1. Introduction
In some fish taxa, polyploidization is an ongoing process and can occur, even within a species (autopolyploidy; i.e., polyploidy derived from a single species), while in other taxa, e.g., Cyprinidae, it is rare [
1]. Polyploidy is relatively frequent in interspecific hybrids of fishes (allopolyploidy; i.e., polyploidy arising from interspecific hybridization). Berrebi [
2] and Birstein et al. [
3] pointed out that polyploidy increased the success of interspecific hybridization in fishes.
Sturgeons represent ancestral ray-finned fishes (besides bichirs and ropefishes) in the class Actinopterygii [
4], originating during the Jurassic era 200 million years ago [
5,
6]. The order Acipenseriformes consists of 27 living species, many of which are listed in the Appendices to CITES (Convention on International Trade in Endangered Species of Wild Fauna and Flora; [
7]). They diverged before the teleost specific genome duplication from the other ray-finned fishes [
8]. The sturgeons split into three families, one extinct and two presently living—Acipenseridae and Polyodontidae [
9]. In the family Polyodontidae, no further genome duplication has occurred, and they are considered “functional diploids” (2n) with around 120 chromosomes. There are only two species in the family: American paddlefish (
P. spathula), Chinese paddlefish (
Psephurus gladius). The extinction of the Chinese paddlefish has been recently reported [
10]. Several polyploidization evens occurred later in Acipenseridae, which can contribute to the resilience and adaptability of species belonging to Acipenseridae. Due to these evolutionary processes, as a byproduct, hybridization can be successful between Acipenseridae, and even among taxa with different chromosome numbers. In Acipenseridae, at least one additional genome duplication happened during the evolution of the Atlantic clade of the family, and more during the shortnose sturgeon (
Acipenser brevirostrum) and the pacific clade evolution. Most of the species in the Atlantic clade, including the Russian sturgeon (
A. gueldenstaedtii), are considered “functional tetraploid“ (4n) with ≈250 chromosomes [
11].
Although hybridization among acipenserid species is common, there are no reports of successful hybridization of acipenserids and polyodontids. Previous hybridization experiments on shovelnose sturgeon (
Scaphirhynchus platorynchus) × American paddlefish [
12,
13] or American paddlefish × Amur sturgeon (
Acipenser schrenckii) [
14] have failed to result in viable offspring. Besides the large phylogenetic distance (i.e., they diverged 184.4 Mya [
15]), representatives of Polyodontidae and Acipenseridae differ in their gross morphology (e.g., presence of scutes, the structure of mouth, rostrum, filter apparatus) as well as feeding behavior, preferred habitat, etc. [
3,
16]. This suggests an inability to hybridize. In hybrids of acipenserids, odd ploidy plasticity may occur, which is highly dependent on the ploidy level of the parental species. Various ploidy levels are caused either by polyspermy or partial or full duplication of the maternal genome and hybridization [
7,
17]. Mosaicism also may occur [
18,
19,
20].
During an experiment to produce gynogenic Russian sturgeon progeny, a negative control was initiated using non-irradiated American paddlefish sperm and eggs from the Russian sturgeon. Unexpectedly, the control cross resulted in viable hybrids. This was the first report of viable hybrids between any acipenserid and Polyodon species. In this study, we described the morphology, ploidy level, and chromosome number, demonstrating the hybrid origin of the progeny. This study addressed whether the simultaneous genome duplication occurs and how it affects the ploidy level of hybrid progenies. Finally, it described the relationship between the external morphology and ploidy level in the hybrids of species with such great phylogenetic distance.
4. Discussion
Former experiences on the hybridization of acipenserids and paddlefishes [
14,
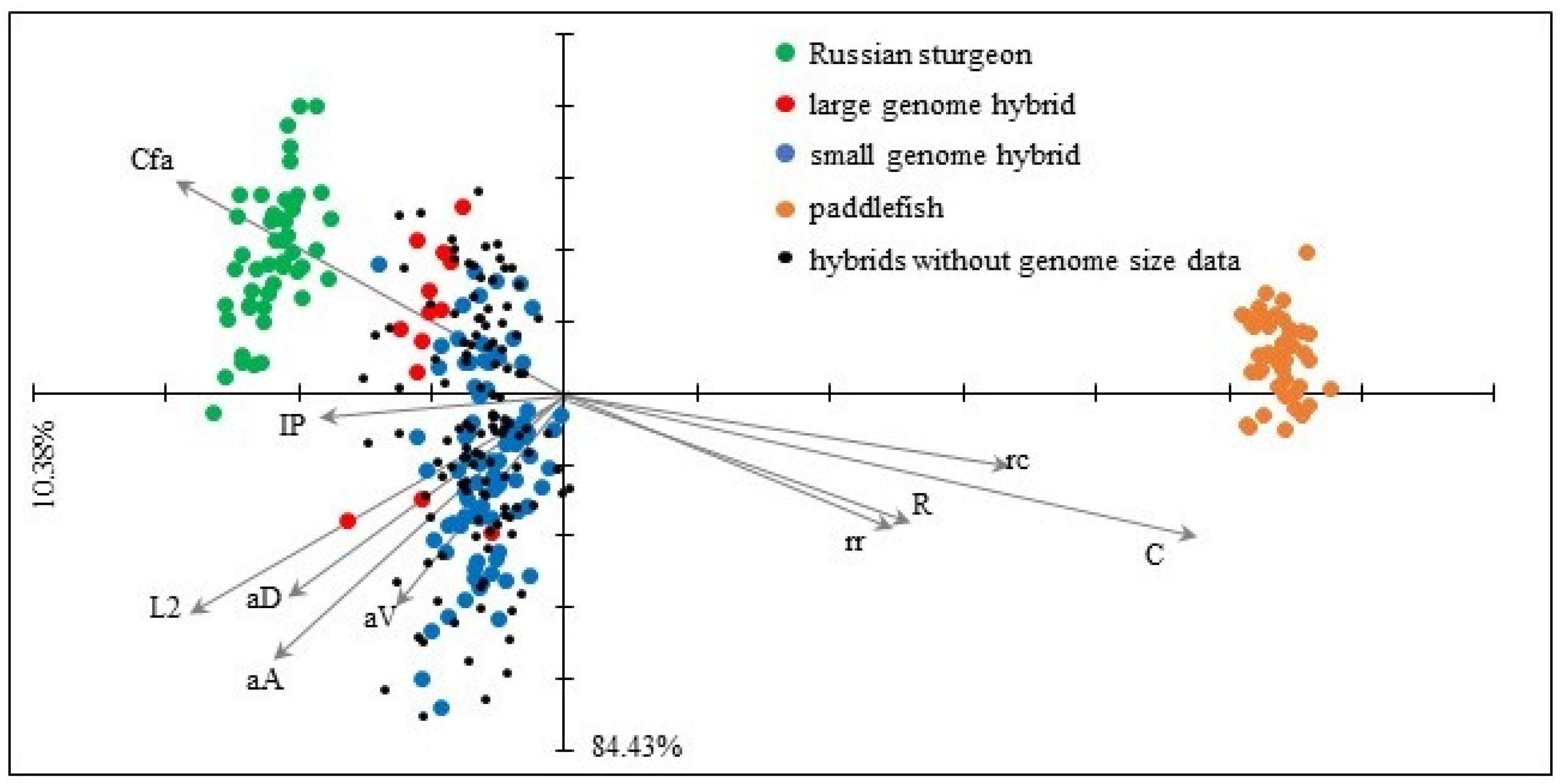
38] support the assumed inability of the hybridization of taxa with such a high phylogenetic distance separating them. Still, the cross of female Russian sturgeon and male American paddlefish, as a negative control for gynogenesis, resulted in viable hybrids. The fertilization, hatching, and survival rates of these hybrids were close to that of the pure maternal species. The progenies exhibited two ploidy levels (triploid and pentaploid) due to spontaneous polyploidization, relating to external morphological and meristic characteristics.
The reproductive success of these two distant species could be the combined consequence of the ancient genome duplication (180 Mya, that occurred in the common ancestor of the two families) and the slow evolution rate of acipenserids. The theory of slow evolution was reinforced by the “small” morphological differences of the living sturgeon species, the smaller substitution rates of mitochondrial or nuclear genes, and the slower protein evolution, compared to other fish species. This theory has been recently confirmed by a detailed evaluation of the sterlet genome [
39]. Since the common sturgeon genome duplication, the sterlet genome has not formed back to a diploid state, and structural and functional elements have been found on the tetraploid level in unexpectedly high degree [
39]. These phenomena could lead to a higher similarity, compatibility, and flexibility among the sturgeon genomes and allow the hybridization between Russian sturgeon and American paddlefish despite the large geographical, physiological, and morphological distances.
The intermediate appearance is common in interspecific hybrids [
40,
41]. A further increase of morphological variability occurs in polyploid hybrids, primarily due to the difference in gene dosage from parent species [
42]. Hybrids showed the widest variability in characters, which differed the most parental species, i.e., rostrum-length-related characters and length of the upper caudal lobe. The LH individuals were more similar to the maternal species, presumably due to the larger gene dosage from Russian sturgeon overwhelming the paddlefish-like traits [
43]. The SH hybrids showed a more diverse appearance since the ration of maternal and paternal chromosome sets is more balanced than LH hybrids [
43].
Similar to the interspecific teleost hybrids, where a substantial difference has been observed in size and the number of scales between hybrids and parental species [
40,
43], the number of scutes differed substantially in hybrids in conjunction with ploidy level. The presence or absence of the scutes, as well as their modified forms in the hybrids, offer the opportunity to study the role of the fibroblast growth factor receptor gene (fgfr) as it influences the number and size of scales in the mirror, nude, and scaled forms of common carp and zebrafish [
44]. Sub-functionalization of paralogues of this gene may have occurred following genome duplication during the evolutionary process [
45].
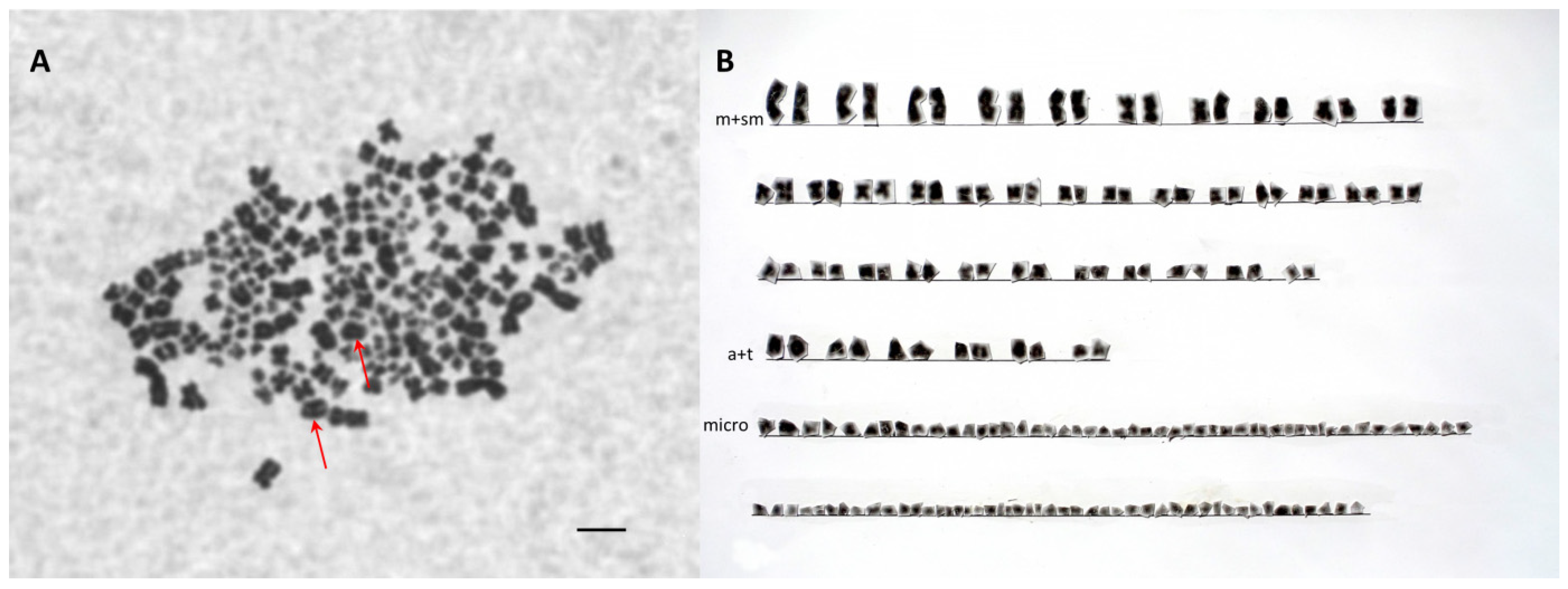
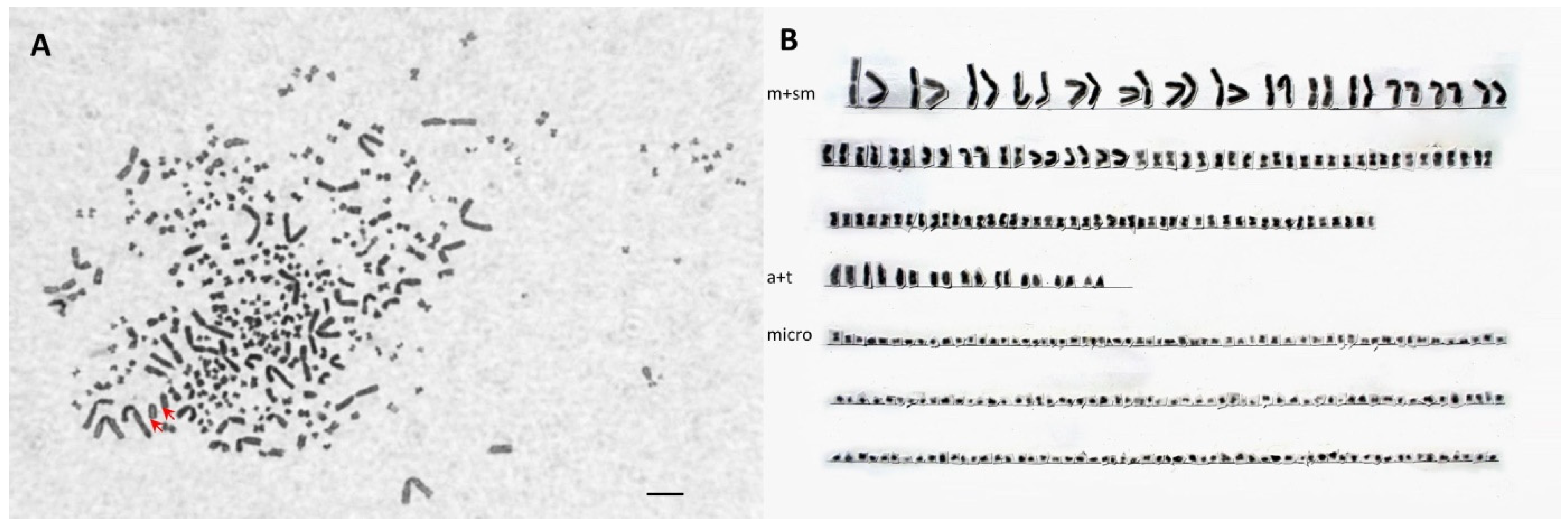
The characteristics of karyotypes and the results of microsatellite analysis confirmed that the progenies were interspecies hybrids. As with Váradi et al. [
46], chromosome analysis revealed the occurrence of two ploidy levels in hybrids: triploid hybrids (SH, AP) and pentaploid hybrids (LH, AAP). Slight variation in chromosome numbers was also observed within the groups. On the one hand, inconsistency in the published chromosome numbers within a pure-bred species could be the consequence of difficulties in the determination of microchromosome numbers [
37]. On the other hand, microchromosome loss also occurs in interspecies hybrids of species characterized by different ploidy levels [
47]. We observed haploid paternal cell lineages in some individuals in both groups, suggesting polyspermy [
18]. In this case, a supernumerary spermatozoon began to develop as a single cell line, and triploid/pentaploid and haploid cell lines were present in the genome of individuals, causing mosaicism in the hybrids.
Investigations on the functional scale of the ploidy level [
20] assume that the interspecific differences in ploidy levels (e.g., diploid American paddlefish and tetraploid Russian sturgeon) originated from whole-genome duplication events during the evolution of Acipenseriformes [
8]. Furthermore, intraspecific polyploidization is also relatively frequent among the acipenseriform fishes. It has been detected in white sturgeon (
Acipenser transmontanus) [
48], Siberian sturgeon (
Acipenser baerii) [
20,
49], kaluga (
Huso dauricus) [
14], sterlet [
50], and Sakhalin sturgeon (
Acipenser mikadoi) [
14]. The spontaneous intraspecific polyploidization can result in variance in genome size. Havelka et al. [
20] described the spontaneous duplication of the maternal chromosome set in Siberian sturgeon of different ploidy caused by either premeiotic endomitosis or the retention of the second polar body. Many other studies [
51,
52,
53] have stated the retention that the second polar body can lead to polyploidization. The different ratios of triploid/pentaploid individuals among the parent combinations in our study also supported this hypothesis.
Microsatellites from both species and hybrids were amplified successfully. All the used microsatellite markers were suitable for distinguishing the two species and for verification of the hybridization. Previously, Zou et al. [
14] used the Psp-28, Psp-29, Psp-32 markers on Polyodontidae and Acipenseridae fishes to verify the success of gynogenesis. They also found different allele ranges between the paddlefish and Amur sturgeon. However, the allele sizes could not be compared directly because different techniques were used for allele sizing (polyacrylamide gels and silver staining). All three Psp loci were disomic in that experiment, while the Psp-29 appeared to be tetrasomic in our experiment as well as in the description of Heist et al. [
24]. The analyses proved the presence of both the Russian sturgeon and American paddlefish genomes in all examined hybrid individuals, discarding the possibility of spontaneous gynogenesis, which was observed in Siberian sturgeons [
20,
49] and sterlet [
54]. The two tetrasomic markers enabled us to follow the segregation of the tetraploid Russian sturgeon genome and to analyze ploidy levels. These markers showed the presence of only one American paddlefish and two Russian sturgeon alleles in the SH genomes, while in the LH genomes, more alleles were found. This suggested the presence of spontaneous duplication of the maternal genomes in the individuals with high chromosome numbers (250 ± 8–304 ± 4) and/or nucleus size.
The polyploid status of the species within the Acipenserdae family, as well as their similar karyotype and genome, contribute to easy interspecific and intergeneric hybridization [
3]. Based on our findings, successful interspecific hybridization is also supposed in other species combinations in the order Acipenseriformes. Consequently, American paddlefish may hybridize with other sturgeons independent of ploidy level.
The aquaculture potential of this hybrid is a subject for debate. Sturgeon hybrids are commonly used in aquaculture to exploit the advantages of heterosis [
55]. It is estimated that hybrids account for approximately 35 and 20% percent of global sturgeon meat and caviar production, respectively [
56]. If the planktivorous feeding habit of parent species of paddlefish is inherited to a certain extent, the new hybrid can play an important role in adapting pond aquaculture to the challenges of climate change. Non-fed species have a lower carbon footprint than fed ones, and the co-culture of a filter feeder with relatively high market value as a supplementary species would strengthen pond aquaculture both from an ecological and environmental perspective [
57].

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}