A Comprehensive Cytogenetic Analysis of Several Members of the Family Columbidae (Aves, Columbiformes)

 , , ,
, , ,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Cell Culture and Chromosome Preparation
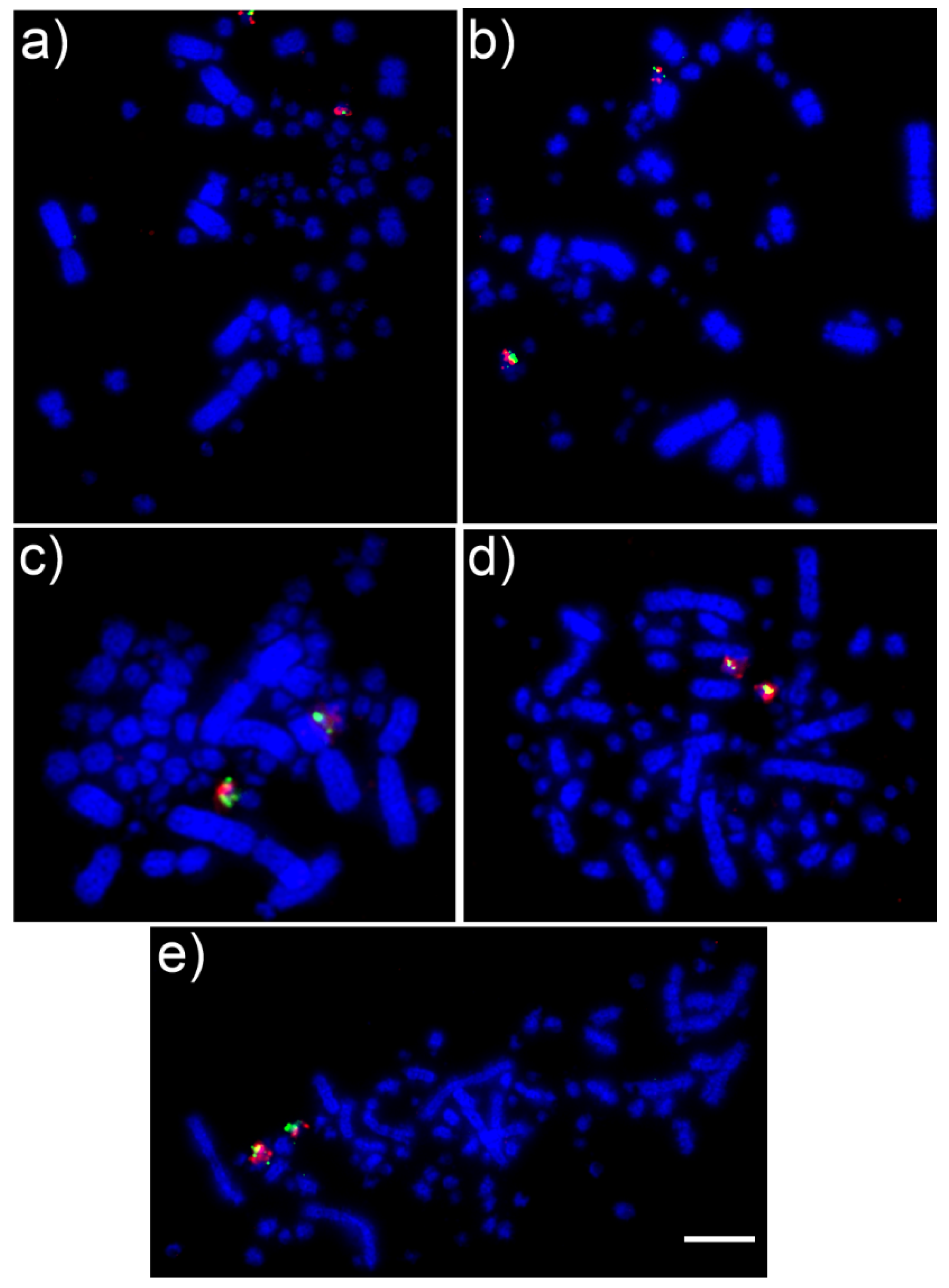
2.2. Comparative Chromosome Painting
2.3. FISH with BAC Probes
2.4. G-Banding
2.5. Phylogenetic Analysis
3. Results
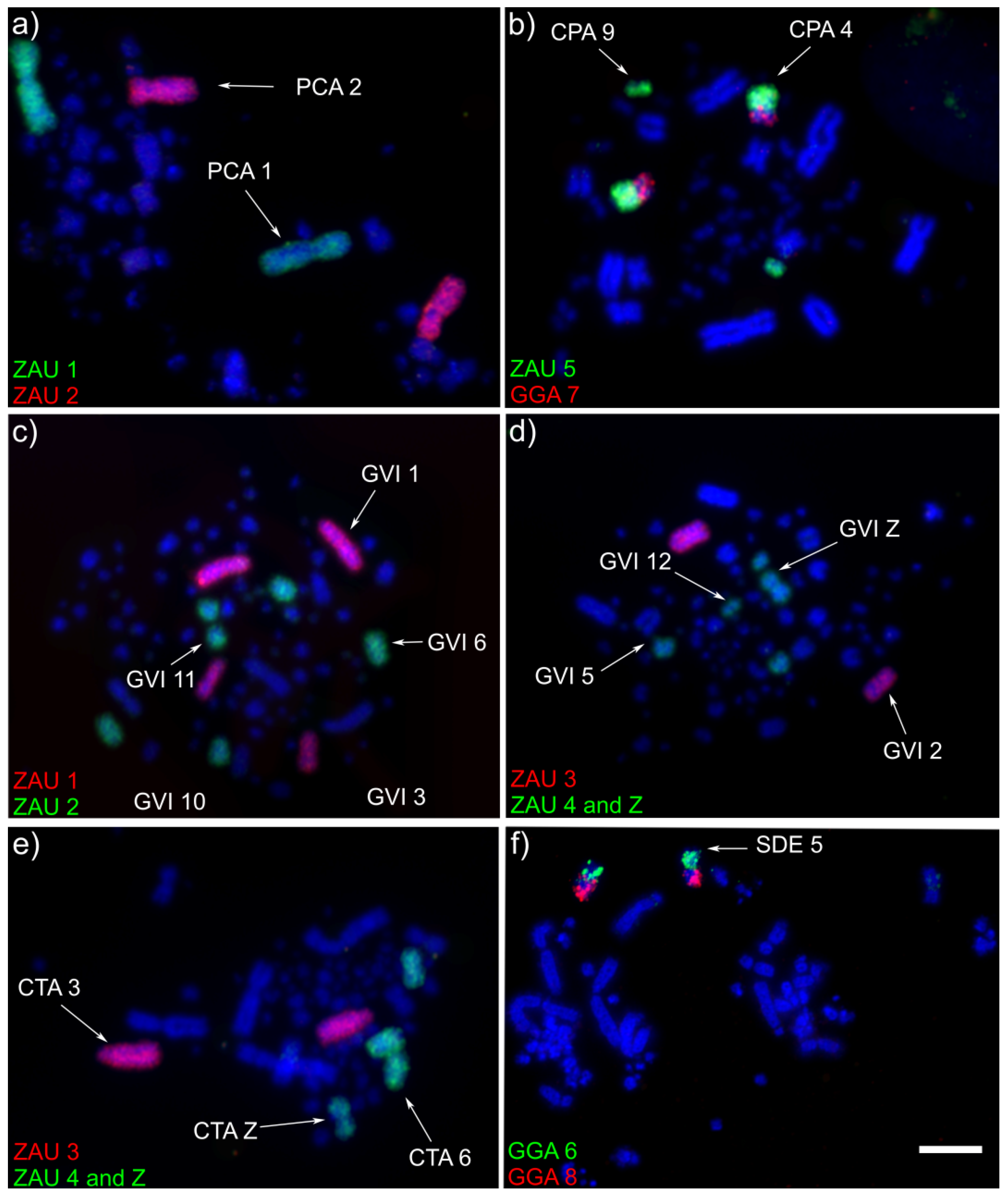
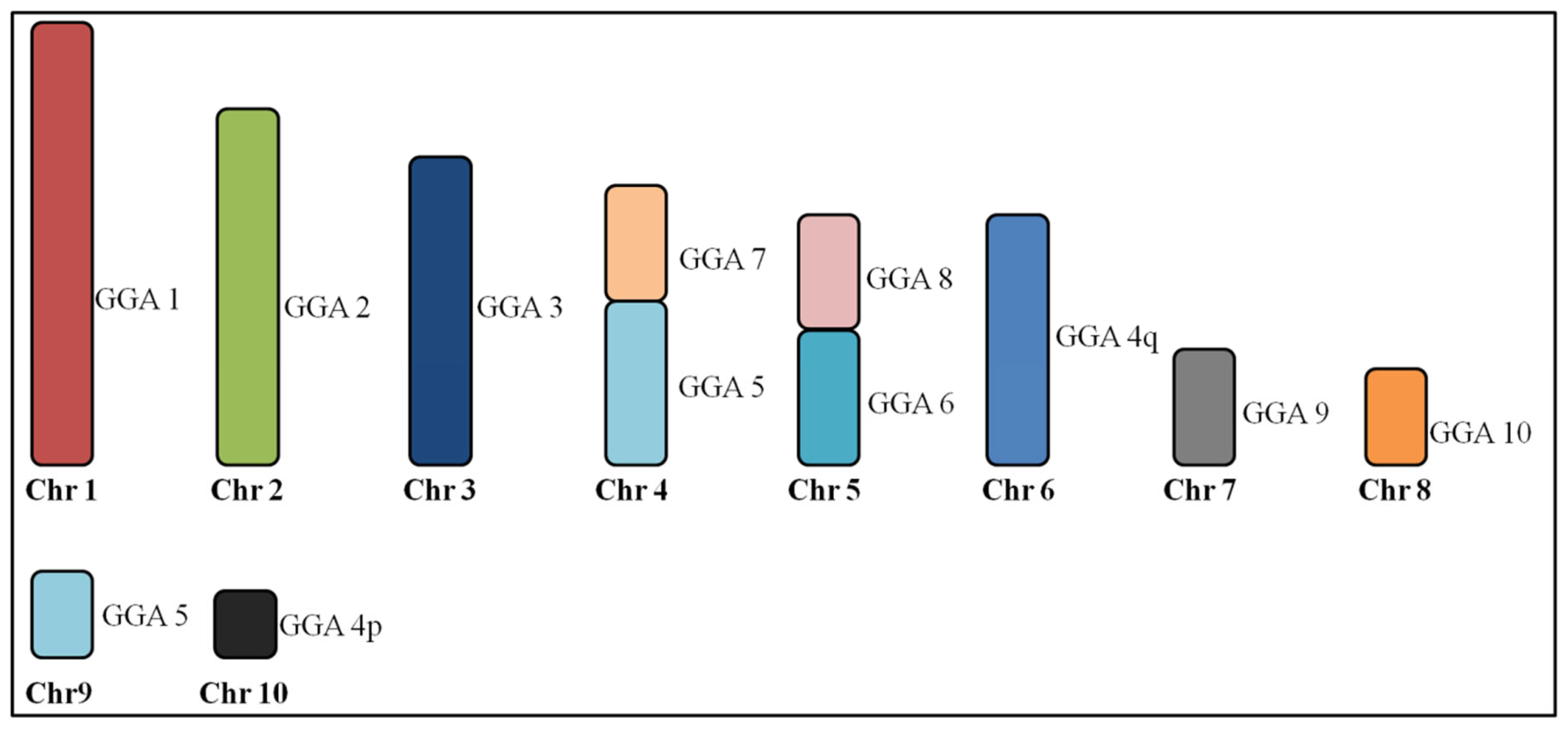
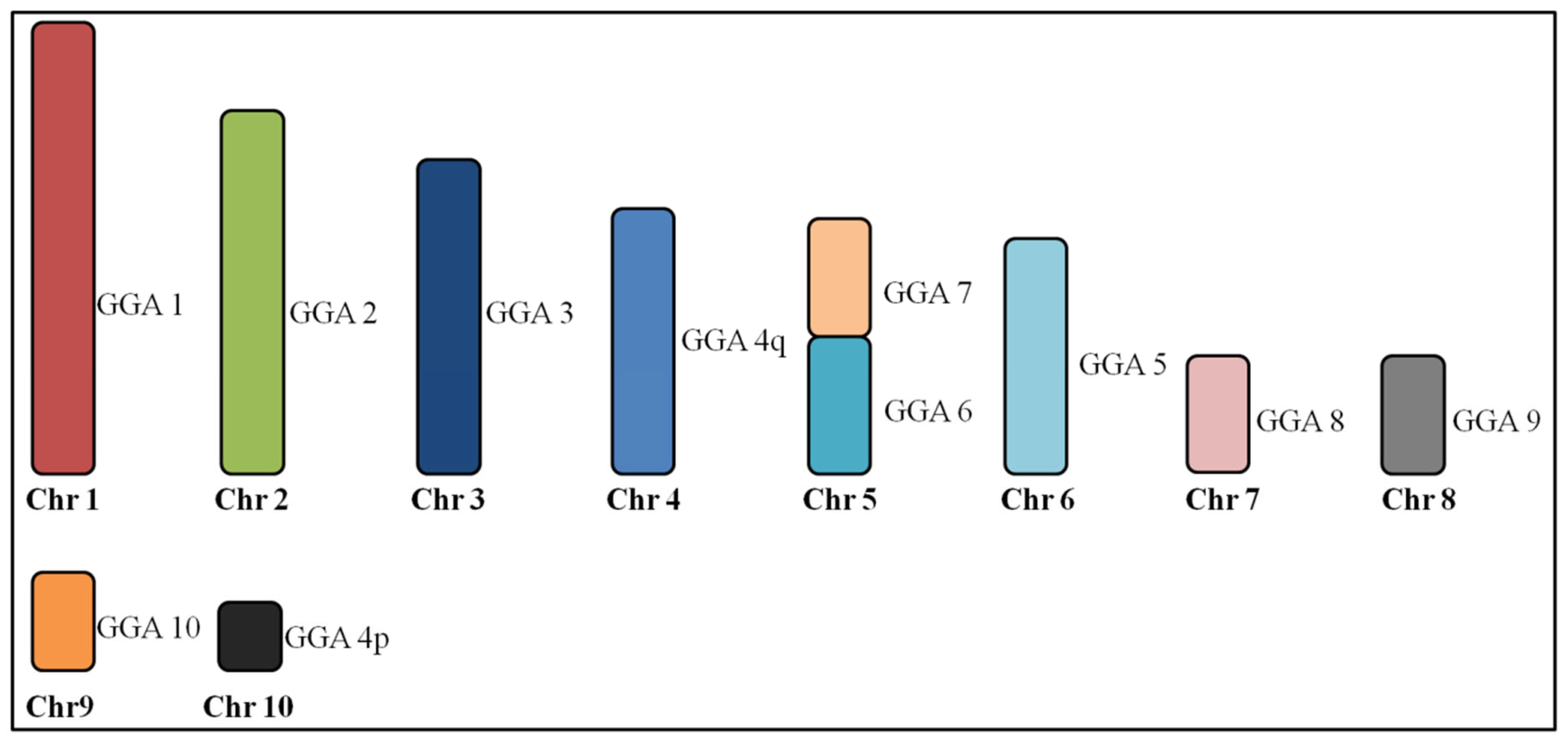
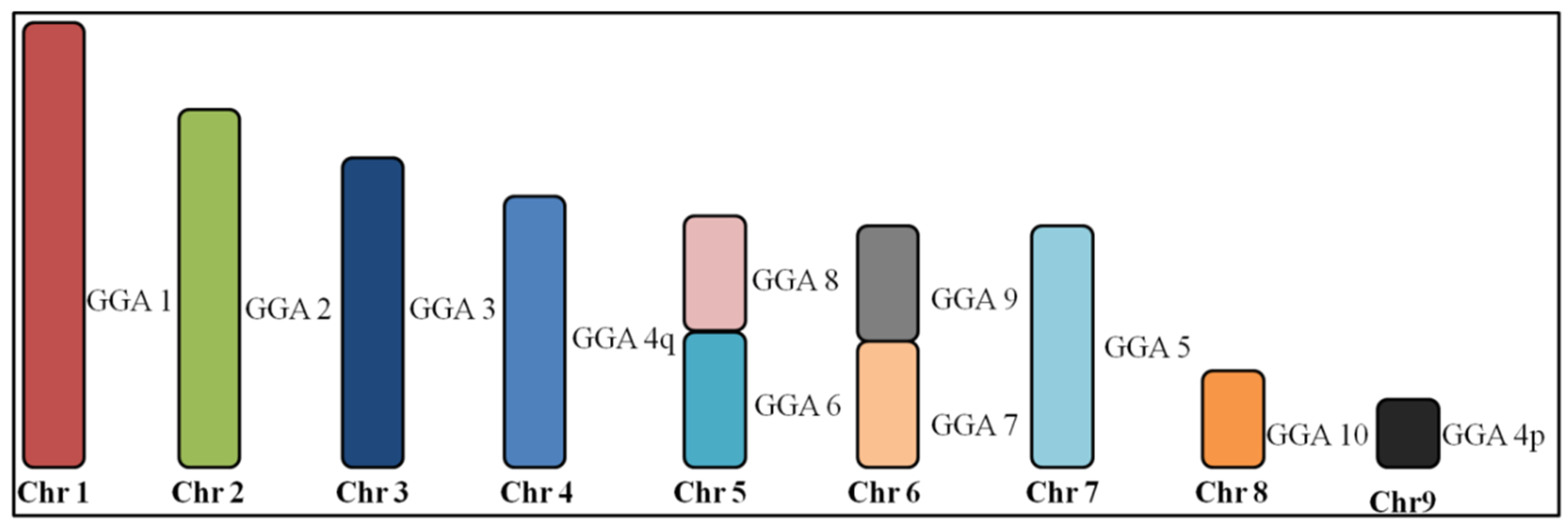
3.1. Macrochromosome Organization (GGA1-10)
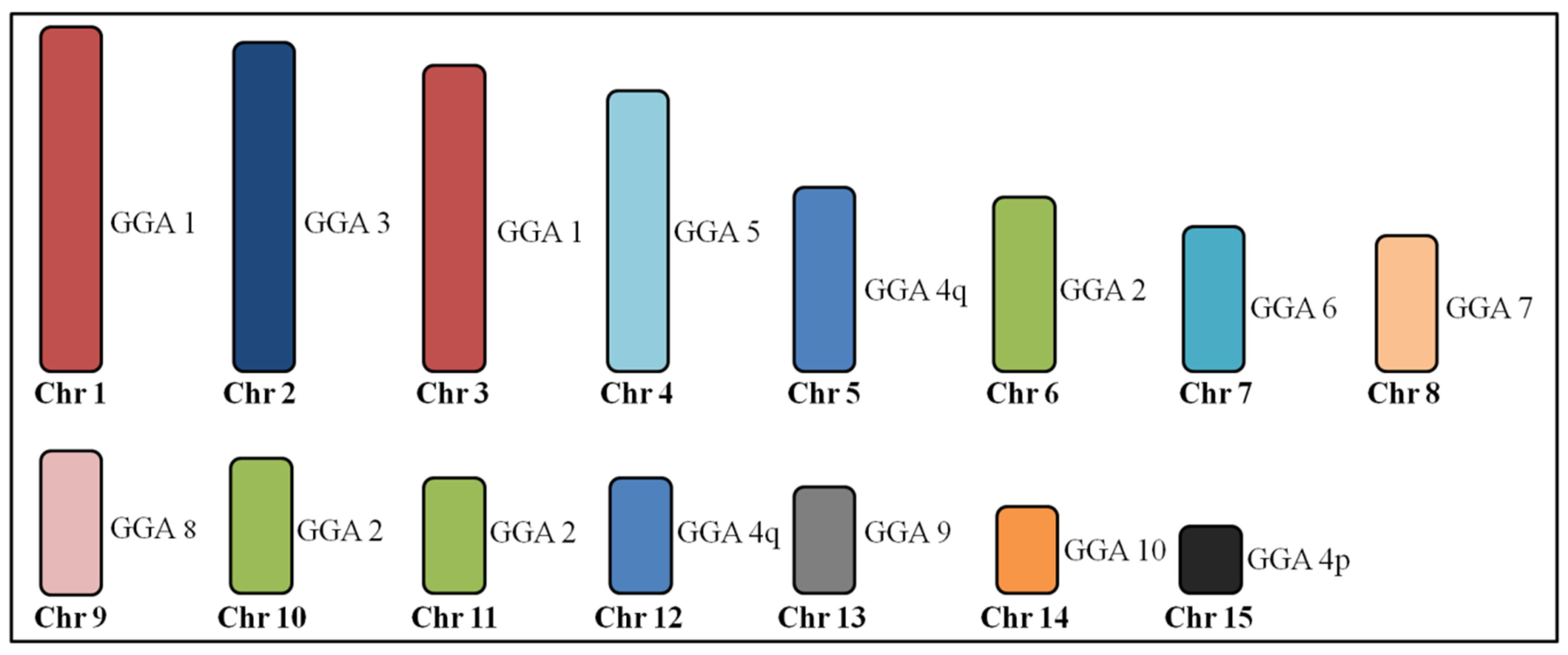
3.2. Microchromosome Organization (GGA11-28, Except GGA16)
3.3. G-Banding
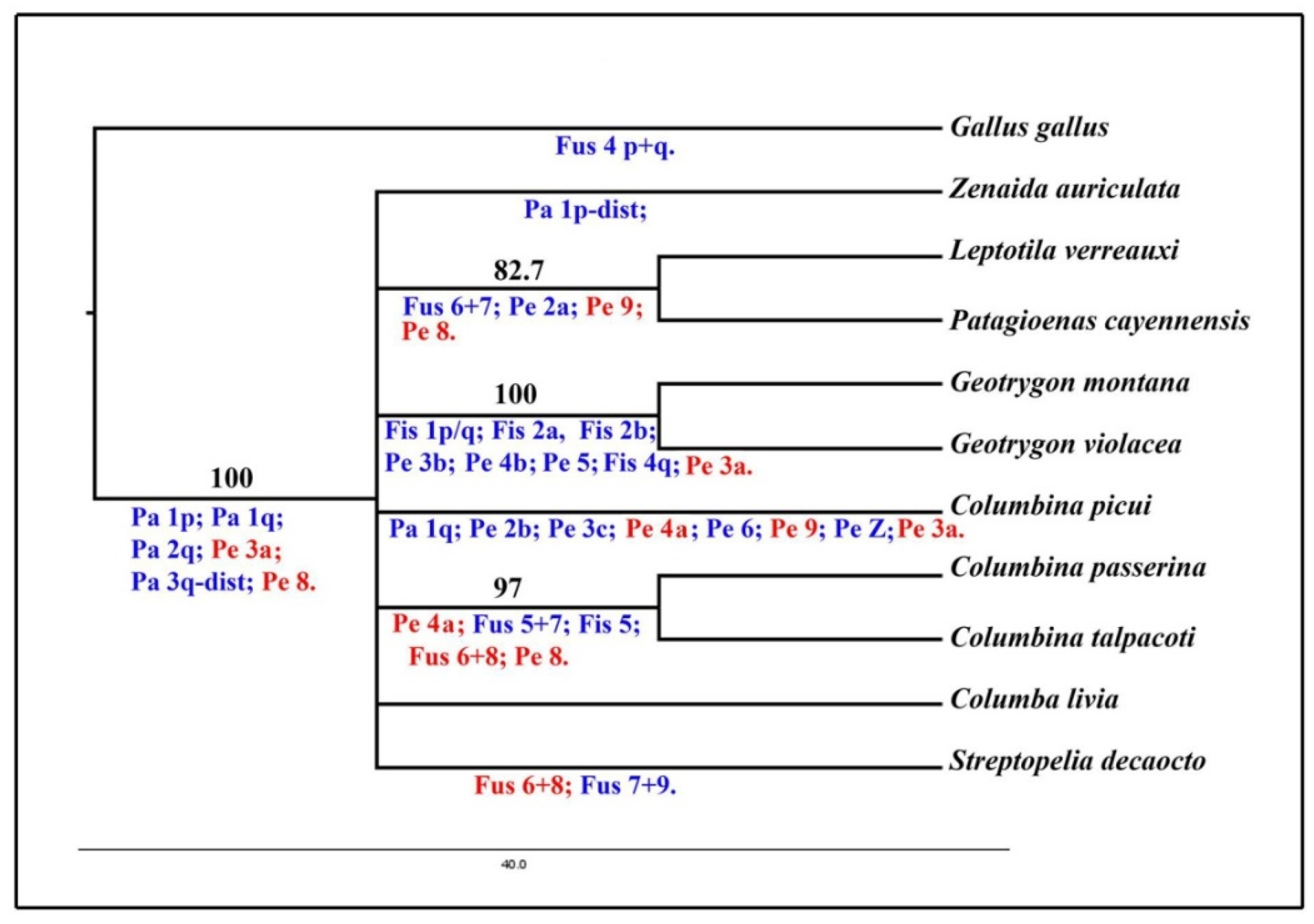
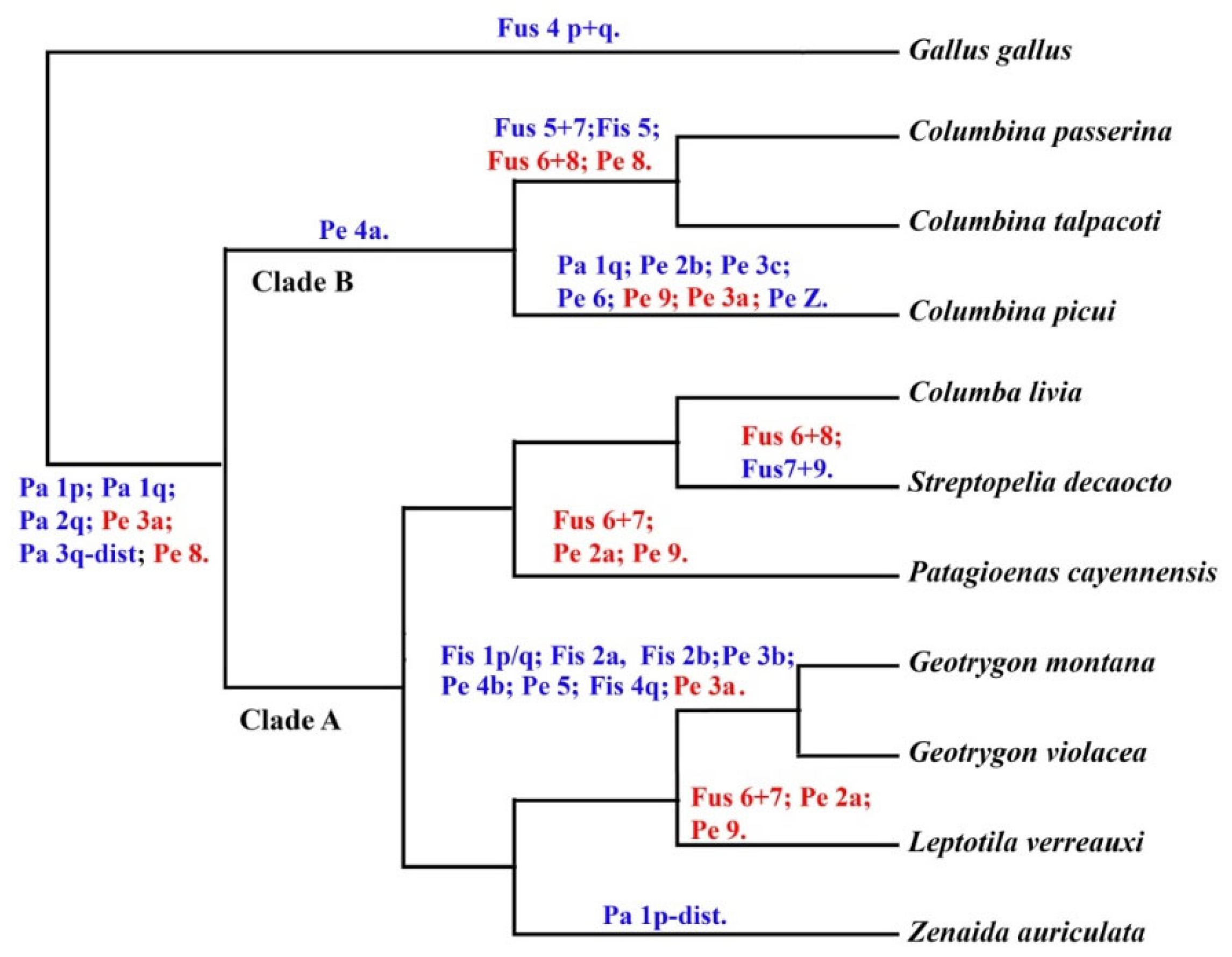
3.4. Phylogenetic Analysis
4. Discussion
4.1. Macrochromosome Organization
4.2. Conservation of Microchromosome Organization
4.3. Phylogenetic Relationships in the Family Columbidae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tegelstrom, H.; Ryttman, H. Chromosomes in birds (Aves): Evolutionary implications of macro- and microchromosome numbers and lengths. Hereditas 1981, 94, 225–233. [Google Scholar] [CrossRef]
- Kretschmer, R.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Karyotype evolution in birds: From conventional staining to chromosome painting. Genes 2018, 9, 181. [Google Scholar] [CrossRef] [Green Version]
- Hillier, L.W.; Miller, W.; Birney, E.; Warren, W.; Hardison, R.C.; Ponting, C.P.; Bork, P.; Burt, D.W.; Groenen, M.A.; Delany, M.E.; et al. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar]
- O’Connor, R.E.; Romanov, M.N.; Kiazim, L.G.; Barrett, P.M.; Farré, M.; Damas, J.; Ferguson-Smith, M.; Valenzuela, N.; Larkin, D.M.; Griffin, D.K. Reconstruction of the diapsid ancestral genome permits chromosome evolution tracing in avian and non-avian dinosaurs. Nat. Commun. 2018, 9, 1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farré, M.; Narayan, J.; Slavov, G.T.; Damas, J.; Auvil, L.; Li, C.; Jarvis, E.D.; Burt, D.W.; Griffin, D.K.; Larkin, D.M. Novel Insights Into Chromosome Evolution in Birds, Archosaurs, and Reptiles. Genome Biol. Evol. 2016, 8, 2442–2451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, D.; Barnes, E.; Cox, J.D. Pigeons and Doves: A Guide to the Pigeons and Doves of the World; Pica Press: Mountfield, UK, 2001. [Google Scholar]
- Pereira, S.L.; Johnson, K.P.; Clayton, D.H.; Baker, A.J. Mitochondrial and nuclear DNA sequences support a cretaceous origin of Columbiformes and a dispersal driven radiation in the paleogene. Syst. Biol. 2007, 56, 656–672. [Google Scholar] [CrossRef] [Green Version]
- Soares, A.E.R.; Novak, B.J.; Haile, J.; Heupink, T.H.; Fjeldså, J.; Gilbert, M.T.P.; Poinar, H.; Church, G.M.; Shapiro, B. Complete mitochondrial genomes of living and extinct pigeons revise the timing of the columbiform radiation. BMC Evol. Biol. 2016, 16, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sibley, C.G.; Ahlquist, J.E. Phylogeny and Classification of Birds: A Study in Molecular Evolution; Yale University Press: New Haven, CT, USA, 1990. [Google Scholar]
- Shapiro, B.; Sibthorpe, D.; Rambaut, A.; Austin, J.; Wragg, G.M.; Bininda-Emonds, O.R.; Lee, P.L.; Cooper, A. Flight of the dodo. Science 2002, 295, 1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lucca, E.J.; de Aguiar, M.L.R. Chromosomal evolution in Columbiformes (Aves). Caryologia 1976, 29, 59–68. [Google Scholar] [CrossRef] [Green Version]
- De Lucca, E.J. Chromosomal evolution of South American Columbiformes (Aves). Genetica 1984, 62, 177–185. [Google Scholar] [CrossRef]
- Guttenbach, M.; Nanda, I.; Feichtinger, W.; Masabanda, J.S.; Griffin, D.K.; Schmid, M. Comparative chromosome painting of chicken autosomal paints 1–9 in nine different bird species. Cytogenet. Genome Res. 2003, 103, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Derjusheva, S.; Kurganova, A.; Haberman, F.; Gaginskaia, E. High chromosome conservation detected by comparative chromosome painting in chicken, pigeon and passerine birds. Chromosome Res. 2004, 12, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; de Oliveira, T.D.; Furo, I.O.; Silva, F.A.O.; Gunski, R.J.; Garnero, A.D.V.; Cioffi, M.B.; de Oliveira, E.H.C.; de Freitas, T.R.O. Repetitive DNAs and shrink genomes: A chromosomal analysis in nine Columbidae species (Aves, Columbiformes). Genet. Mol. Biol. 2018, 41, 98–106. [Google Scholar] [CrossRef]
- Kretschmer, R.; Furo, I.O.; Gunski, R.J.; Garnero, A.D.V.; Pereira, J.C.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C.; de Freitas, T.R.O. Comparative chromosome painting in Columbidae (Columbiformes) reinforces divergence in Passerea and Columbea. Chromosome Res. 2018, 26, 211–223. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, R.E.; Kiazim, L.; Skinner, B.; Fonseka, G.; Joseph, S.; Jennings, R.; Larkin, D.M.; Griffin, D.K. Patterns of microchromosome organization remain highly conserved throughout avian evolution. Chromosoma 2019, 128, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, M.; Ikeuchi, T.; Maino, S. A feather pulp culture for avian chromosomes with notes on the chromosomes of the peafowl and the ostrich. Experientia 1968, 24, 1923–1929. [Google Scholar] [CrossRef]
- Joseph, S.; O’Connor, R.E.; Al Mutery, A.F.; Watson, M.; Larkin, D.M.; Griffin, D.K. Chromosome Level Genome Assembly and Comparative Genomics between Three Falcon Species Reveals an Unusual Pattern of Genome Organisation. Diversity 2018, 10, 113. [Google Scholar] [CrossRef] [Green Version]
- Dobigny, G.; Ozouf-Costaz, C.; Bonillo, C.; Volouev, V. Viability of X-autosome translocations in mammals: An epigenomic hypothesis from a rodent case-study. Chromosoma 2004, 113, 34–41. [Google Scholar] [CrossRef]
- Swoford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Degrandi, T.M.; Barcelos, S.A.; Costa, A.L.; Garnero, A.D.V.; Hass, I.; Gunski, R.J. Introducing the Bird Chromosome Database: An overview of cytogenetic studies on birds. Cytogenet. Genome Res. 2020, in press. [Google Scholar] [CrossRef]
- Shibusawa, M.; Nishibori, M.; Nishida-Umehara, C.; Tsudzuk, M.; Masaband, J.; Griffin, D.K.; Matsuda, Y. Karyotypic evolution in the Galliformes: An examination of the process of karyotypic evolution by comparison of the molecular cytogenetic findings with the molecular phylogeny. Cytogenet. Genome Res. 2004, 106, 111–119. [Google Scholar] [CrossRef]
- Nishida, C.; Ishijima, J.; Kosaka, A.; Tanabe, H.; Habermann, F.A.; Griffin, D.K.; Matsuda, Y. Characterization of chromosome structures of Falconinae (Falconidae, Falconiformes, Aves) by chromosome painting and delineation of chromosome rearrangements during their differentiation. Chromosome Res. 2008, 16, 171–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, M.S.; Furo, I.O.; Tagliarini, M.M.; Kretschmer, R.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. The Karyotype of the Hoatzin (Opisthocomus hoazin)—A Phylogenetic Enigma of the Neornithes. Cytogenet. Genome Res. 2018, 156, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Lapiedra, O.; Sol, D.; Carranza, S.; Beaulieu, J.M. Behavioural changes and the adaptive diversification of pigeons and doves. Proc. R. Soc. B 2013, 280, 20122893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, R.E.; Farré, M.; Joseph, S.; Damas, J.; Kiazim, L.; Jennings, R.; Bennett, S.; Slack, E.A.; Allanson, E.; Larkin, D.M.; et al. Chromosome-level assembly reveals extensive rearrangement in saker falcon and budgerigar, but not ostrich, genomes. Genome Biol. 2018, 19, 171. [Google Scholar] [CrossRef] [Green Version]
- Nanda, I.; Benisch, P.; Fetting, D.; Haaf, T.; Schmid, M. Synteny conservation of chicken macrochromosomes 1–10 in different Avian lineages revealed by cross-species chromosome painting. Cytogenet. Genome Res. 2011, 132, 165–181. [Google Scholar] [CrossRef]
- De Oliveira, E.H.; de Moura, S.P.; dos Anjos, L.J.; Nagamachi, C.Y.; Pieczarka, J.C.; O’Brien, P.C.; Ferguson-Smith, M.A. Comparative chromosome painting between chicken and spectacled owl (Pulsatrix perspicillata): Implications for chromosomal evolution in the Strigidae (Aves, Strigiformes). Cytogenet. Genome Res. 2008, 122, 157–162. [Google Scholar] [CrossRef]
- Degrandi, T.M.; Garnero, A.D.V.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Kretschmer, R.; de Oliveira, E.H.C.; Gunski, R.J. Chromosome Painting in Trogon s. surrucura (Aves, Trogoniformes) Reveals a Karyotype Derived by Chromosomal Fissions, Fusions, and Inversions. Cytogenet. Genome Res. 2017, 151, 208–215. [Google Scholar]
- Nanda, I.; Karl, E.; Griffin, D.K.; Schartl, M.; Schmid, M. Chromosome repatterning in three representative parrots (Psittaciformes) inferred from comparative chromosome painting. Cytogenet. Genome Res. 2007, 117, 43–53. [Google Scholar] [CrossRef]
- Ellegren, H. Evolutionary stasis: The stable chromosomes of birds. Trends Ecol. Evol. 2010, 25, 283–291. [Google Scholar]
- Nie, W.; O’Brien, P.C.M.; Fu, B.; Wang, J.; Su, W.; He, K.; Bed’Hom, B.; Volobouev, V.; Ferguson-Smith, M.A.; Dobigny, G.; et al. Multidirectional chromosome painting substantiates the occurrence of extensive genomic reshuffling within Accipitriformes. BMC Evol. Biol. 2015, 15, 205. [Google Scholar] [CrossRef] [Green Version]
- Sweet, A.D.; Johnson, K.P. Patterns of diversification in small New World ground doves are consistent with major geologic events. Auk 2014, 132, 300–312. [Google Scholar] [CrossRef]
- Warren, W.C.; Clayton, D.F.; Ellegren, H.; Arnold, A.P.; Hillier, L.W.; Künstner, A.; Searle, S.; White, S.; Vilella, A.J.; Fairley, S.; et al. The genome of a songbird. Nature 2010, 464, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Damas, J.; O’Connor, R.; Farré, M.; Lenis, V.P.E.; Martell, H.J.; Mandawala, A.; Fowler, K.E.; Jospeh, S.; Swain, M.; Griffin, D.K.; et al. Upgrading short-read animal genome assemblies to chromosome level using comparative genomics and a universal probe set. Genome Res. 2017, 27, 875–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Number of Individuals/Sex | 2n | Wcp | Micro BACs | G-Banding |
|---|---|---|---|---|---|
| Columbina talpacoti | 2 M | 76 | Present study | Present study | Present study |
| Columbina passerina | 1 M | 76 | Present study | Present study | Present study |
| Columbina picui | 1 M and 1 F | 76 | [16] | - | Present study |
| Columba livia | 1 M | 80 | [16] | [17] | Present study |
| Geotrygon montana | 1 M | 86 | Present study | Present study | Present study |
| Geotrygon violacea | 1 F | 86 | Present study | Present study | Present study |
| Leptotila verreauxi | 2 M | 78 | [16] | - | Present study |
| Patagioenas cayennensis | 2 M | 76 | Present study | Present study | Present study |
| Streptopelia decaocto | 1 F | 76 | Present study | [17] | Present study |
| Zenaida auriculata | 2 M | 76 | [16] | - | Present study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kretschmer, R.; Furo, I.d.O.; Gomes, A.J.B.; Kiazim, L.G.; Gunski, R.J.; Garnero, A.d.V.; Pereira, J.C.; Ferguson-Smith, M.A.; de Oliveira, E.H.C.; Griffin, D.K.; et al. A Comprehensive Cytogenetic Analysis of Several Members of the Family Columbidae (Aves, Columbiformes). Genes 2020, 11, 632. https://doi.org/10.3390/genes11060632
Kretschmer R, Furo IdO, Gomes AJB, Kiazim LG, Gunski RJ, Garnero AdV, Pereira JC, Ferguson-Smith MA, de Oliveira EHC, Griffin DK, et al. A Comprehensive Cytogenetic Analysis of Several Members of the Family Columbidae (Aves, Columbiformes). Genes. 2020; 11(6):632. https://doi.org/10.3390/genes11060632
Chicago/Turabian StyleKretschmer, Rafael, Ivanete de Oliveira Furo, Anderson José Baia Gomes, Lucas G. Kiazim, Ricardo José Gunski, Analía del Valle Garnero, Jorge C. Pereira, Malcolm A. Ferguson-Smith, Edivaldo Herculano Corrêa de Oliveira, Darren K. Griffin, and et al. 2020. "A Comprehensive Cytogenetic Analysis of Several Members of the Family Columbidae (Aves, Columbiformes)" Genes 11, no. 6: 632. https://doi.org/10.3390/genes11060632
APA StyleKretschmer, R., Furo, I. d. O., Gomes, A. J. B., Kiazim, L. G., Gunski, R. J., Garnero, A. d. V., Pereira, J. C., Ferguson-Smith, M. A., de Oliveira, E. H. C., Griffin, D. K., de Freitas, T. R. O., & O’Connor, R. E. (2020). A Comprehensive Cytogenetic Analysis of Several Members of the Family Columbidae (Aves, Columbiformes). Genes, 11(6), 632. https://doi.org/10.3390/genes11060632