shi, H.; Mancuso, N.; Kichaev, G.; Garske, K.M.; Pan, D.Z.; Miao, Z.; Mohlke, K.L.; Laakso, M.; et al. Phenotype-Specific Enrichment of Mendelian Disorder Genes near GWAS Regions across 62 Complex Traits. Am. J. Hum. Genet. 2018, 103, 535–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, M.R.; Tipney, H.; Painter, J.L.; Shen, J.; Nicoletti, P.; Shen, Y.; Floratos, A.; Sham, P.C.; Li, M.J.; Wang, J.; et al. The support of human genetic evidence for approved drug indications. Nat. Genet. 2015, 47, 856–860. [Google Scholar] [CrossRef]
- Floris, M.; Olla, S.; Schlessinger, D.; Cucca, F. Genetic-driven druggable target identification and validation. Trends Genet. 2018, 34, 558–570. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MS Patients n = 561 | ||
|---|---|---|
| CIS | N (%) | 91 (16.2) |
| RIS | N (%) | 17 (3) |
| RR-MS | N (%) | 404 (72) |
| SP/PP-MS | N (%) | 49 (8.8) |
| Sex, F | N (%) | 373 (66.5%) |
| Age, years | Mean, (SD) | 36.1 (10.63) |
| Disease duration, months | Median (IQR) | 8.17 (1.42–48.72) |
| EDSS at diagnosis | Median (IQR) | 2 (1–2.5) |
| Clinical activity at LP | N (%) | 215 (38.3) |
| Radiological activity at LP | N (%) | 238 (42.4) |
| CT/CC n = 92 (16.4%) | TT n = 469 (83.6%) | p Value | ||
|---|---|---|---|---|
| Sex, F | N (%) | 59 (64.1) | 314 (67) | 0.60 |
| Age, years | Mean, (SD) | 38.03 (11.99) | 36.09 (11.01) | 0.213 |
| MS phenotype | RR/CIS/RIS, n (%) | 85 (92.4) | 427 (91) | 0.676 |
| SP/PP, n (%) | 7 (7.6) | 42 (9) | ||
| Disease duration, months | Median (IQR) | 10.4 (1.62–48.17) | 7.9 (1.42–49.70) | 0.930 |
| EDSS | Median (IQR) | 2 (1–3) | 1.5 (1–2.5) | 0.011 |
| Clinical activity at LP | N (%) | 34 (37) | 181 (38.6) | 0.768 |
| Radiological activity at LP | N (%) | 42 (45.7) | 196 (41.8) | 0.493 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stampanoni Bassi, M.; Buttari, F.; Simonelli, I.; Gilio, L.; Furlan, R.; Finardi, A.; Marfia, G.A.; Visconti, A.; Paolillo, A.; Storto, M.; et al. A Single Nucleotide ADA Genetic Variant Is Associated to Central Inflammation and Clinical Presentation in MS: Implications for Cladribine Treatment. Genes 2020, 11, 1152. https://doi.org/10.3390/genes11101152
Stampanoni Bassi M, Buttari F, Simonelli I, Gilio L, Furlan R, Finardi A, Marfia GA, Visconti A, Paolillo A, Storto M, et al. A Single Nucleotide ADA Genetic Variant Is Associated to Central Inflammation and Clinical Presentation in MS: Implications for Cladribine Treatment. Genes. 2020; 11(10):1152. https://doi.org/10.3390/genes11101152
Chicago/Turabian StyleStampanoni Bassi, Mario, Fabio Buttari, Ilaria Simonelli, Luana Gilio, Roberto Furlan, Annamaria Finardi, Girolama Alessandra Marfia, Andrea Visconti, Andrea Paolillo, Marianna Storto, and et al. 2020. "A Single Nucleotide ADA Genetic Variant Is Associated to Central Inflammation and Clinical Presentation in MS: Implications for Cladribine Treatment" Genes 11, no. 10: 1152. https://doi.org/10.3390/genes11101152