Cannabinoid Control of Olfactory Processes: The Where Matters


{kind=link}
Abstract
:1. The Endocannabinoid System: A General Overview
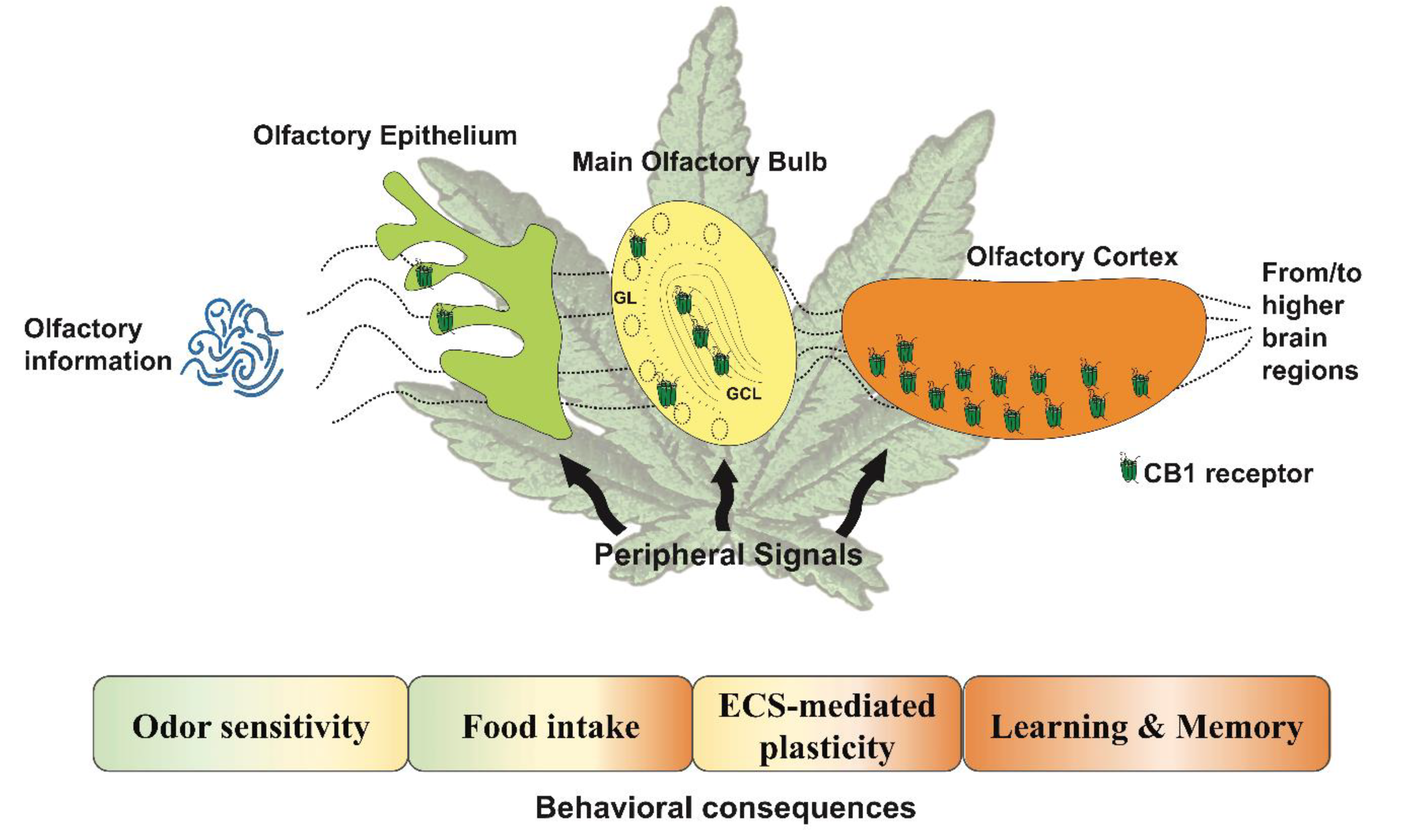
2. Role of the Endocannabinoid System in Olfactory Circuits
3. The Endocannabinoid System in the Olfactory Epithelium
4. The Endocannabinoid System in the Olfactory Bulb
5. The Endocannabinoid System in the Piriform Cortex
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- United Nations Office on Drugs, and Crime. World Drug Report; United Nations Publications: New York, NY, USA, 2010. [Google Scholar]
- Araque, A.; Castillo, P.E.; Manzoni, O.J.; Tonini, R. Synaptic functions of endocannabinoid signaling in health and disease. Neuropharmacology 2017, 124, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Kano, M.; Ohno-Shosaku, T.; Hashimotodani, Y.; Uchigashima, M.; Watanabe, M. Endocannabinoid-mediated control of synaptic transmission. Physiol. Rev. 2009, 89, 309–380. [Google Scholar] [CrossRef] [PubMed]
- Maejima, T.; Hashimoto, K.; Yoshida, T.; Aiba, A.; Kano, M. Presynaptic inhibition caused by retrograde signal from metabotropic glutamate to cannabinoid receptors. Neuron 2001, 31, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Rodríguez, A.; Puente, N.; Elezgarai, I.; Ruehle, S.; Lutz, B.; Reguero, L.; Gerrikagoitia, I.; Marsicano, G.; Grandes, P. Anatomical characterization of the cannabinoid CB1 receptor in cell-type-specific mutant mouse rescue models. J. Comp. Neurol. 2017, 525, 302–318. [Google Scholar] [CrossRef]
- Gutiérrez-Rodríguez, A.; Bonilla-Del Río, I.; Puente, N.; Gómez-Urquijo, S.M.; Fontaine, C.J.; Egaña-Huguet, J.; Elezgarai, I.; Ruehle, S.; Lutz, B.; Robin, L.M.; et al. Localization of the cannabinoid type-1 receptor in subcellular astrocyte compartments of mutant mouse hippocampus. Glia 2018, 66, 1417–1431. [Google Scholar] [CrossRef]
- Kreitzer, A.C.; Regehr, W.G. Retrograde inhibition of presynaptic calcium influx by endogenous cannabinoids at excitatory synapses onto purkinje cells. Neuron 2001, 29, 717–727. [Google Scholar] [CrossRef] [Green Version]
- Ohno-Shosaku, T.; Maejima, T.; Kano, M. Endogenous cannabinoids mediate retrograde signals from depolarized postsynaptic neurons to presynaptic terminals. Neuron 2001, 29, 729–738. [Google Scholar] [CrossRef]
- Wilson, R.I.; Nicoll, R.A. Endogenous cannabinoids mediate retrograde signalling at hippocampal synapses. Nature 2001, 410, 588–592. [Google Scholar] [CrossRef]
- Chevaleyre, V.; Castillo, P.E. Heterosynaptic LTD of hippocampal GABAergic synapses: A novel role of endocannabinoids in regulating excitability. Neuron 2003, 38, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Marsicano, G.; Wotjak, C.T.; Azad, S.C.; Bisogno, T.; Rammes, G.; Cascio, M.G.; Hermann, H.; Tang, J.; Hofmann, C.; Zieglgänsberger, W.; et al. The endogenous cannabinoid system controls extinction of aversive memories. Nature 2002, 418, 530–534. [Google Scholar] [CrossRef]
- Bacci, A.; Huguenard, J.R.; Prince, D.A. Long-lasting self-inhibition of neocortical interneurons mediated by endocannabinoids. Nature 2004, 431, 312–316. [Google Scholar] [CrossRef]
- Gerdeman, G.L.; Ronesi, J.; Lovinger, D.M. Postsynaptic endocannabinoid release is critical to long-term depression in the striatum. Nat. Neurosci. 2002, 5, 446–451. [Google Scholar] [CrossRef]
- Peñasco, S.; Rico-Barrio, I.; Puente, N.; Gómez-Urquijo, S.M.; Fontaine, C.J.; Egaña-Huguet, J.; Achicallende, S.; Ramos, A.; Reguero, L.; Elezgarai, I.; et al. Endocannabinoid long-term depression revealed at medial perforant path excitatory synapses in the dentate gyrus. Neuropharmacology 2019, 153, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, A.; Felder, C.C.; Herkenham, M.; Mackie, K.; Martin, B.R.; et al. International union of pharmacology. XXVII. Classification of cannabinoid receptors. Pharm. Rev. 2002, 54, 161–202. [Google Scholar] [CrossRef] [PubMed]
- Marsicano, G.; Lafenêtre, P. Roles of the endocannabinoid system in learning and memory. Curr. Top. Behav. Neurosci. 2009, 1, 201–230. [Google Scholar] [PubMed]
- Morena, M.; Campolongo, P. The endocannabinoid system: An emotional buffer in the modulation of memory function. Neurobiol. Learn. Mem. 2014, 112, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Soria-Gómez, E.; Bellocchio, L.; Marsicano, G. New insights on food intake control by olfactory processes: The emerging role of the endocannabinoid system. Mol. Cell. Endocrinol. 2014, 397, 59–66. [Google Scholar] [CrossRef]
- Tart, C.T. Marijuana intoxication: Common experiences. Nature 1970, 226, 701–704. [Google Scholar] [CrossRef]
- Herkenham, M.; Lynn, A.B.; Little, M.D.; Johnson, M.R.; Melvin, L.S.; de Costa, B.R.; Rice, K.C. Cannabinoid receptor localization in brain. Proc. Natl. Acad. Sci. USA 1990, 87, 1932–1936. [Google Scholar] [CrossRef] [Green Version]
- Herkenham, M.; Lynn, A.; Johnson, M.; Melvin, L.; de Costa, B.; Rice, K. Characterization and localization of cannabinoid receptors in rat brain: A quantitative in vitro autoradiographic study. J. Neurosci. 1991, 11, 563–583. [Google Scholar] [CrossRef] [PubMed]
- Marsicano, G.; Lutz, B. Expression of the cannabinoid receptor CB1 in distinct neuronal subpopulations in the adult mouse forebrain. Eur. J. Neurosci. 1999, 11, 4213–4225. [Google Scholar] [CrossRef] [PubMed]
- Pettit, D.A.D.; Harrison, M.P.; Olson, J.M.; Spencer, R.F.; Cabral, G.A. Immunohistochemical localization of the neural cannabinoid receptor in rat brain. J. Neurosci. Res. 1998, 51, 391–402. [Google Scholar] [CrossRef]
- Czesnik, D.; Schild, D.; Kuduz, J.; Manzini, I. Cannabinoid action in the olfactory epithelium. Proc. Natl. Acad. Sci. USA 2007, 104, 2967–2972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breunig, E.; Manzini, I.; Piscitelli, F.; Gutermann, B.; Di Marzo, V.; Schild, D.; Czesnik, D. The endocannabinoid 2-arachidonoyl-glycerol controls odor sensitivity in larvae of xenopus laevis. J. Neurosci. 2010, 30, 8965–8973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breunig, E.; Czesnik, D.; Piscitelli, F.; Di Marzo, V.; Manzini, I.; Schild, D. Endocannabinoid modulation in the olfactory epithelium. In Sensory and Metabolic Control of Energy Balance; Meyerhof, W., Beisiegel, U., Joost, H.-G., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 52, pp. 139–145. [Google Scholar]
- Hutch, C.R.; Hillard, C.J.; Jia, C.; Hegg, C.C. An endocannabinoid system is present in the mouse olfactory epithelium but does not modulate olfaction. Neuroscience 2015, 300, 539–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, A.; Jaffé, E.H. Acute immobilization stress modulate GABA release from rat olfactory bulb: Involvement of endocannabinoids—Cannabinoids and acute stress modulate GABA release. Int. J. Cell Biol. 2011, 2011, 1–10. [Google Scholar] [CrossRef]
- Harvey, J.; Heinbockel, T. Neuromodulation of synaptic transmission in the main olfactory bulb. Int. J. Environ. Res. Public Health 2018, 15, 2194. [Google Scholar] [CrossRef] [Green Version]
- Heinbockel, T.; Wang, Z.-J.; Brown, E.A.; Austin, P.T. Endocannabinoid signaling in neural circuits of the olfactory and limbic system. In Cannabinoids in Health and Disease; Meccariello, R., Chianese, R., Eds.; InTech: London, UK, 2016. [Google Scholar]
- Pouille, F.; Schoppa, N.E. Cannabinoid receptors modulate excitation of an olfactory bulb local circuit by cortical feedback. Front. Cell. Neurosci. 2018, 12, 47. [Google Scholar] [CrossRef] [Green Version]
- Soria-Gómez, E.; Bellocchio, L.; Reguero, L.; Lepousez, G.; Martin, C.; Bendahmane, M.; Ruehle, S.; Remmers, F.; Desprez, T.; Matias, I.; et al. The endocannabinoid system controls food intake via olfactory processes. Nat. Neurosci. 2014, 17, 407–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.-J.; Sun, L.; Heinbockel, T. Cannabinoid receptor-mediated regulation of neuronal activity and signaling in glomeruli of the main olfactory bulb. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 8475–8479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.-J.; Hu, S.S.-J.; Bradshaw, H.B.; Sun, L.; Mackie, K.; Straiker, A.; Heinbockel, T. Cannabinoid receptor-mediated modulation of inhibitory inputs to mitral cells in the main olfactory bulb. J. Neurophysiol. 2019, 122, 749–759. [Google Scholar] [CrossRef]
- Ghosh, S.; Reuveni, I.; Zidan, S.; Lamprecht, R.; Barkai, E. Learning-induced modulation of the effect of endocannabinoids on inhibitory synaptic transmission. J. Neurophysiol. 2018, 119, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.J.; Weston, S.E.; Jones, N.A.; Smith, I.; Bevan, S.A.; Williamson, E.M.; Stephens, G.J.; Williams, C.M.; Whalley, B.J. Δ9-Tetrahydrocannabivarin suppresses in vitro epileptiform and in vivo seizure activity in adult rats: Anticonvulsant potential of Δ9-THCV. Epilepsia 2010, 51, 1522–1532. [Google Scholar] [CrossRef] [PubMed]
- Terral, G.; Busquets-Garcia, A.; Varilh, M.; Achicallende, S.; Cannich, A.; Bellocchio, L.; Bonilla-Del Río, I.; Massa, F.; Puente, N.; Soria-Gomez, E.; et al. CB1 Receptors in the anterior piriform cortex control odor preference memory. Curr. Biol. 2019, 29, 2455–2464. [Google Scholar] [CrossRef] [PubMed]
- Zenko, M.; Zhu, Y.; Dremencov, E.; Ren, W.; Xu, L.; Zhang, X. Requirement for the endocannabinoid system in social interaction impairment induced by coactivation of dopamine D1 and D2 receptors in the piriform cortex. J. Neurosci. Res. 2011, 89, 1245–1258. [Google Scholar] [CrossRef] [PubMed]
- Bhutani, S.; Howard, J.D.; Reynolds, R.; Zee, P.C.; Gottfried, J.; Kahnt, T. Olfactory connectivity mediates sleep-dependent food choices in humans. eLife 2019, 8, e49053. [Google Scholar] [CrossRef]
- Terral, G.; Varilh, M.; Cannich, A.; Massa, F.; Ferreira, G.; Marsicano, G. Synaptic Functions of Type-1 Cannabinoid receptors in inhibitory circuits of the anterior piriform cortex. Neuroscience 2020, 433, 121–131. [Google Scholar] [CrossRef]
- Busquets-Garcia, A.; Oliveira da Cruz, J.F.; Terral, G.; Zottola, A.C.P.; Soria-Gómez, E.; Contini, A.; Martin, H.; Redon, B.; Varilh, M.; Ioannidou, C.; et al. Hippocampal CB1 receptors control incidental associations. Neuron 2018, 99, 1247–1259. [Google Scholar] [CrossRef] [Green Version]
- Soria-Gómez, E.; Busquets-Garcia, A.; Hu, F.; Mehidi, A.; Cannich, A.; Roux, L.; Louit, I.; Alonso, L.; Wiesner, T.; Georges, F.; et al. Habenular CB1 receptors control the expression of aversive memories. Neuron 2015, 88, 306–313. [Google Scholar] [CrossRef] [Green Version]
- Laviolette, S.R.; Grace, A.A. Cannabinoids potentiate emotional learning plasticity in neurons of the medial prefrontal cortex through basolateral amygdala inputs. J. Neurosci. 2006, 26, 6458–6468. [Google Scholar] [CrossRef] [Green Version]
- Back, F.P.; Carobrez, A.P. Periaqueductal gray glutamatergic, cannabinoid and vanilloid receptor interplay in defensive behavior and aversive memory formation. Neuropharmacology 2018, 135, 399–411. [Google Scholar] [CrossRef]
- O’Doherty, J.; Rolls, E.T.; Francis, S.; Bowtell, R.; McGlone, F.; Kobal, G.; Renner, B.; Ahne, G. Sensory-specific satiety-related olfactory activation of the human orbitofrontal cortex. Neuroreport 2000, 4, 893–897. [Google Scholar] [CrossRef] [PubMed]
- Pager, J.; Giachetti, I.; Holley, A.; Le Magnen, J. A selective control of olfactory bulb electrical activity in relation to food deprivation and satiety in rats. Physiol. Behav. 1972, 9, 573–579. [Google Scholar] [CrossRef]
- Di Marzo, V.; Matias, I. Endocannabinoid control of food intake and energy balance. Nat. Neurosci. 2005, 8, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Palouzier-Paulignan, B.; Lacroix, M.-C.; Aime, P.; Baly, C.; Caillol, M.; Congar, P.; Julliard, A.K.; Tucker, K.; Fadool, D.A. Olfaction under metabolic influences. Chem. Senses 2012, 37, 769–797. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, A. Genetic Manipulation of the Endocannabinoid System; Springer: New York, NY, USA, 2015. [Google Scholar]
- Pinching, A.J.; Powell, T.P.S. The neuropil of the glomeruli of the olfactory bulb. J. Cell Sci. 1971, 9, 347–377. [Google Scholar]
- Piredda, S.; Gale, K. A crucial epileptogenic site in the deep prepiriform cortex. Nature 1985, 317, 623–625. [Google Scholar] [CrossRef]
- Marsicano, G.; Goodenough, S.; Monory, K.; Hermann, H.; Eder, M.; Cannich, A.; Azad, S.C.; Cascio, M.G.; Gutiérrez, S.O.; van der Stelt, M.; et al. CB1 cannabinoid receptors and on-demand defense against excitotoxicity. Science 2003, 302, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Monory, K.; Massa, F.; Egertová, M.; Eder, M.; Blaudzun, H.; Westenbroek, R.; Kelsch, W.; Jacob, W.; Marsch, R.; Ekker, M.; et al. The endocannabinoid system controls key epileptogenic circuits in the hippocampus. Neuron 2006, 51, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Neville, K.R.; Haberly, L.B. Olfactory cortex. In The Synaptic Organization of the Brain, 5th ed.; Shepherd, G.M., Ed.; Oxford University Press: Oxford, UK, 2004; Volume 8, pp. 415–454. [Google Scholar]
- Sigel, E.; Baur, R.; Racz, I.; Marazzi, J.; Smart, T.G.; Zimmer, A.; Gertsch, J. The major central endocannabinoid directly acts at GABAA receptors. Proc. Natl. Acad. Sci. USA 2011, 108, 18150–18155. [Google Scholar] [CrossRef] [Green Version]
- Gottfried, J.A.; O’Doherty, J.; Dolan, R.J. Appetitive and aversive olfactory learning in humans studied using event-related functional magnetic resonance imaging. J. Neurosci. 2002, 22, 10829–10837. [Google Scholar] [CrossRef] [Green Version]
- Buchanan, T.W. A specific role for the human amygdala in olfactory memory. Learn. Mem. 2003, 10, 319–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoenbaum, G.; Eichenbaum, H. Information coding in the rodent prefrontal cortex. I. Single-neuron activity in orbitofrontal cortex compared with that in pyriform cortex. J. Neurophysiol. 1995, 74, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Nogi, Y.; Ahasan, M.M.; Murata, Y.; Taniguchi, M.; Sha, M.F.R.; Ijichi, C.; Yamaguchi, M. Expression of feeding-related neuromodulatory signalling molecules in the mouse central olfactory system. Sci. Rep. 2020, 10, 890. [Google Scholar] [CrossRef]
- Lotsch, J.; Hummel, T. Cannabinoid-related olfactory neuroscience in mice and humans. Chem. Senses 2015, 40, 3–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, C.; Oertel, B.G.; Ludyga, D.; Ultsch, A.; Hummel, T.; Lötsch, J. Effects of 20 mg oral Δ9-tetrahydrocannabinol on the olfactory function of healthy volunteers: Effects of Δ9-tetrahydrocannabinol on olfaction. Br. J. Clin. Pharm. 2014, 78, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Walter, C.; Oertel, B.G.; Felden, L.; Nöth, U.; Vermehren, J.; Deichmann, R.; Lötsch, J. Effects of oral Δ9-tetrahydrocannabinol on the cerebral processing of olfactory input in healthy non-addicted subjects. Eur. J. Clin. Pharm. 2017, 73, 1579–1587. [Google Scholar] [CrossRef]
- Galindo, L.; Moreno, E.; López-Armenta, F.; Guinart, D.; Cuenca-Royo, A.; Izquierdo-Serra, M.; Xicota, L.; Fernandez, C.; Menoyo, E.; Fernández-Fernández, J.M.; et al. Cannabis users show enhanced expression of CB1-5HT2A receptor heteromers in olfactory neuroepithelium cells. Mol. Neurobiol. 2018, 55, 6347–6361. [Google Scholar] [CrossRef] [Green Version]
- Bellocchio, L.; Lafenêtre, P.; Cannich, A.; Cota, D.; Puente, N.; Grandes, P.; Chaouloff, F.; Piazza, P.V.; Marsicano, G. Bimodal control of stimulated food intake by the endocannabinoid system. Nat. Neurosci. 2010, 13, 281–283. [Google Scholar] [CrossRef]
- Basavarajappa, B.S.; Shivakumar, M.; Joshi, V.; Subbanna, S. Endocannabinoid system in neurodegenerative disorders. J. Neurochem. 2017, 142, 624–648. [Google Scholar] [CrossRef]
- Godoy, M.; Voegels, R.; Pinna, F.; Imamura, R.; Farfel, J. Olfaction in neurologic and neurodegenerative diseases: A literature review. Int. Arch. Otorhinolaryngol. 2014, 19, 176–179. [Google Scholar] [PubMed] [Green Version]
- Philpott, C.M.; Boak, D. The impact of olfactory disorders in the United Kingdom. Chem. Senses 2014, 39, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Yin, A.; Wang, F.; Zhang, X. Integrating endocannabinoid signaling in the regulation of anxiety and depression. Acta Pharm. Sin. 2018, 40, 336–341. [Google Scholar] [CrossRef] [PubMed]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terral, G.; Marsicano, G.; Grandes, P.; Soria-Gómez, E. Cannabinoid Control of Olfactory Processes: The Where Matters. Genes 2020, 11, 431. https://doi.org/10.3390/genes11040431
Terral G, Marsicano G, Grandes P, Soria-Gómez E. Cannabinoid Control of Olfactory Processes: The Where Matters. Genes. 2020; 11(4):431. https://doi.org/10.3390/genes11040431
Chicago/Turabian StyleTerral, Geoffrey, Giovanni Marsicano, Pedro Grandes, and Edgar Soria-Gómez. 2020. "Cannabinoid Control of Olfactory Processes: The Where Matters" Genes 11, no. 4: 431. https://doi.org/10.3390/genes11040431
APA StyleTerral, G., Marsicano, G., Grandes, P., & Soria-Gómez, E. (2020). Cannabinoid Control of Olfactory Processes: The Where Matters. Genes, 11(4), 431. https://doi.org/10.3390/genes11040431