Prolactin-Responsive Circular RNA circHIPK3 Promotes Proliferation of Mammary Epithelial Cells from Dairy Cow

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Cell Lines
2.2. Cell Culture
2.3. RNA Library Preparation
2.4. Real-Time PCR
2.5. Western Blotting Analysis
2.6. Cell Proliferation Assay
2.7. Statistical Analysis
3. Results
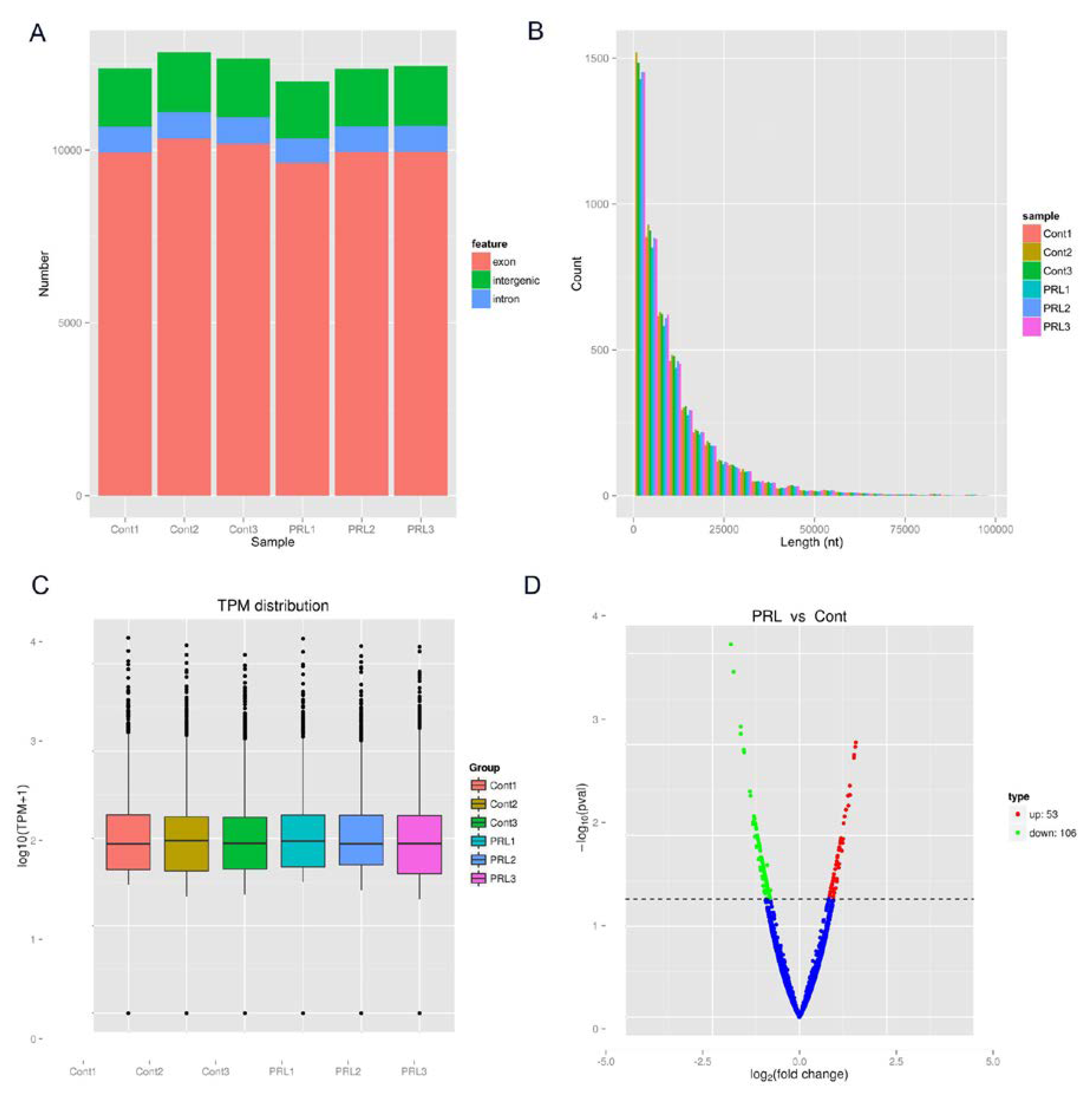
3.1. Length Source Distribution and the Differential Expression of circRNAs
3.2. Identification of circHIPK3 as a Candidate circRNA
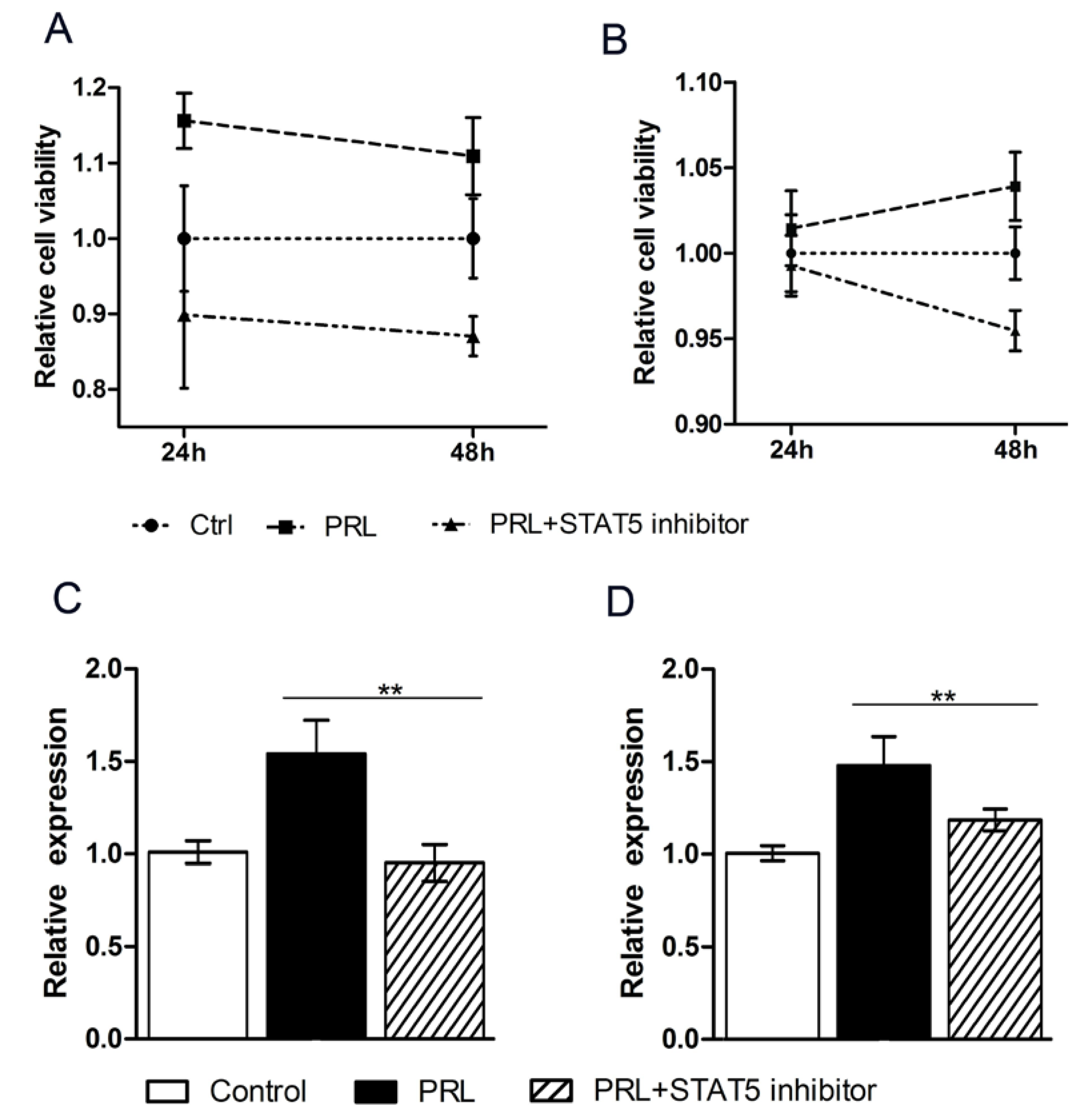
3.3. CircHIPK3 Promotes the Proliferation of HC11 Cells
3.4. The STAT Pathway Promotes the Proliferation of Mammary Epithelial Cells
3.5. PRL Affects circHIPK3 Expression and Regulates Alternative Splicing Factors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Capuco, A.V.; Ellis, S.E.; Hale, S.A.; Long, E.; Erdman, R.A.; Zhao, X.; Paape, M.J. Lactation persistency: Insights from mammary cell proliferation studies. J. Anim. Sci. 2003, 81, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Oakes, S.R.; Rogers, R.L.; Naylor, M.J.; Ormandy, C.J. Prolactin regulation of mammary gland development. J. Mammary Gland Biol. 2008, 13, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Ormandy, C.J.; Camus, A.; Barra, J.; Damotte, D.; Lucas, B.; Buteau, H.; Edery, M.; Brousse, N.; Babinet, N.; Kelly, P.A. Null mutation of the prolactin receptor gene produces multiple reproductive defects in the mouse. Genes 1997, 11, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Alex, C.; Pasquale, P.; Adrián, S.M.; Eva, M.T.; Jordi, S.M.; Chetan, D.; Willian, C.D.; Miguel, A.P.; Eva, G.S. Rankl impairs lactogenic differentiation through inhibition of the prolactin/stat5 pathway at midgestation. Stem Cells 2016, 34, 1027–1039. [Google Scholar]
- Boutinaud, M.; Lollivier, V.; Finot, L.; Bruckmaier, R.M.; Lacasse, P. Mammary cell activity and turnover in dairy cows treated with the prolactin-release inhibitor quinagolide and milked once daily. J. Dairy Sci. 2012, 95, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Lollivier, V.; Lacasse, P.; Angulo-Arizala, J.; Lamberton, P.; Wiart, S.; Portanguen, J.; Bruckmaier, R.; Boutinaud, M. In vivo inhibition followed by exogenous supplementation demonstrates galactopoietic effects of prolactin on mammary tissue and milk production in dairy cows. J. Dairy Sci. 2015, 98, 8775–8787. [Google Scholar] [CrossRef]
- Olazabal, I.; Munoz, J.; Ogueta, S.; Obregon, E.; Garcia-Ruiz, J.P. Prolactin (PRL)-PRL receptor system increases cell proliferation involving JNK (c-Jun amino terminal kinase) and AP-1 activation: Inhibition by glucocorticoids. Mol. Endocrinol 2000, 14, 564–575. [Google Scholar] [CrossRef]
- Wagner, K.U.; Krempler, A.; Triplett, A.A.; Qi, Y.; George, N.M.; Zhu, J.Q.; Rui, H. Impaired alveologenesis and maintenance of secretory mammary epithelial cells in jak2 conditional knockout mice. Mol. Cell. Biol. 2004, 24, 5510–5520. [Google Scholar] [CrossRef]
- Shillingford, J.M.; Miyoshi, K.; Robinson, G.W.; Grimm, S.L.; Rosen, J.M.; Neubauer, H.; Pfeffer, K.; Hennighausen, L. Jak2 is an essential tyrosine kinase involved in pregnancy-mediated development of mammary secretory epithelium. Mol. Endocrinol. 2002, 16, 563–570. [Google Scholar] [CrossRef]
- Cui, Y.; Riedlinger, G.; Miyoshi, K.; Tang, W.; Li, C.; Deng, C.X.; Robinson, G.W.; Hennighausen, L. Inactivation of stat5 in mouse mammary epithelium during pregnancy reveals distinct functions in cell proliferation, survival, and differentiation. Mol. Cell. Biol. 2004, 24, 8037–8047. [Google Scholar] [CrossRef]
- Zhou, J.; Chehab, R.; Tkalcevic, J.; Naylor, M.J.; Harris, J.; Wilson, T.J.; Tsao, S.; Tellis, I.; Zavarsek, S.; Xu, D.; et al. Elf5 is essential for early embryogenesis and mammary gland development during pregnancy and lactation. EMBO J. 2005, 24, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Nigro, J.M.; Cho, K.R.; Fearon, E.R.; Kern, S.E.; Vogelstein, B. Scrambled exons. Cell 1991, 64, 607–613. [Google Scholar] [CrossRef]
- Zhang, C.L.; Wu, H.; Wang, Y.H.; Zhao, Y.; Fang, X.T.; Chen, C.; Chen, H. Expression patterns of circular RNAs from primary kinase transcripts in the mammary glands of lactating rats. J. Breast Cancer 2015, 18, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.L.; Wu, H.; Wang, Y.H.; Zhu, S.Q.; Liu, J.Q.; Fang, X.T.; Chen, H. Circular RNA of cattle casein genes are highly expressed in bovine mammary gland. J. Dairy Sci. 2016, 99, 4750–4760. [Google Scholar] [CrossRef]
- Zheng, Q.; Bao, C.; Guo, W.; Li, S.; Chen, J.; Chen, B.; Luo, Y.; Lyu, D.; Li, Y.; Shi, G.; et al. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat Commun. 2016, 7, 11215. [Google Scholar] [CrossRef]
- Zeng, K.; Chen, X.; Xu, M.; Liu, X.; Hu, X.; Xu, T.; Sun, H.; Pan, Y.; He, B.; Wang, S. CircHIPK3 promotes colorectal cancer growth and metastasis by sponging miR-7. Cell Death Dis. 2018, 9, 417. [Google Scholar] [CrossRef]
- Cai, C.; Zhi, Y.; Wang, K.; Zhang, P.; Ji, Z.; Xie, C.; Sun, F. CircHIPK3 overexpression accelerates the proliferation and invasion of prostate cancer cells through regulating miRNA-338-3p. OncoTargets Ther. 2019, 12, 3363–3372. [Google Scholar] [CrossRef]
- Chen, B.; Yu, J.; Guo, L.; Byers, M.; Wang, Z.; Chen, X.; Xu, H.; Nie, Q. Circular RNA circHIPK3 promotes the proliferation and differentiation of chicken myoblast cells by sponging miR-30a-3p. Cells 2019, 8, 177. [Google Scholar] [CrossRef]
- Li, D.; Xie, X.; Wang, J.; Bian, Y.; Li, Q.; Gao, X.; Wang, C. MiR-486 regulates lactation and targets the PTEN gene in cow mammary glands. PLoS ONE 2015, 10, e0118284. [Google Scholar] [CrossRef]
- Gong, D.; Ferrell, J.J. The roles of cyclin A2, B1, and B2 in early and late mitotic events. Mol. Biol. Cell 2010, 21, 3149–3161. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Patel, N.A.; Watson, J.E.; Apostolatos, H.; Kleiman, E.; Hanson, O.; Hagiwara, M.; Cooper, D.R. Akt2 regulation of Cdc2-Like kinases (Clk/Sty), Serine/Arginine-Rich (SR) protein phosphorylation, and Insulin-Induced alternative splicing of PKCβII messenger ribonucleic acid. Endocrinology 2009, 150, 2087–2097. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Ebbesen, K.K.; Kjems, J.; Hansen, T.B. Circular RNAs: Identification, biogenesis and function. Biochim. Biophys. Acta Gene Regul. Mech. 2016, 1859, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Khaitovich, P.; Kelso, J.; Franz, H.; Visagie, J.; Giger, T.; Joerchel, S.; Petzold, E.; Green, R.E.; Lachmann, M.; Paabo, S. Functionality of intergenic transcription: An evolutionary comparison. PLoS Genet. 2006, 2, 1590–1598. [Google Scholar] [CrossRef]
- Bolha, L.; Ravnik, M.; Glavač, D. Circular RNAs: Biogenesis, function, and a role as possible cancer biomarkers. Int. J. Genomics 2017, 2017, 6218353. [Google Scholar] [CrossRef]
- Kai, D.; Yannian, L.; Yitian, C.; Dinghao, G.; Xin, Z.; Wu, J. Circular RNA HIPK3 promotes gallbladder cancer cell growth by sponging microRNA-124. Biochem. Bioph. Res. Commun. 2018, 503, 863–869. [Google Scholar] [CrossRef]
- Yu, H.; Chen, Y.; Jiang, P. Circular RNA HIPK3 exerts oncogenic properties through suppression of miR-124 in lung cancer. Biochem. Bioph. Res. Commun. 2018, 506, 455–462. [Google Scholar] [CrossRef]
- Zhu, C.; Yao, C.; Zhu, L.; She, C.; Zhou, X. Dexamethasone-induced cytotoxicity in human osteoblasts is associated with circular RNA HIPK3 downregulation. Biochem. Bioph. Res. Commun. 2019, 516, 645–652. [Google Scholar] [CrossRef]
- Chen, G.; Shi, Y.; Liu, M.; Sun, J. CircHIPK3 regulates cell proliferation and migration by sponging miR-124 and regulating AQP3 expression in hepatocellular carcinoma. Cell Death Dis. 2018, 9, 175–188. [Google Scholar] [CrossRef]
- Do, D.N.; Li, R.; Dudemaine, P.; Ibeagha-Awemu, E.M. MicroRNA roles in signalling during lactation: An insight from differential expression, time course and pathway analyses of deep sequence data. Sci. Rep. 2017, 7, 44605. [Google Scholar] [CrossRef]
- Lin, X.; Luo, J.; Zhang, L.; Wang, W.; Gou, D. MiR-103 controls milk fat accumulation in goat (Capra hircus) mammary gland during lactation. PLoS ONE 2013, 8, e79258. [Google Scholar] [CrossRef]
- Zhang, C.L.; Zhao, Y.; Wang, Y.H.; Wu, H.; Fang, X.T.; Chen, H. Deep RNA sequencing reveals that MicroRNAs play a key role in lactation in rats. J. Nutr. 2014, 144, 1142–1149. [Google Scholar] [CrossRef] [PubMed]
- García-Vazquez, R.; Ruiz-García, E.; Meneses-García, A.; Astudillo-de la Vega, H.; Lara-Medina, F.; Alvarado-Miranda, A.; Maldonado-Martínez, H.; González-Barrios, J.A.; Campos-Parra, A.D.; Cuevas, S.R.; et al. A microRNA signature associated with pathological complete response to novel neoadjuvant therapy regimen in triple-negative breast cancer. Tumor Biol. 2017, 39, 568835407. [Google Scholar] [CrossRef]
- Hennighausen, L.; Robinson, G.W.; Wagner, K.U.; Liu, W. Prolactin signaling in mammary gland development. J. Biol. Chem. 1997, 272, 7567–7569. [Google Scholar] [CrossRef] [PubMed]
- Mapes, J.; Li, Q.; Kannan, A.; Anandan, L.; Laws, M.; Lydon, J.P.; Bagchi, I.C.; Bagchi, M.K. CUZD1 is a critical mediator of the JAK/STAT5 signaling pathway that controls mammary gland development during pregnancy. PLoS Genet. 2017, 13, e1006654. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, T.W.; Graveley, B.R. Expansion of the eukaryotic proteome by alternative splicing. Nature 2010, 463, 457–463. [Google Scholar] [CrossRef]
- Weg-Remers, S.; Ponta, H.; Herrlich, P.; König, H. Regulation of alternative pre-mRNA splicing by the ERK MAP-kinase pathway. EMBO J. 2001, 20, 4194–4203. [Google Scholar] [CrossRef]
- Li, X.; Liu, C.; Xue, W.; Zhang, Y.; Jiang, S.; Yin, Q.; Wei, J.; Yao, R.; Yang, L.; Chen, L. Coordinated circRNA biogenesis and function with NF90/NF110 in viral infection. Mol. Cell 2017, 67, 214–227. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Zhang, M.; Li, D.; Li, M.; Kong, L.; Cao, M.; Wang, Y.; Song, C.; Fang, X.; Chen, H.; et al. Prolactin-Responsive Circular RNA circHIPK3 Promotes Proliferation of Mammary Epithelial Cells from Dairy Cow. Genes 2020, 11, 336. https://doi.org/10.3390/genes11030336
Liu J, Zhang M, Li D, Li M, Kong L, Cao M, Wang Y, Song C, Fang X, Chen H, et al. Prolactin-Responsive Circular RNA circHIPK3 Promotes Proliferation of Mammary Epithelial Cells from Dairy Cow. Genes. 2020; 11(3):336. https://doi.org/10.3390/genes11030336
Chicago/Turabian StyleLiu, Jia, MoLan Zhang, DeWei Li, MengLu Li, LingHao Kong, MengWen Cao, YanHong Wang, ChengChuang Song, XingTang Fang, Hong Chen, and et al. 2020. "Prolactin-Responsive Circular RNA circHIPK3 Promotes Proliferation of Mammary Epithelial Cells from Dairy Cow" Genes 11, no. 3: 336. https://doi.org/10.3390/genes11030336
APA StyleLiu, J., Zhang, M., Li, D., Li, M., Kong, L., Cao, M., Wang, Y., Song, C., Fang, X., Chen, H., Xu, H., & Zhang, C. (2020). Prolactin-Responsive Circular RNA circHIPK3 Promotes Proliferation of Mammary Epithelial Cells from Dairy Cow. Genes, 11(3), 336. https://doi.org/10.3390/genes11030336