Knockout of Pi21 by CRISPR/Cas9 and iTRAQ-Based Proteomic Analysis of Mutants Revealed New Insights into M. oryzae Resistance in Elite Rice Line

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Material Used and Experimental Conditions
2.2. Construction of Vector and Rice Transformation
2.3. Genotyping, Screening of Transgene-free Plants, and Off-target Assessment
2.4. Pathogen Inoculation and Disease Scoring
2.5. Phenotyping of Mutant Lines
2.6. Protein Extraction and Digestion
2.7. iTRAQ Labeling and Mass Spectrophotometry Analysis
2.8. Data Analysis
2.9. RT–qPCR-Based Validation of Proteomic Data
2.10. Statistical Analysis
3. Results
3.1. Construction of Vector
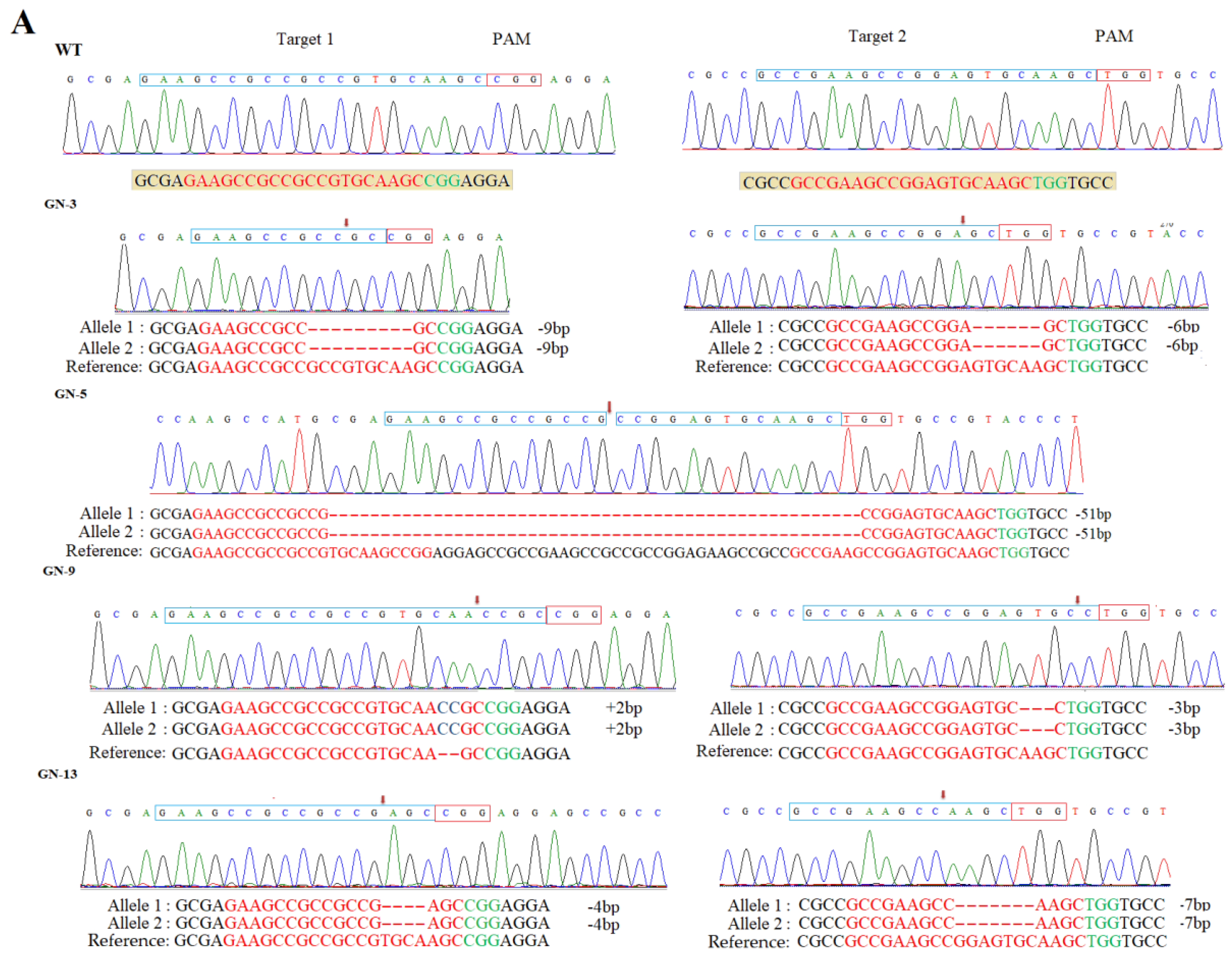
3.2. Mutation Frequency, Off-Target Effects, and Segregation of Mutants
3.3. T-DNA-Free Plants in T1 Generation
3.4. Resistance to Pathogen in Pi21 Mutants
3.5. Agronomic Performance Assessment of Wild Type and Mutant Lines
3.6. Proteins Extraction and Quantification
3.7. Functional Networks of DEPs
3.8. Gene Ontology (GO) Annotation of DEPs
3.9. Pathway Enrichment Analysis
3.10. Identification of Hub-Proteins
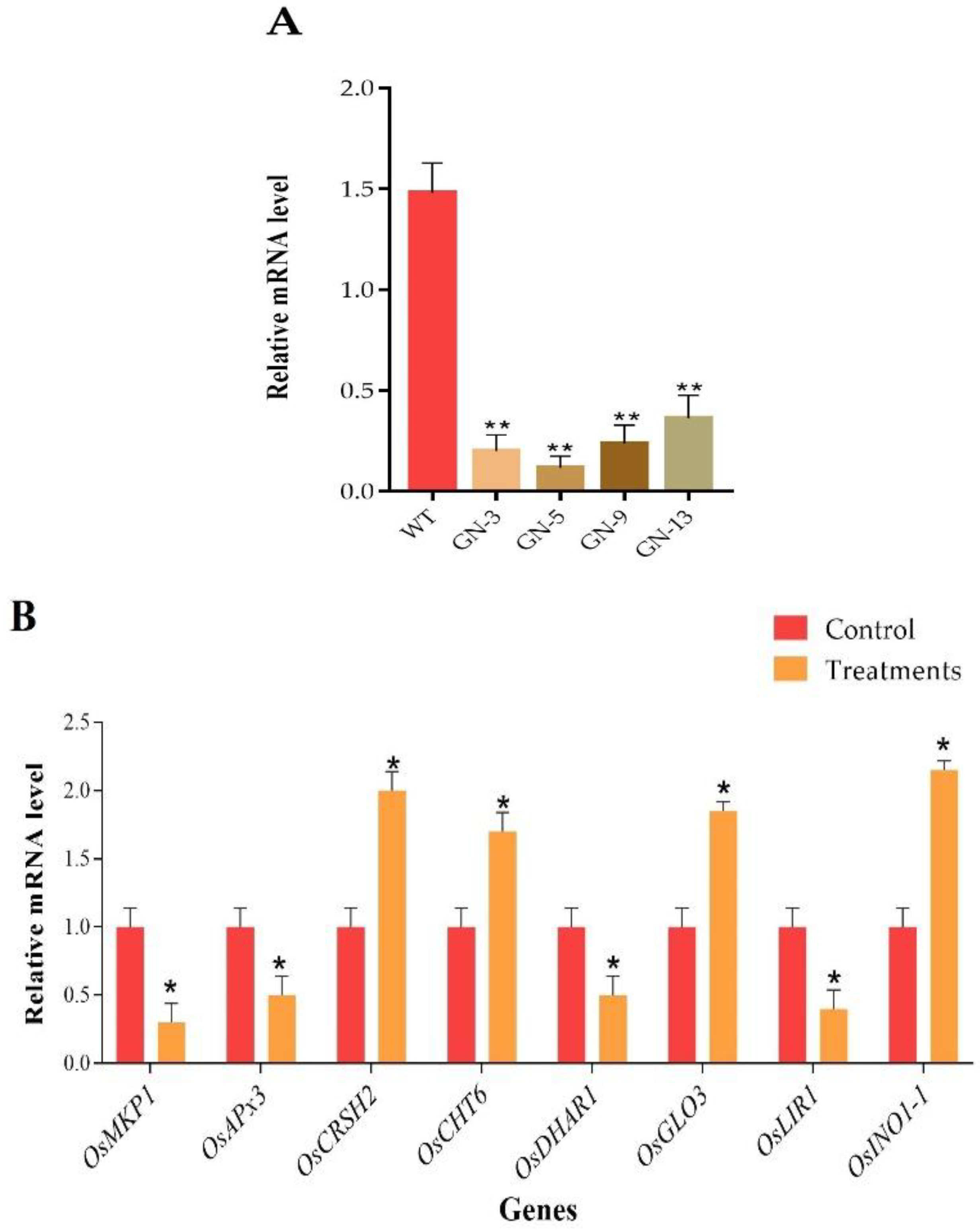
3.11. RT-qPCR-Based Analysis of CRISPR Mutants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khush, G.S.; Jena, K. Current status and future prospects for research on blast resistance in rice (Oryza sativa L.). In Advances in Genetics, Genomics and Control of Rice Blast Disease; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–10. [Google Scholar]
- Dean, R.A.; Talbot, N.J.; Ebbole, D.J.; Farman, M.L.; Mitchell, T.K.; Orbach, M.J.; Thon, M.; Kulkarni, R.; Xu, J.-R.; Pan, H. The genome sequence of the rice blast fungus Magnaporthe grisea. Nature 2005, 434, 980. [Google Scholar] [CrossRef] [PubMed]
- Skamnioti, P.; Gurr, S.J. Against the grain: Safeguarding rice from rice blast disease. Trends Biotechnol. 2009, 27, 141–150. [Google Scholar] [CrossRef]
- Boyd, L.A.; Ridout, C.; O’Sullivan, D.M.; Leach, J.E.; Leung, H. Plant–pathogen interactions: Disease resistance in modern agriculture. Trends Genet. 2013, 29, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, S.; Yamamoto, S.-I.; Mizobuchi, R.; Yamanouchi, U.; Ono, K.; Kitazawa, N.; Yasuda, N.; Fujita, Y.; Nguyen, T.T.T.; Koizumi, S. Multiple functional polymorphisms in a single disease resistance gene in rice enhance durable resistance to blast. Sci. Rep. 2014, 4, 4550. [Google Scholar] [CrossRef]
- Fukuoka, S.; Saka, N.; Koga, H.; Ono, K.; Shimizu, T.; Ebana, K.; Hayashi, N.; Takahashi, A.; Hirochika, H.; Okuno, K. Loss of function of a proline-containing protein confers durable disease resistance in rice. Science 2009, 325, 998–1001. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-guided human genome engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef]
- Li, D.; Qiu, Z.; Shao, Y.; Chen, Y.; Guan, Y.; Liu, M.; Li, Y.; Gao, N.; Wang, L.; Lu, X. Heritable gene targeting in the mouse and rat using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 681. [Google Scholar] [CrossRef]
- Yang, H.; Wang, H.; Jaenisch, R. Generating genetically modified mice using CRISPR/Cas-mediated genome engineering. Nat. Protoc. 2014, 9, 1956. [Google Scholar] [CrossRef] [PubMed]
- Fujii, W.; Onuma, A.; Sugiura, K.; Naito, K. Efficient generation of genome-modified mice via offset-nicking by CRISPR/Cas system. Biochem. Biophys. Res. Commun. 2014, 445, 791–794. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.W.; Kim, S.; Kim, J.M.; Kim, J.-S. Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 230. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Regan, S.N.; Xia, Y.; Oostrom, L.A.; Cowan, C.A.; Musunuru, K. Enhanced efficiency of human pluripotent stem cell genome editing through replacing TALENs with CRISPRs. Cell Stem Cell 2013, 12, 393–394. [Google Scholar] [CrossRef]
- Han, Y.; Luo, D.J.; Usman, B.; Nawaz, G.; Zhao, N.; Liu, F.; Li, R.B. Development of High Yielding Glutinous Cytoplasmic Male Sterile Rice (Oryza sativa L.) Lines through CRISPR/Cas9 Based Mutagenesis of Wx and TGW6 and Proteomic Analysis of Anther. Agronomy 2018, 8, 290. [Google Scholar] [CrossRef]
- Nawaz, G.; Han, Y.; Usman, B.; Liu, F.; Qin, B.; Li, R. Knockout of OsPRP1, a gene encoding proline-rich protein, confers enhanced cold sensitivity in rice (Oryza sativa L.) at the seedling stage. 3 Biotech 2019, 9, 254. [Google Scholar] [CrossRef]
- Li, M.; Li, X.; Zhou, Z.; Wu, P.; Fang, M.; Pan, X.; Lin, Q.; Luo, W.; Wu, G.; Li, H. Reassessment of the four yield-related genes Gn1a, DEP1, GS3, and IPA1 in rice using a CRISPR/Cas9 system. Front. Plant Sci. 2016, 7, 377. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, H.; Xu, N.; Zhang, B.; Gou, F.; Zhu, J.-K. Application of the CRISPR–Cas system for efficient genome engineering in plants. Mol. Plant 2013, 6, 2008–2011. [Google Scholar] [CrossRef]
- Jiang, W.; Yang, B.; Weeks, D.P. Efficient CRISPR/Cas9-mediated gene editing in Arabidopsis thaliana and inheritance of modified genes in the T2 and T3 generations. PLoS ONE 2014, 9, e99225. [Google Scholar] [CrossRef]
- Feng, Z.; Mao, Y.; Xu, N.; Zhang, B.; Wei, P.; Yang, D.-L.; Wang, Z.; Zhang, Z.; Zheng, R.; Yang, L. Multigeneration analysis reveals the inheritance, specificity, and patterns of CRISPR/Cas-induced gene modifications in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 4632–4637. [Google Scholar] [CrossRef]
- Gao, J.; Wang, G.; Ma, S.; Xie, X.; Wu, X.; Zhang, X.; Wu, Y.; Zhao, P.; Xia, Q. CRISPR/Cas9-mediated targeted mutagenesis in Nicotiana tabacum. Plant Mol. Biol. 2015, 87, 99–110. [Google Scholar] [CrossRef]
- Xing, H.-L.; Dong, L.; Wang, Z.-P.; Zhang, H.-Y.; Han, C.-Y.; Liu, B.; Wang, X.-C.; Chen, Q.-J. A CRISPR/Cas9 toolkit for multiplex genome editing in plants. BMC Plant Biol. 2014, 14, 327. [Google Scholar] [CrossRef] [PubMed]
- Svitashev, S.; Young, J.K.; Schwartz, C.; Gao, H.; Falco, S.C.; Cigan, A.M. Targeted mutagenesis, precise gene editing, and site-specific gene insertion in maize using Cas9 and guide RNA. Plant Physiol. 2015, 169, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, T.B.; LaFayette, P.R.; Schmitz, R.J.; Parrott, W.A. Targeted genome modifications in soybean with CRISPR/Cas9. BMC Biotechnol. 2015, 15, 16. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, Z.B.; Xing, A.; Moon, B.P.; Koellhoffer, J.P.; Huang, L.; Ward, R.T.; Clifton, E.; Falco, S.C.; Cigan, A.M. Cas9-Guide RNA Directed Genome Editing in Soybean. Plant Physiol. 2015, 169, 960–970. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Minkenberg, B.; Yang, Y. Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system. Proc. Natl. Acad. Sci. USA 2015, 112, 3570–3575. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, Q.; Zhu, Q.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.; Li, H.; Lin, Y.; et al. A Robust CRISPR/Cas9 System for Convenient, High-Efficiency Multiplex Genome Editing in Monocot and Dicot Plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef]
- Usman, B.; Nawaz, G.; Zhao, N.; Liu, Y.; Li, R. Generation of High Yielding and Fragrant Rice (Oryza sativa L.) Lines by CRISPR/Cas9 Targeted Mutagenesis of Three Homoeologs of Cytochrome P450 Gene Family and OsBADH2 and Transcriptome and Proteome Profiling of Revealed Changes Triggered by Mutations. Plants 2020, 9, 788. [Google Scholar] [CrossRef]
- Bikard, D.; Jiang, W.; Samai, P.; Hochschild, A.; Zhang, F.; Marraffini, L.A. Programmable repression and activation of bacterial gene expression using an engineered CRISPR-Cas system. Nucleic Acids Res. 2013, 41, 7429–7437. [Google Scholar] [CrossRef]
- Wang, T.; Wei, J.J.; Sabatini, D.M.; Lander, E.S. Genetic screens in human cells using the CRISPR-Cas9 system. Science 2014, 343, 80–84. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, B.; Weeks, D.P.; Spalding, M.H.; Yang, B. Large chromosomal deletions and heritable small genetic changes induced by CRISPR/Cas9 in rice. Nucleic Acids Res. 2014, 42, 10903–10914. [Google Scholar] [CrossRef] [PubMed]
- Domżalska, L.; Kędracka-Krok, S.; Jankowska, U.; Grzyb, M.; Sobczak, M.; Rybczyński, J.J.; Mikuła, A. Proteomic analysis of stipe explants reveals differentially expressed proteins involved in early direct somatic embryogenesis of the tree fern Cyathea delgadii Sternb. Plant Sci. 2017, 258, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.; Noirel, J.; Ow, S.Y.; Salim, M.; Pereira-Medrano, A.G.; Couto, N.; Pandhal, J.; Smith, D.; Pham, T.K.; Karunakaran, E. An insight into iTRAQ: Where do we stand now? Anal. Bioanal. Chem. 2012, 404, 1011–1027. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.X.; Luo, Y.M.; Ye, Z.Q.; Cao, X.; Liang, J.N.; Wang, Q.; Wu, Y.; Wu, J.H.; Wang, H.Y.; Zhang, M.; et al. iTRAQ-based proteomics analysis of autophagy-mediated immune responses against the vascular fungal pathogen Verticillium dahliae in Arabidopsis. Autophagy 2018, 14, 598–618. [Google Scholar] [CrossRef]
- Wang, J.; Islam, F.; Li, L.; Long, M.; Yang, C.; Jin, X.; Ali, B.; Mao, B.; Zhou, W. Complementary RNA-Sequencing Based Transcriptomics and iTRAQ Proteomics Reveal the Mechanism of the Alleviation of Quinclorac Stress by Salicylic Acid in Oryza sativa ssp. japonica. Int. J. Mol. Sci. 2017, 18, 1975. [Google Scholar] [CrossRef]
- Chen, S.; Chen, J.; Hou, F.; Feng, Y.; Zhang, R. iTRAQ-based quantitative proteomic analysis reveals the lateral meristem developmental mechanism for branched spike development in tetraploid wheat (Triticum turgidum L.). BMC Genom. 2018, 19, 228. [Google Scholar] [CrossRef]
- Bu, T.-t.; Shen, J.; Chao, Q.; Shen, Z.; Yan, Z.; Zheng, H.-y.; Wang, B.-c. Dynamic N-glycoproteome analysis of maize seedling leaves during de-etiolation using Concanavalin A lectin affinity chromatography and a nano-LC–MS/MS-based iTRAQ approach. Plant Cell Rep. 2017, 36, 1943–1958. [Google Scholar] [CrossRef]
- Lin, J.; Wong, K.-C. Off-target predictions in CRISPR-Cas9 gene editing using deep learning. Bioinformatics 2018, 34, i656–i663. [Google Scholar] [CrossRef]
- Akcakaya, P.; Bobbin, M.L.; Guo, J.A.; Malagon-Lopez, J.; Clement, K.; Garcia, S.P.; Fellows, M.D.; Porritt, M.J.; Firth, M.A.; Carreras, A. In vivo CRISPR editing with no detectable genome-wide off-target mutations. Nature 2018, 561, 416. [Google Scholar] [CrossRef]
- Liang, G.; Zhang, H.; Lou, D.; Yu, D. Selection of highly efficient sgRNAs for CRISPR/Cas9-based plant genome editing. Sci. Rep. 2016, 6, 21451. [Google Scholar] [CrossRef]
- Ma, X.; Chen, L.; Zhu, Q.; Chen, Y.; Liu, Y.G. Rapid Decoding of Sequence-Specific Nuclease-Induced Heterozygous and Biallelic Mutations by Direct Sequencing of PCR Products. Mol. Plant 2015, 8, 1285–1287. [Google Scholar] [CrossRef]
- Ma, X.; Liu, Y.G. CRISPR/Cas9-Based Multiplex Genome Editing in Monocot and Dicot Plants. Curr. Protoc. Mol. Biol. 2016, 115, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 1994, 6, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Kawasaki, S.; Fujimura, T.; Wang, C.T. A protocol for high-throughput extraction of DNA from rice leaves. Plant Mol. Biol. Rep. 2005, 23, 291–295. [Google Scholar] [CrossRef]
- Tian, D.; Chen, Z.; Chen, Z.; Zhou, Y.; Wang, Z.; Wang, F.; Chen, S. Allele-specific marker-based assessment revealed that the rice blast resistance genes Pi2 and Pi9 have not been widely deployed in Chinese indica rice cultivars. Rice 2016, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Mackill, D.; Bonman, J. Inheritance of blast resistance in near-isogenic lines of rice. Phytopathology 1992, 82, 746–749. [Google Scholar] [CrossRef]
- Chen, L.; Huang, Y.; Xu, M.; Cheng, Z.; Zhang, D.; Zheng, J. iTRAQ-Based Quantitative Proteomics Analysis of Black Rice Grain Development Reveals Metabolic Pathways Associated with Anthocyanin Biosynthesis. PLoS ONE 2016, 11, e0159238. [Google Scholar] [CrossRef]
- Yang, L.; Ji, J.; Harris-Shultz, K.R.; Wang, H.; Wang, H.; Abd-Allah, E.F.; Luo, Y.; Hu, X. The dynamic changes of the plasma membrane proteins and the protective roles of nitric oxide in rice subjected to heavy metal cadmium stress. Front. Plant Sci. 2016, 7, 190. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Li, T.; Liu, B.; Spalding, M.H.; Weeks, D.P.; Yang, B. High-efficiency TALEN-based gene editing produces disease-resistant rice. Nat. Biotechnol. 2012, 30, 390. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.-L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-A.; Moon, H.; Park, C.-J. CRISPR/Cas9-targeted mutagenesis of Os8N3 in rice to confer resistance to Xanthomonas oryzae pv. oryzae. Rice 2019, 12, 1–13. [Google Scholar]
- Han, Y.; Teng, K.; Nawaz, G.; Feng, X.; Usman, B.; Wang, X.; Luo, L.; Zhao, N.; Liu, Y.; Li, R. Generation of semi-dwarf rice (Oryza sativa L.) lines by CRISPR/Cas9-directed mutagenesis of OsGA20ox2 and proteomic analysis of unveiled changes caused by mutations. 3 Biotech 2019, 9, 387. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.Y.; Qin, X.M.; Luo, L.; Han, Y.; Wang, X.; Usman, B.; Nawaz, G.; Zhao, N.; Liu, Y.G.; Li, R.B. CRISPR/Cas9-Induced Mutagenesis of Semi-Rolled Leaf1,2 Confers Curled Leaf Phenotype and Drought Tolerance by Influencing Protein Expression Patterns and ROS Scavenging in Rice (Oryza sativa L.). Agronomy-Basel 2019, 9, 728. [Google Scholar] [CrossRef]
- Tozawa, Y.; Nozawa, A.; Kanno, T.; Narisawa, T.; Masuda, S.; Kasai, K.; Nanamiya, H. Calcium-activated (p) ppGpp synthetase in chloroplasts of land plants. J. Biol. Chem. 2007, 282, 35536–35545. [Google Scholar] [CrossRef] [PubMed]
- Toivola, J.; Nikkanen, L.; Dahlström, K.M.; Salminen, T.A.; Lepistö, A.; Vignols, F.; Rintamäki, E. Overexpression of chloroplast NADPH-dependent thioredoxin reductase in Arabidopsis enhances leaf growth and elucidates in vivo function of reductase and thioredoxin domains. Front Plant Sci 2013, 4, 389. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.-Y.; Barman, D.N.; Kim, M.G.; Kim, W.-Y. Stress defense mechanisms of NADPH-dependent thioredoxin reductases (NTRs) in plants. Plant signaling & behavior 2015, 10, e1017698. [Google Scholar] [CrossRef]
- Torras-Claveria, L.; Jáuregui, O.; Codina, C.; Tiburcio, A.F.; Bastida, J.; Viladomat, F. Analysis of phenolic compounds by high-performance liquid chromatography coupled to electrospray ionization tandem mass spectrometry in senescent and water-stressed tobacco. Plant Sci. 2012, 182, 71–78. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Foyer, C.H. The roles of reactive oxygen metabolism in drought: Not so cut and dried. Plant Physiol. 2014, 164, 1636–1648. [Google Scholar] [CrossRef]
- Che, Y.; Fu, A.; Hou, X.; McDonald, K.; Buchanan, B.B.; Huang, W.; Luan, S. C-terminal processing of reaction center protein D1 is essential for the function and assembly of photosystem II in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 16247–16252. [Google Scholar] [CrossRef]
- Nakazaki, T.; Tsukiyama, T.; Okumoto, Y.; Kageyama, D.; Naito, K.; Inouye, K.; Tanisaka, T. Distribution, structure, organ-specific expression, and phylogenic analysis of the pathogenesis-related protein-3 chitinase gene family in rice (Oryza sativa L.). Genome 2006, 49, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Arisz, S.A.; Testerink, C.; Munnik, T. Plant PA signaling via diacylglycerol kinase. Biochimica et Biophysica Acta (BBA)-Molecular and Cell Biology of Lipids 2009, 1791, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Laxalt, A.M.; Munnik, T. Phospholipid signalling in plant defence. Curr. Opin. Plant Biol. 2002, 5, 332–338. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, J.; Zhang, H.; Song, F. Overexpression of a rice diacylglycerol kinase gene OsBIDK1 enhances disease resistance in transgenic tobacco. Molecules & Cells (Springer Science & Business Media BV) 2008, 26, 258–264. [Google Scholar]
- Almagro, L.; Gómez Ros, L.; Belchi-Navarro, S.; Bru, R.; Ros Barceló, A.; Pedreno, M. Class III peroxidases in plant defence reactions. J. Exp. Bot. 2008, 60, 377–390. [Google Scholar] [CrossRef]
- Zhang, J.; Yuan, H.; Yang, Y.; Fish, T.; Lyi, S.M.; Thannhauser, T.W.; Zhang, L.; Li, L. Plastid ribosomal protein S5 is involved in photosynthesis, plant development, and cold stress tolerance in Arabidopsis. J. Exp. Bot. 2016, 67, 2731–2744. [Google Scholar] [CrossRef]
- Woloshen, V.; Huang, S.; Li, X. RNA-binding proteins in plant immunity. J. Pathog. 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Kang, H. Emerging roles of RNA-binding proteins in plant growth, development, and stress responses. Mol. Cells 2016, 39, 179. [Google Scholar] [CrossRef]
- Jardim-Messeder, D.; Caverzan, A.; Rauber, R.; de Souza Ferreira, E.; Margis-Pinheiro, M.; Galina, A. Succinate dehydrogenase (mitochondrial complex II) is a source of reactive oxygen species in plants and regulates development and stress responses. New Phytol. 2015, 208, 776–789. [Google Scholar] [CrossRef]
- Belt, K.; Huang, S.; Thatcher, L.F.; Casarotto, H.; Singh, K.B.; Van Aken, O.; Millar, A.H. Salicylic Acid-Dependent Plant Stress Signaling via Mitochondrial Succinate Dehydrogenase. Plant Physiol. 2017, 173, 2029–2040. [Google Scholar] [CrossRef]
- Herrera-Vásquez, A.; Salinas, P.; Holuigue, L. Salicylic acid and reactive oxygen species interplay in the transcriptional control of defense genes expression. Front. Plant Sci. 2015, 6, 171. [Google Scholar] [CrossRef]
- Halim, V.; Vess, A.; Scheel, D.; Rosahl, S. The role of salicylic acid and jasmonic acid in pathogen defence. Plant Biol. 2006, 8, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, J.; Li, Y.; Yuan, Z.; He, H.; Yang, H.; Qu, H.; Ma, C.; Qu, S. Transcriptome analysis highlights defense and signaling pathways mediated by rice pi21 gene with partial resistance to Magnaporthe oryzae. Front. Plant Sci. 2016, 7, 1834. [Google Scholar] [CrossRef] [PubMed]
- Wamaitha, M.J.; Yamamoto, R.; Wong, H.L.; Kawasaki, T.; Kawano, Y.; Shimamoto, K. OsRap2. 6 transcription factor contributes to rice innate immunity through its interaction with Receptor for ctivated Kinase-C 1 (RACK1). Rice 2012, 5, 35. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutation Type | |||||||
|---|---|---|---|---|---|---|---|
| Targets | Bi-Allelic | Homozygous | Heterozygous | Chimeric | WT | Total | |
| Pi21T1 | NOP | 7 | 6 | 2 | 2 | 8 | 25 |
| MR | 28.00 | 24.00 | 8.00 | 8.00 | 68.00 | ||
| Pi21T2 | NOP | 6 | 5 | 4 | 1 | 9 | 25 |
| MR | 24.00 | 20.00 | 16.00 | 4.00 | 64.00 | ||
| Mean MR | 66.00% | ||||||
| Generation | Line | PH (cm) | FLL (cm) | FLW (cm) | PN | PL (cm) | GNPP | SSR (%) | GWT (g) |
|---|---|---|---|---|---|---|---|---|---|
| WT | 105.8 ± 4.3 | 37.36 ± 1.4 | 1.63 ± 0.2 | 10.81 ± 0.7 | 21.36 ± 1.8 | 155 ± 12 | 82.12 ± 5.5 | 29.34 ± 1.1 | |
| GN-3 | 102.5 ± 2.3 ns | 36.55 ± 1.6 ns | 1.51 ± 0.3 ns | 10.44 ± 0.4 ns | 21.84 ± 1.9 ns | 158 ± 10 ns | 83.10 ± 4.1 ns | 28.63 ± 1.6 ns | |
| T1 | GN-5 | 106.6 ± 3.4 ns | 35.23 ± 1.5 ns | 1.67 ± 0.2 ns | 9.98 ± 0.6 ns | 20.70 ± 2.2 ns | 159 ± 11 ns | 85.33 ± 4.5 ns | 29.93 ± 1.1 ns |
| GN-9 | 107.4 ± 3.8 ns | 35.88 ± 1.3 ns | 1.64 ± 0.3 ns | 10.12 ± 0.5 ns | 21.30 ± 1.7 ns | 157 ± 12 ns | 82.03 ± 3.7 ns | 28.43 ± 1.5 ns | |
| GN-13 | 108.1 ± 3.6 ns | 36.28 ± 1.6 ns | 1.59 ± 0.4 ns | 10.29 ± 0.3 ns | 21.88 ± 1.5 ns | 156 ± 13 ns | 83.44 ± 4.3 ns | 28.90 ± 1.2 ns | |
| WT | 103.5 ± 3.9 | 38.16 ± 1.4 | 1.70 ± 0.3 | 9.94 ± 0.9 | 20.75 ± 1.6 | 157 ± 11 | 84.52 ± 3.9 | 28.75 ± 1.2 | |
| GN-3 | 105.2 ± 2.3 ns | 37.21 ± 1.3 ns | 1.62 ± 0.1 ns | 9.70 ± 0.4 ns | 20.99 ± 1.7 ns | 159 ± 10 ns | 84.30 ± 4.1 ns | 29.21 ± 1.5 ns | |
| T2 | GN-5 | 104.8 ± 3.6 ns | 34.29 ± 1.9 ns | 1.56 ± 0.2 ns | 9.50 ± 0.6 ns | 20.26 ± 1.9 ns | 158 ± 12 ns | 85.80 ± 3.6 ns | 28.27 ± 1.2 ns |
| GN-9 | 108.6 ± 3.8 ns | 35.12 ± 1.3 ns | 1.59 ± 0.3 ns | 9.75 ± 0.5 ns | 21.60 ± 2.1 ns | 156 ± 13 ns | 83.83 ± 4.7 ns | 29.34 ± 1.3 ns | |
| GN-13 | 103.3 ± 2.7 ns | 35.50 ± 1.6 ns | 1.54 ± 0.1 ns | 10.50 ± 0.7 ns | 21.40 ± 2.8 ns | 158 ± 12 ns | 82.85 ± 4.9 ns | 28.72 ± 1.4 ns | |
| WT | 107.5 ± 4.1 | 36.46 ± 1.3 | 1.58 ± 0.2 | 10.21 ± 0.8 | 21.15 ± 1.5 | 156 ± 13 | 83.12 ± 3.8 | 29.60 ± 1.4 | |
| GN-3 | 101.4 ± 2.5 ns | 36.12 ± 1.6 ns | 1.69 ± 0.3 ns | 10.17 ± 0.3 ns | 20.41 ± 1.4 ns | 158 ± 11 ns | 83.65 ± 3.3 ns | 28.93 ± 1.3 ns | |
| T3 | GN-5 | 105.9 ± 3.7 ns | 35.91 ± 1.5 ns | 1.62 ± 0.3 ns | 9.72 ± 0.5 ns | 20.55 ± 2.1 ns | 157 ± 12 ns | 85.00 ± 4.4 ns | 29.95 ± 1.2 ns |
| GN-9 | 104.6 ± 3.2 ns | 36.10 ± 1.4 ns | 1.67 ± 0.3 ns | 9.60 ± 0.6 ns | 20.95 ± 2.3 ns | 155 ± 14 ns | 85.52 ± 3.5 ns | 28.75 ± 1.5 ns | |
| GN-13 | 106.7 ± 2.8 ns | 35.85 ± 1.7 ns | 1.60 ± 0.2 ns | 10.21 ± 0.7 ns | 21.75 ± 2.4 ns | 157 ± 12 ns | 84.28 ± 5.3 ns | 29.29 ± 1.2 ns |
| Protein ID | Gene | Annotation | Regulation |
|---|---|---|---|
| Q6Z9C3 | Os08g0547100 | Probable 6-phosphogluconolactonase 3, chloroplastic | Up |
| Q5KQI6 | SNAT1 | Serotonin N-acetyltransferase 1, chloroplastic | Up |
| Q69PS6 | OsNTRA | Thioredoxin reductase NTRA | Up |
| Q0IZQ2 | Os09g0556500 | Cysteine—tRNA ligase CPS1 homolog, chloroplastic | Up |
| Q6ATB2 | CRSH2 | Probable GTP diphosphokinase CRSH2, chloroplastic | Up |
| Q7XTK3 | AMI1 | Amidase 1 | Up |
| Q6K8R2 | Cht6 | Pathogenesis related (PR)-3 chitinase 6 | Up |
| Q67WM9 | OS9 | Protein OS-9 homolog | Up |
| Q75T45 | RSOsPR10 | Os12g0555000 protein | Up |
| Q10HD0 | RCABP89 | Chlorophyll a-b binding protein, chloroplastic | Up |
| Q6ESR4 | DHN1 | Dehydration stress-inducible protein 1 | Up |
| Q6K4P5 | Os02g0787800 | diacylglycerol kinase | Up |
| Q84ZE8 | OsJ_24279 | Auxin-regulated protein-like | Up |
| A0A0P0XMI4 | Os09g0446200 | γ-tubulin complex component | Up |
| Q2R351 | LOC_Os11g33120 | Respiratory burst oxidase protein D | Up |
| A3BUY8 | OsJ_27981 | PEROXIDASE_4 domain-containing protein | Up |
| A0A0P0VAH1 | Os01g0849000 | Os01g0849000 protein | Up |
| Q75IS1 | Os05g0162000 | Peroxidase | Up |
| Q5U1N1 | prx62 | Peroxidase | Up |
| Q8HCQ0 | nad3 | NADH-ubiquinone oxidoreductase chain 3 | Up |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nawaz, G.; Usman, B.; Peng, H.; Zhao, N.; Yuan, R.; Liu, Y.; Li, R. Knockout of Pi21 by CRISPR/Cas9 and iTRAQ-Based Proteomic Analysis of Mutants Revealed New Insights into M. oryzae Resistance in Elite Rice Line. Genes 2020, 11, 735. https://doi.org/10.3390/genes11070735
Nawaz G, Usman B, Peng H, Zhao N, Yuan R, Liu Y, Li R. Knockout of Pi21 by CRISPR/Cas9 and iTRAQ-Based Proteomic Analysis of Mutants Revealed New Insights into M. oryzae Resistance in Elite Rice Line. Genes. 2020; 11(7):735. https://doi.org/10.3390/genes11070735
Chicago/Turabian StyleNawaz, Gul, Babar Usman, Haowen Peng, Neng Zhao, Ruizhi Yuan, Yaoguang Liu, and Rongbai Li. 2020. "Knockout of Pi21 by CRISPR/Cas9 and iTRAQ-Based Proteomic Analysis of Mutants Revealed New Insights into M. oryzae Resistance in Elite Rice Line" Genes 11, no. 7: 735. https://doi.org/10.3390/genes11070735
APA StyleNawaz, G., Usman, B., Peng, H., Zhao, N., Yuan, R., Liu, Y., & Li, R. (2020). Knockout of Pi21 by CRISPR/Cas9 and iTRAQ-Based Proteomic Analysis of Mutants Revealed New Insights into M. oryzae Resistance in Elite Rice Line. Genes, 11(7), 735. https://doi.org/10.3390/genes11070735