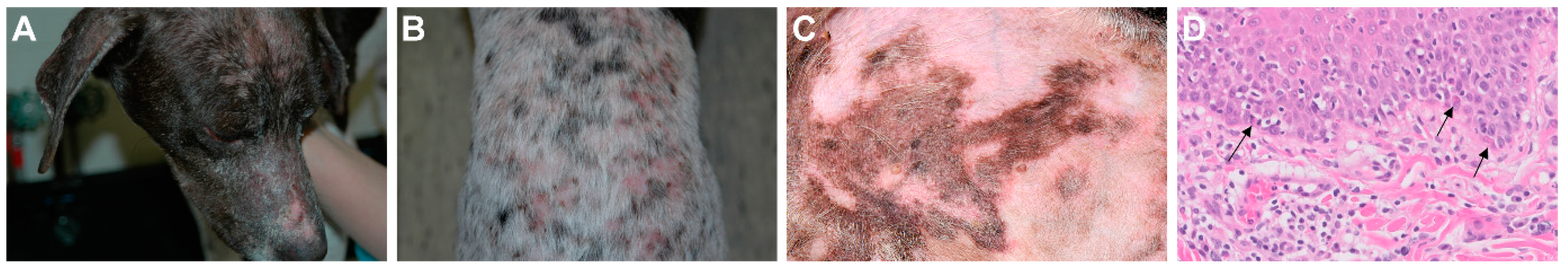
A Missense Variant Affecting the C-Terminal Tail of UNC93B1 in Dogs with Exfoliative Cutaneous Lupus Erythematosus (ECLE)

 ,
,  , ,
, ,  , , ,
, , ,  , ,
, , Highlights
- We identify a missense variant in UNC93B1 as the cause of a monogenic form of lupus erythematosus in dogs.
- Our genetic results in dogs complement two recent functional studies on the role of Unc93b1 in mice.
- The canine disease has been known for decades and hundreds of affected dogs have died from it.
- Similar UNC93B1 variants might also contribute to autoimmune disease in human patients.
- Affected dogs are an excellent model for studying the disease and potential treatment options.
- Our findings enable genetic testing to avoid the unintentional breeding of further affected dogs.
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animal Selection
2.3. DNA Extraction and SNV Genotyping
2.4. GWAS and Autozygosity Mapping
2.5. Whole Genome Sequencing of Two Affected German Shorthaired Pointers
2.6. Variant Calling
2.7. Gene Analysis
2.8. Sanger Sequencing
3. Results
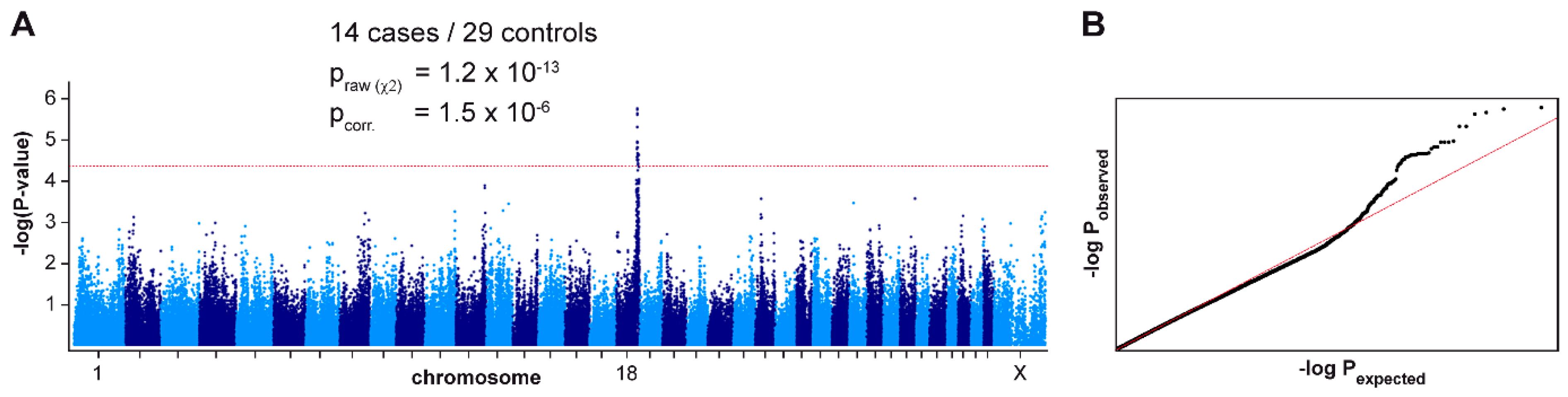
3.1. Mapping of the ECLE Locus
3.2. Identification of a Candidate Causative Variant
3.3. Genotype Phenotype Association of the UNC93B1:p.Pro480Thr Variant
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sontheimer, R.D. The lexicon of cutaneous lupus erythematosus—A review and personal perspective on the nomenclature and classification of the cutaneous manifestations of lupus erythematosus. Lupus 1997, 6, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Rothfield, N.; Sontheimer, R.D.; Bernstein, M. Lupus erythematosus: Systemic and cutaneous manifestations. Clin. Dermatol. 2006, 24, 348–362. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, A.; Landmann, A. The classification and diagnosis of cutaneous lupus erythematosus. J. Autoimmun. 2014, 48–49, 14–19. [Google Scholar] [CrossRef]
- Olivry, T.; Linder, K.E.; Banovic, F. Cutaneous lupus erythematosus in dogs: A comprehensive review. BMC Vet. Res. 2018, 14, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durosaro, O.; Davis, M.D.; Reed, K.B.; Rohlinger, A.L. Incidence of cutaneous lupus erythematosus, 1965-2005: A population-based study. Arch. Dermatol. 2009, 145, 249–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grönhagen, C.M.; Fored, C.M.; Granath, F.; Nyberg, F. Cutaneous lupus erythematosus and the association with systemic lupus erythematosus: A population-based cohort of 1088 patients in Sweden. Br. J. Dermatol. 2011, 164, 1335–1341. [Google Scholar] [CrossRef] [PubMed]
- Biazar, C.; Sigges, J.; Patsinakidis, N.; Ruland, V.; Amler, S.; Bonsmann, G.; Kuhn, A.; EUSCLE co-authors. Cutaneous lupus erythematosus: First multicenter database analysis of 1002 patients from the European Society of Cutaneous Lupus Erythematosus (EUSCLE). Autoimmun. Rev. 2013, 12, 444–454. [Google Scholar] [CrossRef]
- Jarukitsopa, S.; Hoganson, D.D.; Crowson, C.S.; Sokumbi, O.; Davis, M.D.; Michet, C.J., Jr.; Matteson, E.L.; Maradit Kremers, H.; Chowdhary, V.R. Epidemiology of systemic lupus erythematosus and cutaneous lupus erythematosus in a predominantly white population in the United States. Arthritis Care Res. (Hoboken) 2015, 67, 817–828. [Google Scholar] [CrossRef]
- Chong, B.F.; Song, J.; Olsen, N.J. Determining risk factors for developing systemic lupus erythematosus in patients with discoid lupus erythematosus. Br. J. Dermatol. 2012, 166, 29–35. [Google Scholar] [CrossRef]
- Hersh, A.O.; Arkin, L.M.; Prahalad, S. Immunogenetics of cutaneous lupus erythematosus. Curr. Opin. Pediatr. 2016, 28, 470–475. [Google Scholar] [CrossRef]
- Wenzel, J. Cutaneous lupus erythematosus: New insights into pathogenesis and therapeutic strategies. Nat. Rev. Rheumatol. 2019, 15, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Rice, G.; Newman, W.G.; Dean, J.; Patrick, T.; Parmar, R.; Flintoff, K.; Robins, P.; Harvey, S.; Hollis, T.; O’Hara, A.; et al. Heterozygous mutations in TREX1 cause familial chilblain lupus and dominant Aicardi-Goutieres syndrome. Am. J. Hum. Genet. 2007, 80, 811–815. [Google Scholar] [CrossRef] [PubMed]
- Peschke, K.; Friebe, F.; Zimmermann, N.; Wahlicht, T.; Schumann, T.; Achleitner, M.; Berndt, N.; Luksch, H.; Behrendt, R.; Lee-Kirsch, M.A.; et al. Deregulated type I IFN response in TREX1-associated familial chilblain lupus. J. Investig. Dermatol. 2014, 134, 1456–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasutomo, K.; Horiuchi, T.; Kagami, S.; Tsukamoto, H.; Hashimura, C.; Urushihara, M.; Kuroda, Y. Mutation of DNASE1 in people with systemic lupus erythematosus. Nat. Genet. 2001, 28, 313–314. [Google Scholar] [CrossRef] [PubMed]
- Napirei, M.; Karsunky, H.; Zevnik, B.; Stephan, H.; Mannherz, H.G.; Möröy, T. Features of systemic lupus erythematosus in Dnase1-deficient mice. Nat. Genet. 2000, 25, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Vroom, M.W.; Theaker, A.J.; Rest, J.R.; White, S.D. Case report: Lupoid dermatosis in 5 German short-hair pointer. Vet. Dermatol. 1995, 6, 93–98. [Google Scholar] [CrossRef]
- Bryden, S.L.; White, S.D.; Dunston, S.M.; Burrows, A.K.; Olivry, T. Clinical, histopathological and immunological characteristics of exfoliative cutaneous lupus erythematosus in 25 German short-haired pointers. Vet. Dermatol. 2005, 16, 239–252. [Google Scholar] [CrossRef]
- Mauldin, E.A.; Morris, D.O.; Brown, D.C.; Casal, M.L. Exfoliative cutaneous lupus erythematosus in German shorthaired pointer dogs: Disease development, progression and evaluation of three immunomodulatory drugs (ciclosporin, hydroxychloroquine, and adalimumab) in a controlled environment. Vet. Dermatol. 2010, 21, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Ferrigno, A.; Hoover, K.; Blubaugh, A.; Rissi, D.; Banovic, F. Treatment of exfoliative cutaneous lupus erythematosus in a German shorthaired pointer dog with mycophenolate mofetil. Vet. Dermatol. 2019, 30, 350. [Google Scholar] [CrossRef]
- Wang, P.; Zangerl, B.; Werner, P.; Mauldin, E.A.; Casal, M.L. Familial cutaneous lupus erythematosus (CLE) in the German shorthaired pointer maps to CFA18, a canine orthologue to human CLE. Immunogenetics 2010, 63, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Stephens, M. Genome-wide efficient mixed-model analysis for association studies. Nat. Genet. 2012, 44, 821–824. [Google Scholar] [CrossRef] [Green Version]
- Qqman: Q-Q and Manhattan Plots for GWAS Data. R Package Version 0.1.4. Available online: https://CRAN.R-project.org/package=qqman (accessed on 5 December 2019).
- Jagannathan, V.; Drögemüller, C.; Leeb, T.; Dog Biomedical Variant Database Consortium (DBVDC). A comprehensive biomedical variant catalogue based on whole genome sequences of 582 dogs and eight wolves. Anim. Genet. 2019, 50, 695–704. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Bai, B.; Zhao, W.M.; Tang, B.X.; Wang, Y.Q.; Wang, L.; Zhang, Z.; Yang, H.C.; Liu, Y.H.; Zhu, J.W.; Irwin, D.M.; et al. DoGSD: The dog and wolf genome SNP database. Nucleic Acids Res. 2015, 43, 777–783. [Google Scholar] [CrossRef] [Green Version]
- Plassais, J.; Kim, J.; Davis, B.W.; Karyadi, D.M.; Hogan, A.N.; Harris, A.C.; Decker, B.; Parker, H.G.; Ostrander, E.A. Whole genome sequencing of canids reveals genomic regions under selection and variants influencing morphology. Nat. Commun. 2019, 10, 1489. [Google Scholar] [CrossRef]
- The Welcome Trust Case Control Consortium. Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls. Nature 2007, 447, 661–678. [Google Scholar] [CrossRef] [Green Version]
- Majer, O.; Liu, B.; Kreuk, L.S.M.; Krogan, N.; Barton, G.M. UNC93B1 recruits syntenin-1 to dampen TLR7 signaling and prevent autoimmunity. Nature 2019, 575, 366–370. [Google Scholar] [CrossRef]
- Kashuba, V.I.; Protopopov, A.I.; Kvasha, S.M.; Gizatullin, R.Z.; Wahlestedt, C.; Kisselev, L.L.; Klein, G.; Zabarovsky, E.R. hUNC93B1: A novel human gene representing a new gene family and encoding an unc-93-like protein. Gene 2002, 283, 209–217. [Google Scholar] [CrossRef]
- Kim, Y.-M.; Brinkmann, M.M.; Paquet, M.-E.; Ploegh, H.L. UNC93B1 delivers nucleotide-sensing toll-like receptors to endolysosomes. Nature 2008, 452, 234–238. [Google Scholar] [CrossRef]
- Saitoh, S.; Miyake, K. Regulatory molecules required for nucleotide-sensing Toll-like receptors. Immunol. Rev. 2009, 227, 32–43. [Google Scholar] [CrossRef]
- Majer, O.; Liu, B.; Woo, B.J.; Kreuk, L.S.M.; Van Dis, E.; Barton, G.M. Release from UNC93B1 reinforces the compartmentalized activation of select TLRs. Nature 2019, 575, 371–374. [Google Scholar] [CrossRef]
- Fukui, R.; Saitoh, S.; Kanno, A.; Onji, M.; Shibata, T.; Ito, A.; Onji, M.; Matsumoto, M.; Akira, S.; Yoshida, N.; et al. Unc93B1 restricts systemic lethal inflammation by orchestrating Toll-like receptor 7 and 9 trafficking. Immunity 2011, 35, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Casrouge, A.; Zhang, S.Y.; Eidenschenk, C.; Jouanguy, E.; Puel, A.; Yang, K.; Alcais, A.; Picard, C.; Mahfoufi, N.; Nicolas, N.; et al. Herpes simplex virus encephalitis in human UNC-93B deficiency. Science 2006, 314, 308–312. [Google Scholar] [CrossRef]
- Tabeta, K.; Hoebe, K.; Janssen, E.M.; Du, X.; Georgel, P.; Crozat, K.; Mudd, S.; Mann, N.; Sovath, S.; Goode, J.; et al. The Unc93b1 mutation 3d disrupts exogenous antigen presentation and signaling via Toll-like receptors 3, 7 and 9. Nat. Immunol. 2006, 7, 156–164. [Google Scholar] [CrossRef]
- Deane, J.A.; Pisitkun, P.; Barrett, R.S.; Feigenbaum, L.; Town, T.; Ward, J.M.; Flavell, R.A.; Bolland, S. Control of toll-like receptor 7 expression is essential to restrict autoimmunity and dendritic cell proliferation. Immunity 2007, 27, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Pisitkun, P.; Deane, J.A.; Difilippantonio, M.J.; Tarasenko, T.; Satterthwaite, A.B.; Bolland, S. Autoreactive B cell responses to RNA-related antigens due to TLR7 gene duplication. Science 2006, 312, 1669–1672. [Google Scholar] [CrossRef]
- Subramanian, S.; Tus, K.; Li, Q.Z.; Wang, A.; Tian, X.H.; Zhou, J.; Liang, C.; Bartov, G.; McDaniel, L.D.; Zhou, X.J.; et al. A Tlr7 translocation accelerates systemic autoimmunity in murine lupus. Proc. Natl. Acad. Sci. USA 2006, 103, 9970–9975. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Filtering Step | Variants |
|---|---|
| Shared homozygous variants in whole genome | 1,420,602 |
| Private homozygous variants (absent from 654 control genomes) in whole genome | 25 |
| Shared homozygous variants in 493 kb critical interval | 851 |
| Private variants (absent from 654 control genomes) in critical interval | 2 |
| Protein changing private variants in critical interval | 1 |
| ECLE Phenotype | Breed | C/C | C/A | A/A |
|---|---|---|---|---|
| Affected | German Shorthaired Pointer (n = 24) | 1 | – | 23 |
| Control | German Shorthaired Pointer (n = 520) | 457 | 63 | – |
| Control | German Longhaired Pointer (n = 52) | 50 | 2 | – |
| Control | German Wirehaired Pointer (n = 210) | 210 | – | – |
| Control | Braque du Bourbonnais (n = 7) | 7 | – | – |
| Affected | Vizsla (n = 1) | 1 | ||
| Control | Vizsla (n = 56) | 51 | 5 | – |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leeb, T.; Leuthard, F.; Jagannathan, V.; Kiener, S.; Letko, A.; Roosje, P.; Welle, M.M.; Gailbreath, K.L.; Cannon, A.; Linek, M.; et al. A Missense Variant Affecting the C-Terminal Tail of UNC93B1 in Dogs with Exfoliative Cutaneous Lupus Erythematosus (ECLE). Genes 2020, 11, 159. https://doi.org/10.3390/genes11020159
Leeb T, Leuthard F, Jagannathan V, Kiener S, Letko A, Roosje P, Welle MM, Gailbreath KL, Cannon A, Linek M, et al. A Missense Variant Affecting the C-Terminal Tail of UNC93B1 in Dogs with Exfoliative Cutaneous Lupus Erythematosus (ECLE). Genes. 2020; 11(2):159. https://doi.org/10.3390/genes11020159
Chicago/Turabian StyleLeeb, Tosso, Fabienne Leuthard, Vidhya Jagannathan, Sarah Kiener, Anna Letko, Petra Roosje, Monika M. Welle, Katherine L. Gailbreath, Andrea Cannon, Monika Linek, and et al. 2020. "A Missense Variant Affecting the C-Terminal Tail of UNC93B1 in Dogs with Exfoliative Cutaneous Lupus Erythematosus (ECLE)" Genes 11, no. 2: 159. https://doi.org/10.3390/genes11020159
APA StyleLeeb, T., Leuthard, F., Jagannathan, V., Kiener, S., Letko, A., Roosje, P., Welle, M. M., Gailbreath, K. L., Cannon, A., Linek, M., Banovic, F., Olivry, T., White, S. D., Batcher, K., Bannasch, D., Minor, K. M., Mickelson, J. R., Hytönen, M. K., Lohi, H., ... Casal, M. L. (2020). A Missense Variant Affecting the C-Terminal Tail of UNC93B1 in Dogs with Exfoliative Cutaneous Lupus Erythematosus (ECLE). Genes, 11(2), 159. https://doi.org/10.3390/genes11020159