Clinical Ketosis-Associated Alteration of Gene Expression in Holstein Cows

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
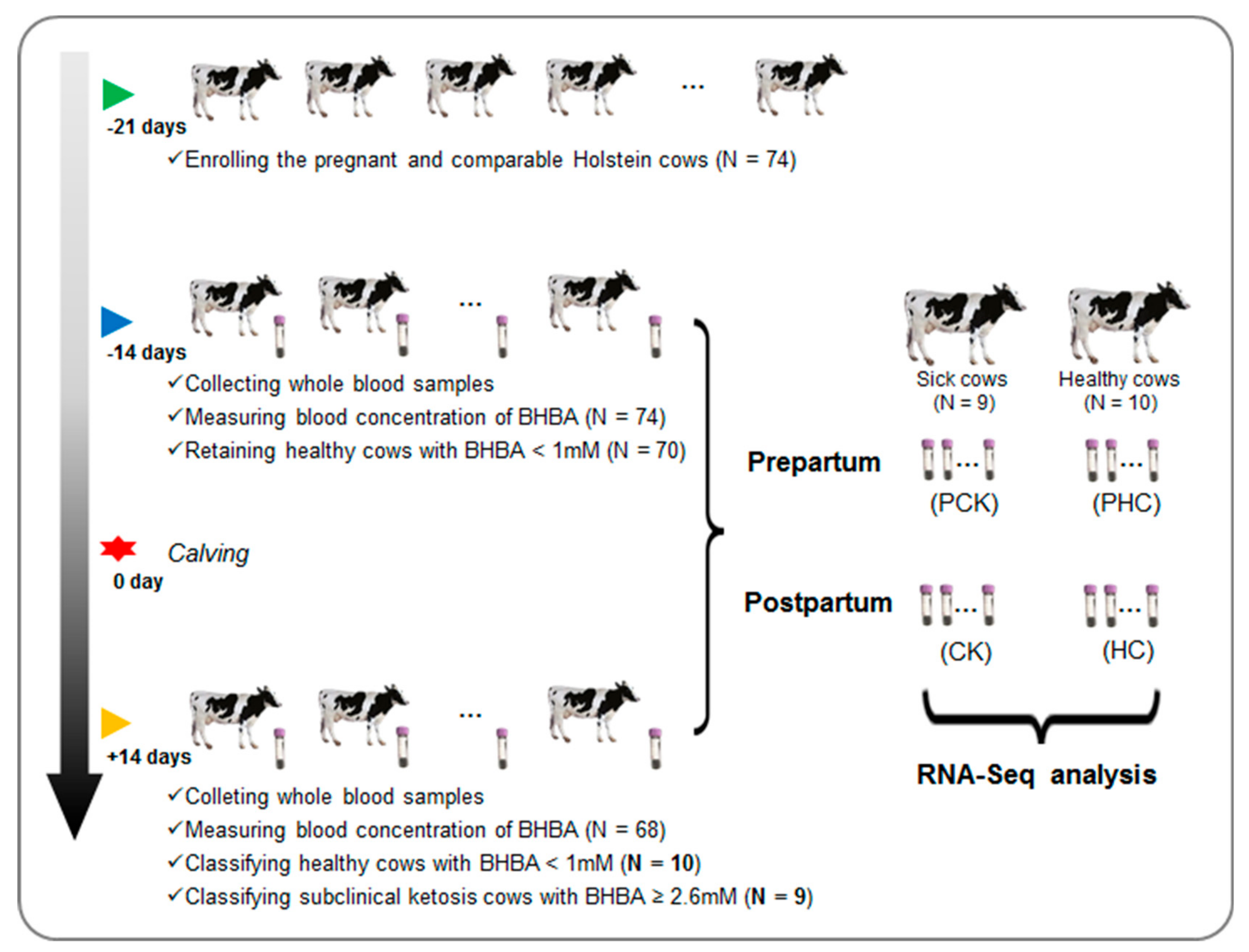
2.1. Animals and Ketosis Dagnosis
2.2. Collection of Blood Samples and RNA Extraction
2.3. Library Preparation and Sequencing
2.4. Reads Mapping and Quantification of Gene Expression
2.5. Differentially Expressed Genes and Functional Enrichment
2.6. Validation of RNA-Seq Data by qPCR
3. Results
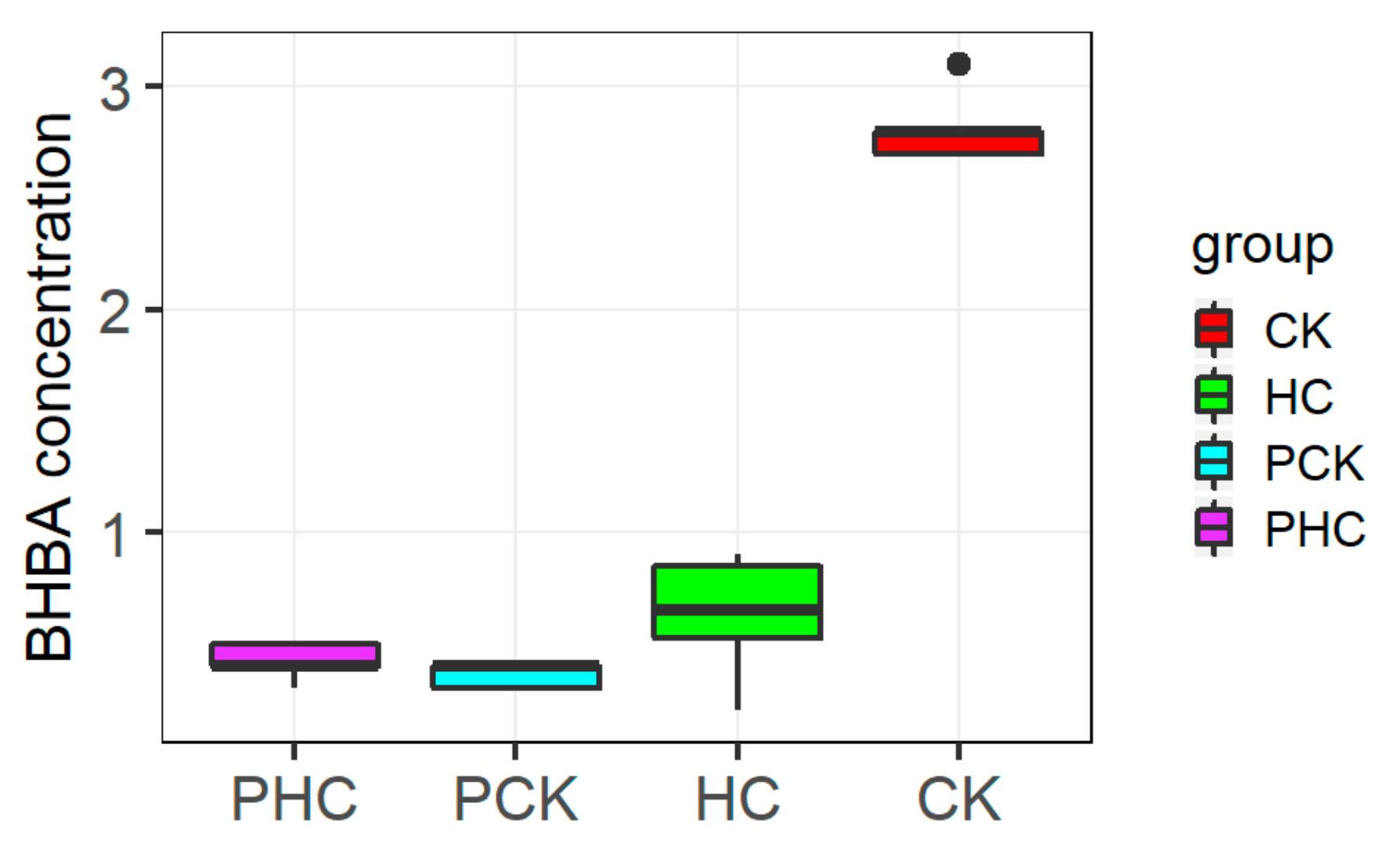
3.1. BHBA Parameters and Ketosis Diagnosis
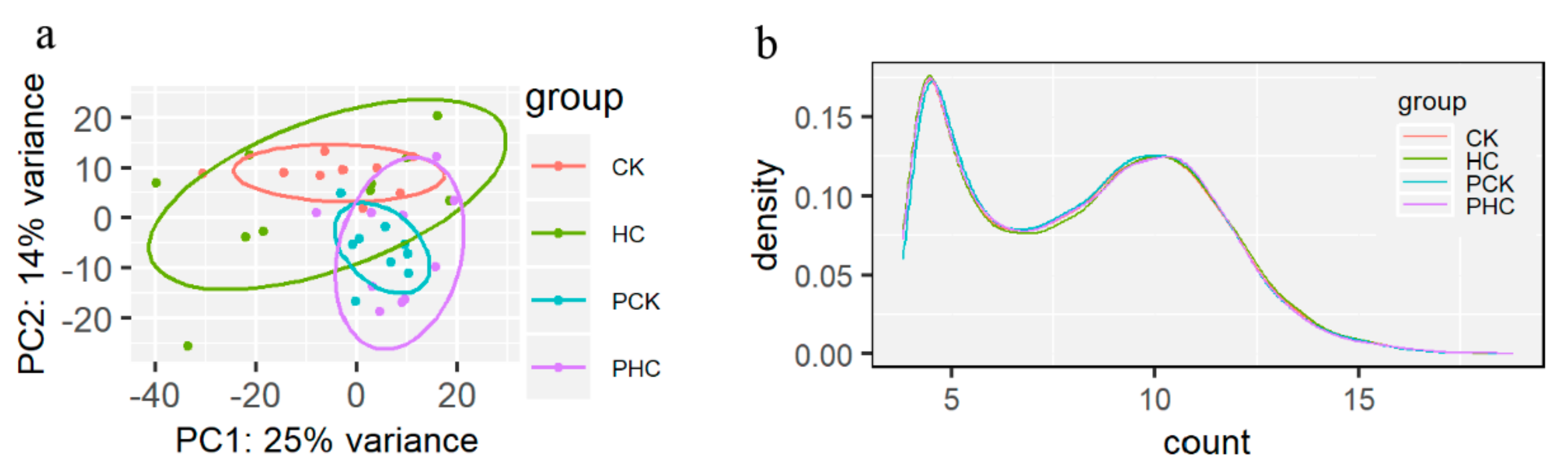
3.2. Gene Expression and Cluster Analysis
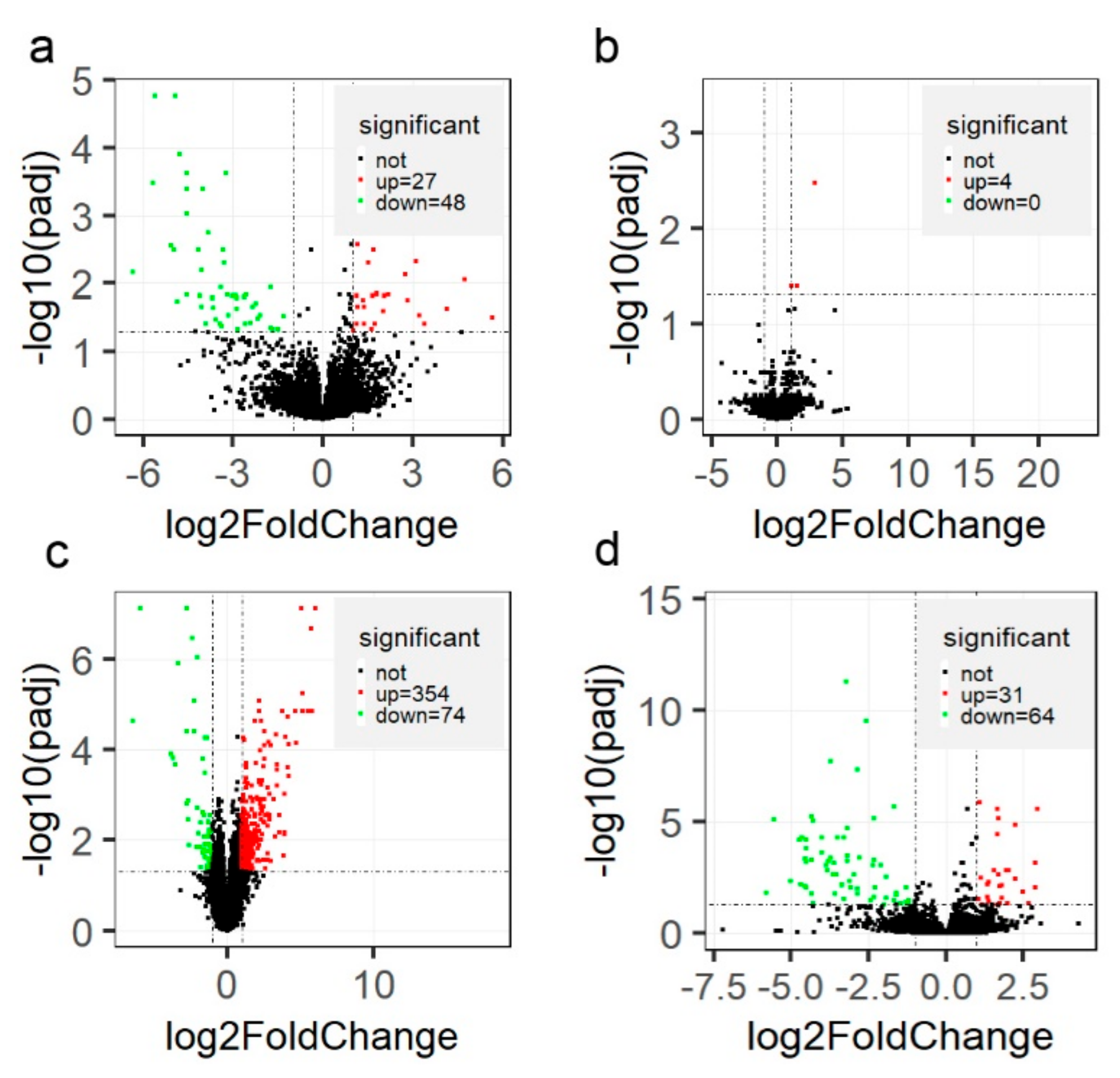
3.3. Differential Expression of Genes and Validation
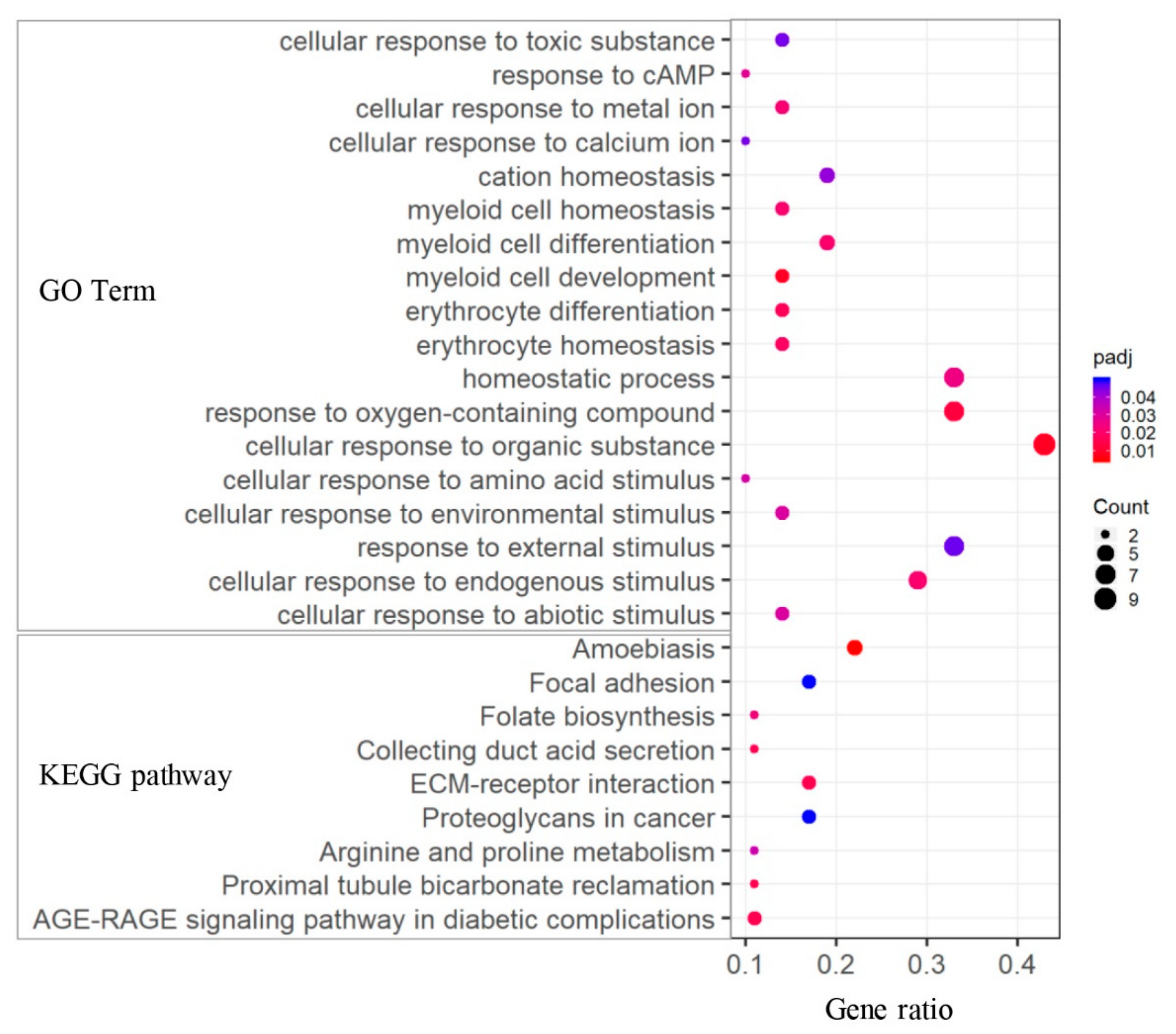
3.4. Functional Enrichment Analysis of DEGs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Drackley, J.K. Biology of dairy cows during the transition period: The final frontier? J. Dairy Sci. 1999, 82, 2259–2273. [Google Scholar] [CrossRef]
- Li, C.; Dai, S.; Lu, J.; Zhao, B.; Wang, J.; Li, P.; Wu, Z.; Mu, Y.; Feng, C.; Dong, Q. Methylglyoxal: A newly detected and potentially harmful metabolite in the blood of ketotic dairy cows. J. Dairy Sci. 2018, 101, 8513–8523. [Google Scholar] [CrossRef]
- McArt, J.A.A.; Nydam, D.V.; Oetzel, G.R. Epidemiology of subclinical ketosis in early lactation dairy cattle. J. Dairy Sci. 2012, 95, 5056–5066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffield, T. Subclinical Ketosis in Lactating Dairy Cattle. Vet. Clin. N. Am. Food Anim. Pract. 2000, 16, 231–253. [Google Scholar] [CrossRef]
- Suthar, V.S.; Canelas-Raposo, J.; Deniz, A.; Heuwieser, W. Prevalence of subclinical ketosis and relationships with postpartum diseases in European dairy cows. J. Dairy Sci. 2013, 96, 2925–2938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raboisson, D.; Mounié, M.; Maigné, E. Diseases, reproductive performance, and changes in milk production associated with subclinical ketosis in dairy cows: A meta-analysis and review. J. Dairy Sci. 2014, 97, 7547–7563. [Google Scholar] [CrossRef] [PubMed]
- Herdt, T.H. Ruminant adaptation to negative energy balance: Influences on the etiology of ketosis and fatty liver. Vet. Clin. N. Am. Food Anim. Pract. 2000, 16, 215–230. [Google Scholar] [CrossRef]
- Enjalbert, F.; Nicot, M.C.; Bayourthe, C.; Moncoulon, R. Ketone bodies in milk and blood of dairy cows: Relationship between concentrations and utilization for detection of subclinical ketosis. J. Dairy Sci. 2001, 84, 583–589. [Google Scholar] [CrossRef]
- Xia, C.; Wang, Z.; Xu, C.; Zhang, H.Y. Concentrations of plasma metabolites, hormones, and mRNA abundance of adipose leptin and hormone-sensitive lipase in ketotic and nonketotic dairy cows. J. Vet. Intern. Med. 2012, 26, 415–417. [Google Scholar] [CrossRef]
- Carrier, J.; Stewart, S.; Godden, S.; Fetrow, J.; Rapnicki, P. Evaluation and use of three cowside tests for detection of subclinical ketosis in early postpartum cows. J. Dairy Sci. 2004, 87, 3725–3735. [Google Scholar] [CrossRef] [Green Version]
- Oetzel, G.R. Herd-Level Ketosis-Diagnosis and Risk Factors, in American Association of Bovine Practitioners. In Proceedings of the 40th Annual Conference, Vancouver, BC, Canada, 19 September 2007. [Google Scholar]
- Jorjong, S.; van Knegsel, A.T.M.; Verwaeren, J.; Bruckmaier, R.M.; De Baets, B.; Kemp, B.; Fievez, V. Milk fatty acids as possible biomarkers to diagnose hyperketonemia in early lactation. J. Dairy Sci. 2015, 98, 5211–5221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhu, X.; She, G.; Kong, Y.; Guo, Y.; Wang, Z.; Liu, G.; Zhao, B. Serum hepatokines in dairy cows: Periparturient variation and changes in energy-related metabolic disorders. BMC Vet. Res. 2018, 14, 236. [Google Scholar] [CrossRef] [PubMed]
- Abuajamieh, M.; Kvidera, S.K.; Fernandez, M.V.S.; Nayeri, A.; Upah, N.C.; Nolan, E.A.; Lei, S.M.; DeFrain, J.M.; Green, H.B.; Schoenberg, K.M.; et al. Inflammatory biomarkers are associated with ketosis in periparturient Holstein cows. Res. Vet. Sci. 2016, 109, 81–85. [Google Scholar] [CrossRef]
- Hailemariam, D.; Mandal, R.; Saleem, F.; Dunn, S.M.; Wishart, D.S.; Ametaj, B.N. Identification of predictive biomarkers of disease state in transition dairy cows. J. Dairy Sci. 2014, 97, 2680–2693. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Dervishi, E.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Metallotyping of ketotic dairy cows reveals major alterations preceding, associating, and following the disease occurrence. Metabolomics 2017, 13, 97. [Google Scholar] [CrossRef]
- Fan, Z.; Shu, S.; Xu, C.; Xiao, X.; Wang, G.; Bai, Y.; Xia, C.; Wu, L.; Zhang, H.; Xu, C.; et al. Protein profiling of plasma proteins in dairy cows with subclinical hypocalcaemia. Ir. Vet. J. 2017, 70, 3. [Google Scholar] [CrossRef] [Green Version]
- Marczuk, J.; Brodzki, P.; Brodzki, A. The concentration of free amino acids in blood serum of dairy cows with primary ketosis. Pol. J. Vet. Sci. 2018, 21, 149–156. [Google Scholar]
- Parker Gaddis, K.L.; Cole, J.B.; Clay, J.S.; Maltecca, C. Genomic selection for producer-recorded health event data in US dairy cattle. J. Dairy Sci. 2014, 97, 3190–3199. [Google Scholar] [CrossRef]
- Parker Gaddis, K.L.; Megonigal, J.H.; Clay, J.S.; Wolfe, C.W. Genome-wide association study for ketosis in US Jerseys using producer-recorded data. J. Dairy Sci. 2018, 101, 413–424. [Google Scholar] [CrossRef]
- Srikanth, K.; Kwon, A.; Lee, E.; Chung, H. Characterization of genes and pathways that respond to heat stress in Holstein calves through transcriptome analysis. Cell Stress Chaperones 2017, 22, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Dervishi, E.; Zhang, G.; Dunn, S.M.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. GC–MS Metabolomics Identifies Metabolite Alterations That Precede Subclinical Mastitis in the Blood of Transition Dairy Cows. J. Proteome Res. 2017, 16, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Zhang, H.; Guo, D.; Sun, A.; Wang, H. Shotgun proteomic analysis of plasma from dairy cattle suffering from footrot: Characterization of potential disease-associated factors. PLoS ONE 2013, 8, e55973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humer, E.; Khol-Parisini, A.; Metzler-Zebeli, B.U.; Gruber, L.; Zebeli, Q. Alterations of the Lipid Metabolome in Dairy Cows Experiencing Excessive Lipolysis Early Postpartum. PLoS ONE 2016, 11, e0158633. [Google Scholar] [CrossRef] [Green Version]
- Mach, N.; Gao, Y.; Lemonnier, G.; Lecardonnel, J.; Oswald, I.P.; Estelle, J.; Rogel-Gaillard, C. The peripheral blood transcriptome reflects variations in immunity traits in swine: Towards the identification of biomarkers. BMC Genom. 2013, 14, 894. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Ren, H.; Wang, G.; Chen, L.; Jiang, J.; Liu, L.; Li, N.; Zhao, J.; Sun, X.; Zhou, P. Genome-wide analysis of long non-coding RNAs at early stage of skin pigmentation in goats (Capra hircus). BMC Genom. 2016, 17, 67. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Asl, A.N.; Nazifi, S.; Ghasrodashti, A.R.; Olyaee, A. Prevalence of subclinical ketosis in dairy cattle in the Southwestern Iran and detection of cutoff point for NEFA and glucose concentrations for diagnosis of subclinical ketosis. Prev. Vet. Med. 2011, 100, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, H.; Guan, Y.; Li, X.; Lei, L.; Liu, J.; Yin, L.; Liu, G.; Wang, Z. Acetic acid activates the AMP-activated protein kinase signaling pathway to regulate lipid metabolism in bovine hepatocytes. PLoS ONE 2013, 8, e67880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Li, X.; Li, Y.; Guan, Y.; Song, Y.; Yin, L.; Chen, H.; Lei, L.; Liu, J.; Li, X.; et al. Effects of nonesterified fatty acids on the synthesis and assembly of very low density lipoprotein in bovine hepatocytes in vitro. J. Dairy Sci. 2014, 97, 1328–1335. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Hailemariam, D.; Dervishi, E.; Goldansaz, S.A.; Deng, Q.; Dunn, S.M.; Ametaj, B.N. Dairy cows affected by ketosis show alterations in innate immunity and lipid and carbohydrate metabolism during the dry off period and postpartum. Res. Vet. Sci. 2016, 107, 246–256. [Google Scholar] [CrossRef]
- Gross, J.J.; Schwarz, F.J.; Eder, K.; van Dorland, H.A.; Bruckmaier, R.M. Liver fat content and lipid metabolism in dairy cows during early lactation and during a mid-lactation feed restriction. J. Dairy Sci. 2013, 96, 5008–5017. [Google Scholar] [CrossRef] [Green Version]
- Bezerra, L.R.; de Oliveira Neto, C.B.; de Araújo, M.J.; Edvan, R.L.; de Oliveira, W.D.C.; Pereira, F.B. Major metabolic diseases affecting cows in transition period. Int. J. Biol. 2014, 6, 85–94. [Google Scholar] [CrossRef]
- González, F.D.; Muiño, R.; Pereira, V.; Campos, R.; Benedito, J.L. Relationship among blood indicators of lipomobilization and hepatic function during early lactation in high-yielding dairy cows. J. Vet. Sci. 2011, 12, 251–255. [Google Scholar]
- Berend, K.; de Vries, A.P.J.; Gans, R.O.B. Physiological approach to assessment of acid–base disturbances. N. Engl. J. Med. 2014, 371, 1434–1445. [Google Scholar] [CrossRef] [Green Version]
- Krieger, N.S.; Frick, K.K.; LaPlante Strutz, K.; Michalenka, A.; Bushinsky, D.A. Regulation of COX-2 mediates acid-induced bone calcium efflux in vitro. J. Bone Miner. Res. 2007, 22, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Iwersen, M.; Falkenberg, U.; Voigtsberger, R.; Forderung, D.; Heuwieser, W. Evaluation of an electronic cowside test to detect subclinical ketosis in dairy cows. J. Dairy Sci. 2009, 92, 2618–2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McArt, J.A.; Nydam, D.V.; Ospina, P.A.; Oetzel, G.R. A field trial on the effect of propylene glycol on milk yield and resolution of ketosis in fresh cows diagnosed with subclinical ketosis. J. Dairy Sci. 2011, 94, 6011–6020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marutsova, V.; Binev, R.; Marutsov, P. Comparative clinical and haematological investigations in lactating cows with subclinical and clinical ketosis. Mac. Vet. Rev. 2015, 38, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Puppel, K.; Kuczynska, B. Metabolic profiles of cow’s blood; a review. J. Sci. Food Agric. 2016, 96, 4321–4328. [Google Scholar] [CrossRef] [PubMed]
- Martinez, N.; Sinedino, L.D.; Bisinotto, R.S.; Ribeiro, E.S.; Gomes, G.C.; Lima, F.S.; Greco, L.F.; Risco, C.A.; Galvao, K.N.; Taylor-Rodriguez, D.; et al. Effect of induced subclinical hypocalcemia on physiological responses and neutrophil function in dairy cows. J. Dairy Sci. 2014, 97, 874–887. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, E.M.; Arís, A.; Bach, A. Associations between subclinical hypocalcemia and postparturient diseases in dairy cows. J. Dairy Sci. 2017, 100, 7427–7434. [Google Scholar] [CrossRef]
- Bosman, G.J. The involvement of erythrocyte metabolism in organismal homeostasis in health and disease. Proteom. Clin. Appl. 2016, 10, 774–777. [Google Scholar] [CrossRef]
- Trevisi, E.; Minuti, A. Assessment of the innate immune response in the periparturient cow. Res. Vet. Sci. 2018, 116, 47–54. [Google Scholar] [CrossRef]
- Sundrum, A. Metabolic Disorders in the Transition Period Indicate that the Dairy Cows’ Ability to Adapt is Overstressed. Animals 2015, 5, 978–1020. [Google Scholar] [CrossRef]
- Luo, Z.Z.; Shen, L.H.; Jiang, J.; Huang, Y.X.; Bai, L.P.; Yu, S.M.; Yao, X.P.; Ren, Z.H.; Yang, Y.X.; Cao, S.Z. Plasma metabolite changes in dairy cows during parturition identified using untargeted metabolomics. J. Dairy Sci. 2019, 102, 4639–4650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baghel, R.; Grover, S.; Kaur, H.; Jajodia, A.; Parween, S.; Sinha, J.; Srivastava, A.; Srivastava, A.K.; Bala, K.; Chandna, P.; et al. Synergistic association of STX1A and VAMP2 with cryptogenic epilepsy in North Indian population. Brain Behav. 2016, 6, e00490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, C.; Liang, T.; Kang, Y.; Lin, X.; Xie, H.; Feng, Z.P.; Gaisano, H.Y. Syntaxin-1A inhibits KATP channels by interacting with specific conserved motifs within sulfonylurea receptor 2A. J. Mol. Cell. Cardiol. 2011, 51, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Wenlan, L.; Zhongyuan, X.; Shaoqing, L.; Liying, Z.; Bo, Z.; Min, L. MiR-34a-5p mediates sevoflurane preconditioning induced inhibition of hypoxia/reoxygenation injury through STX1A in cardiomyocytes. Biomed. Pharmacother. 2018, 102, 153–159. [Google Scholar] [CrossRef]
- Cao, F.; Hata, R.; Zhu, P.; Niinobe, M.; Sakanaka, M. Up-regulation of syntaxin1 in ischemic cortex after permanent focal ischemia in rats. Brain Res. 2009, 1272, 52–61. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.-L.; Chen, S.-Y.; Qin, C.; Jia, X.; Deng, F.; Wang, J.; Lai, S.-J. Clinical Ketosis-Associated Alteration of Gene Expression in Holstein Cows. Genes 2020, 11, 219. https://doi.org/10.3390/genes11020219
Wu Z-L, Chen S-Y, Qin C, Jia X, Deng F, Wang J, Lai S-J. Clinical Ketosis-Associated Alteration of Gene Expression in Holstein Cows. Genes. 2020; 11(2):219. https://doi.org/10.3390/genes11020219
Chicago/Turabian StyleWu, Zhou-Lin, Shi-Yi Chen, Chao Qin, Xianbo Jia, Feilong Deng, Jie Wang, and Song-Jia Lai. 2020. "Clinical Ketosis-Associated Alteration of Gene Expression in Holstein Cows" Genes 11, no. 2: 219. https://doi.org/10.3390/genes11020219
APA StyleWu, Z.-L., Chen, S.-Y., Qin, C., Jia, X., Deng, F., Wang, J., & Lai, S.-J. (2020). Clinical Ketosis-Associated Alteration of Gene Expression in Holstein Cows. Genes, 11(2), 219. https://doi.org/10.3390/genes11020219