Allergic Inflammation Alters microRNA Expression Profile in Adipose Tissue in the Rat

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
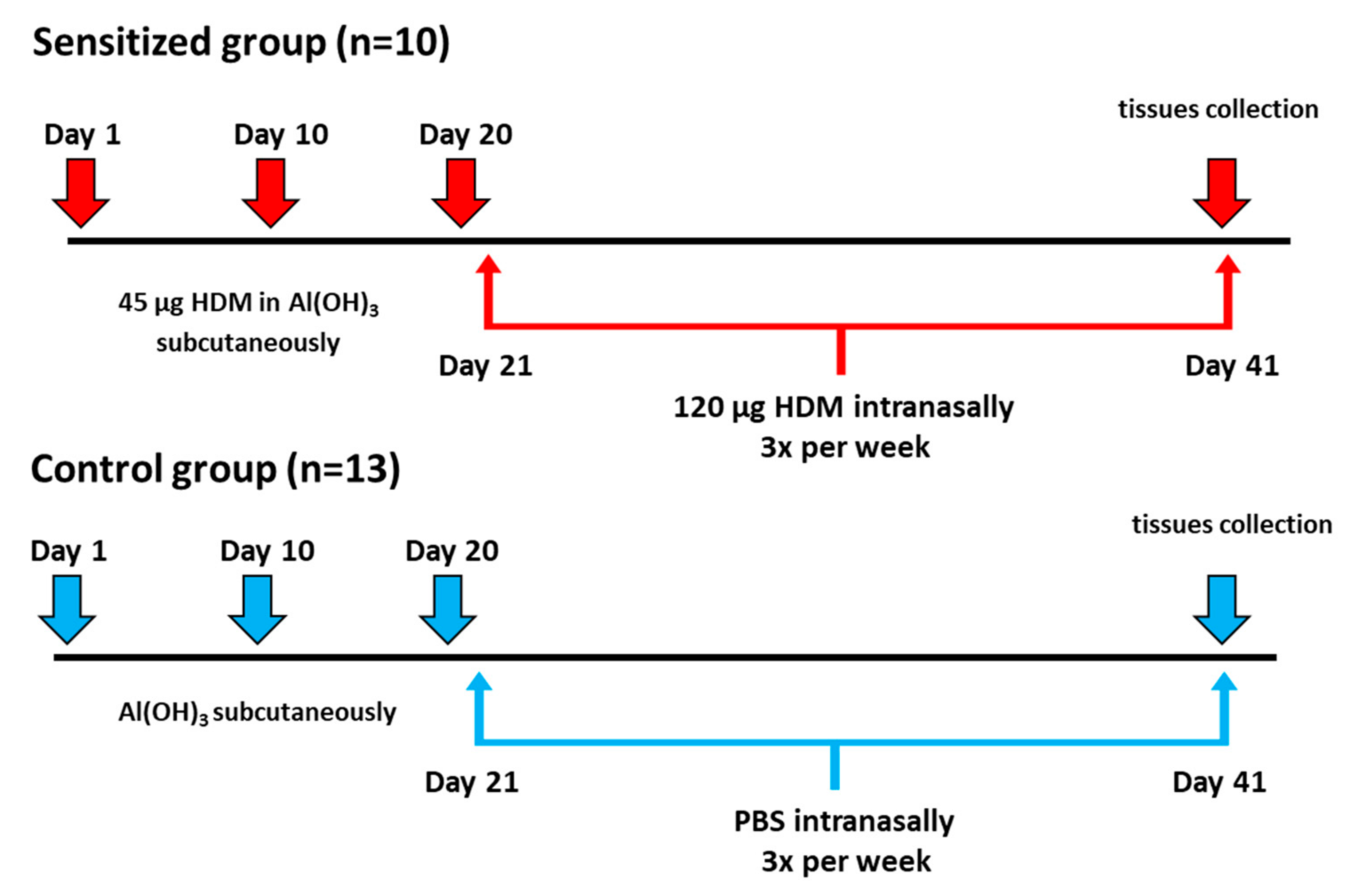
2.1. Animal Model of Allergic Inflammation
2.2. Histopathological Analysis
2.3. Eosinophil Count
2.4. Glucose Measurement
2.5. ELISA Method
2.6. Small RNA Sequencing
2.7. MiRNA Expression Analysis
2.8. Gene Expression Analysis
2.9. MiRNA Pathway Analysis
2.10. Statistical Analysis
3. Results
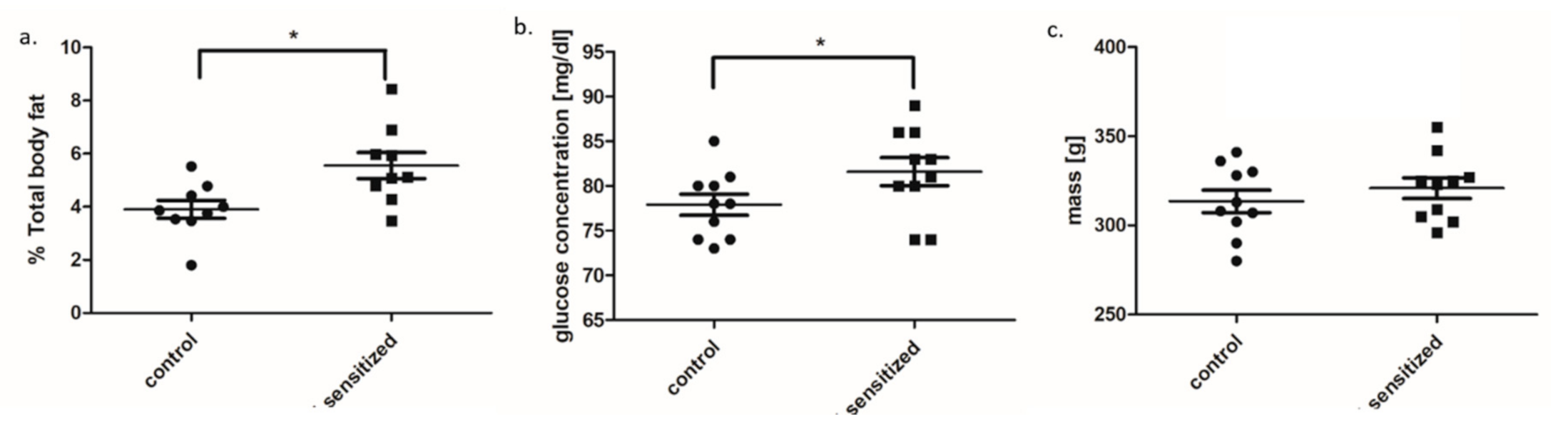
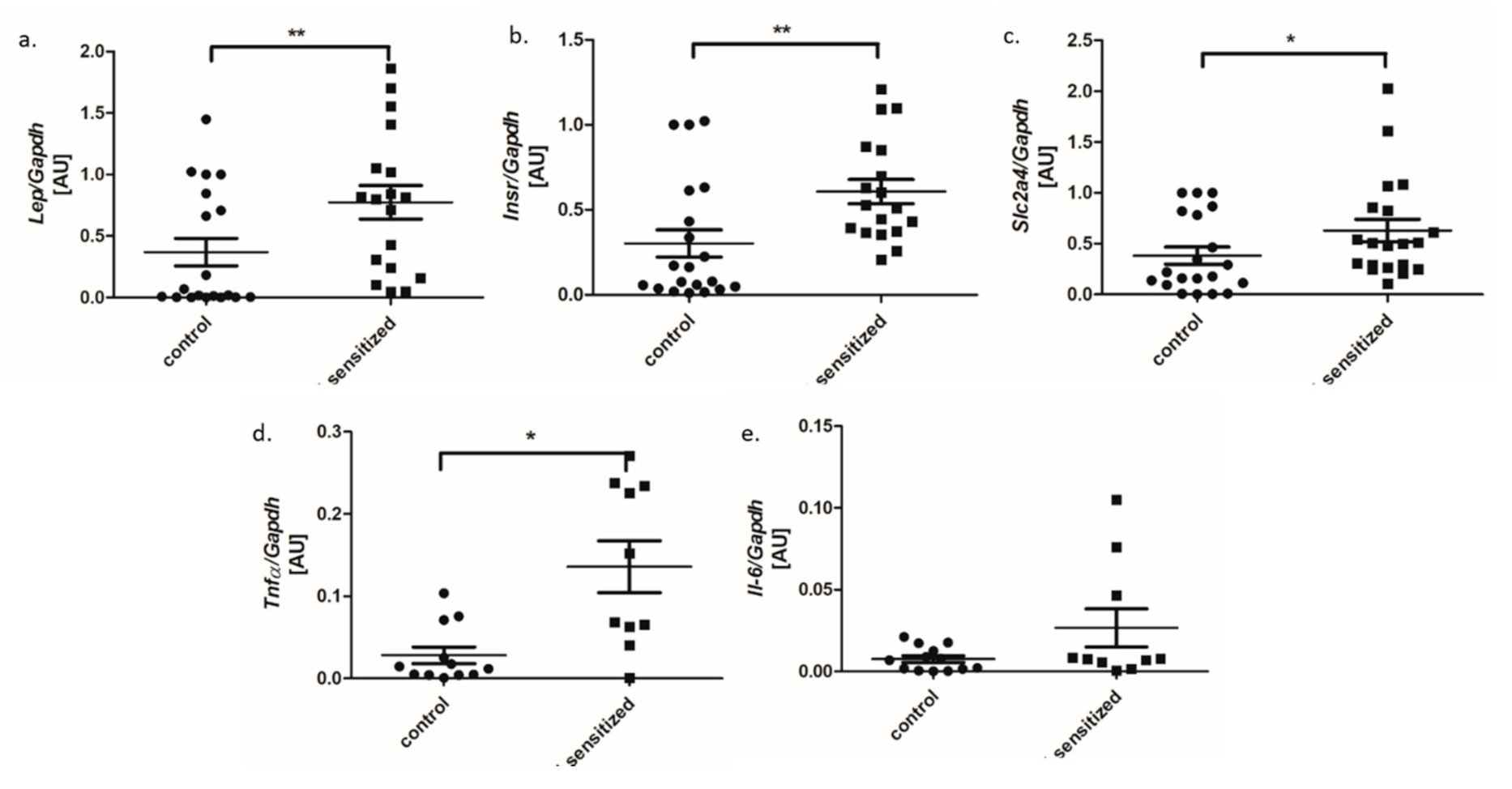
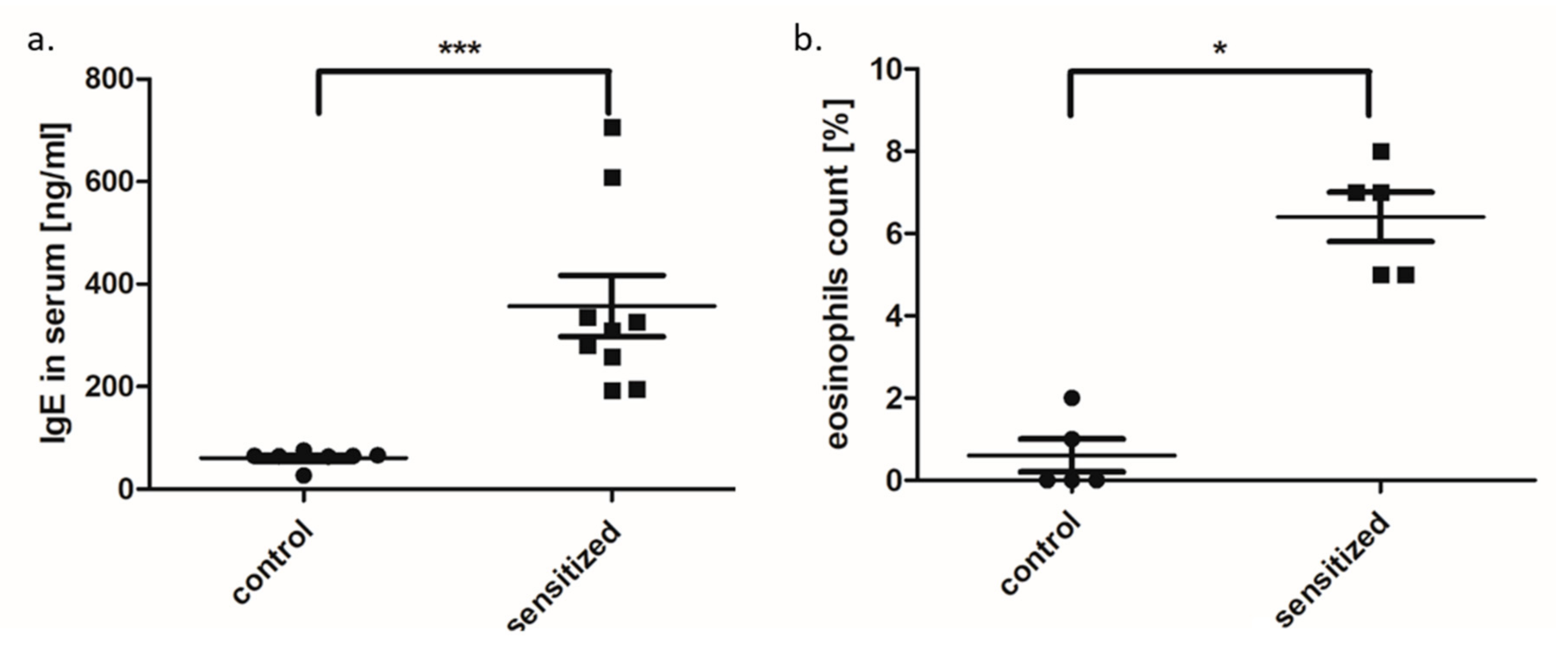
3.1. Metabolic and Inflammatory Parameters in An Allergy Model
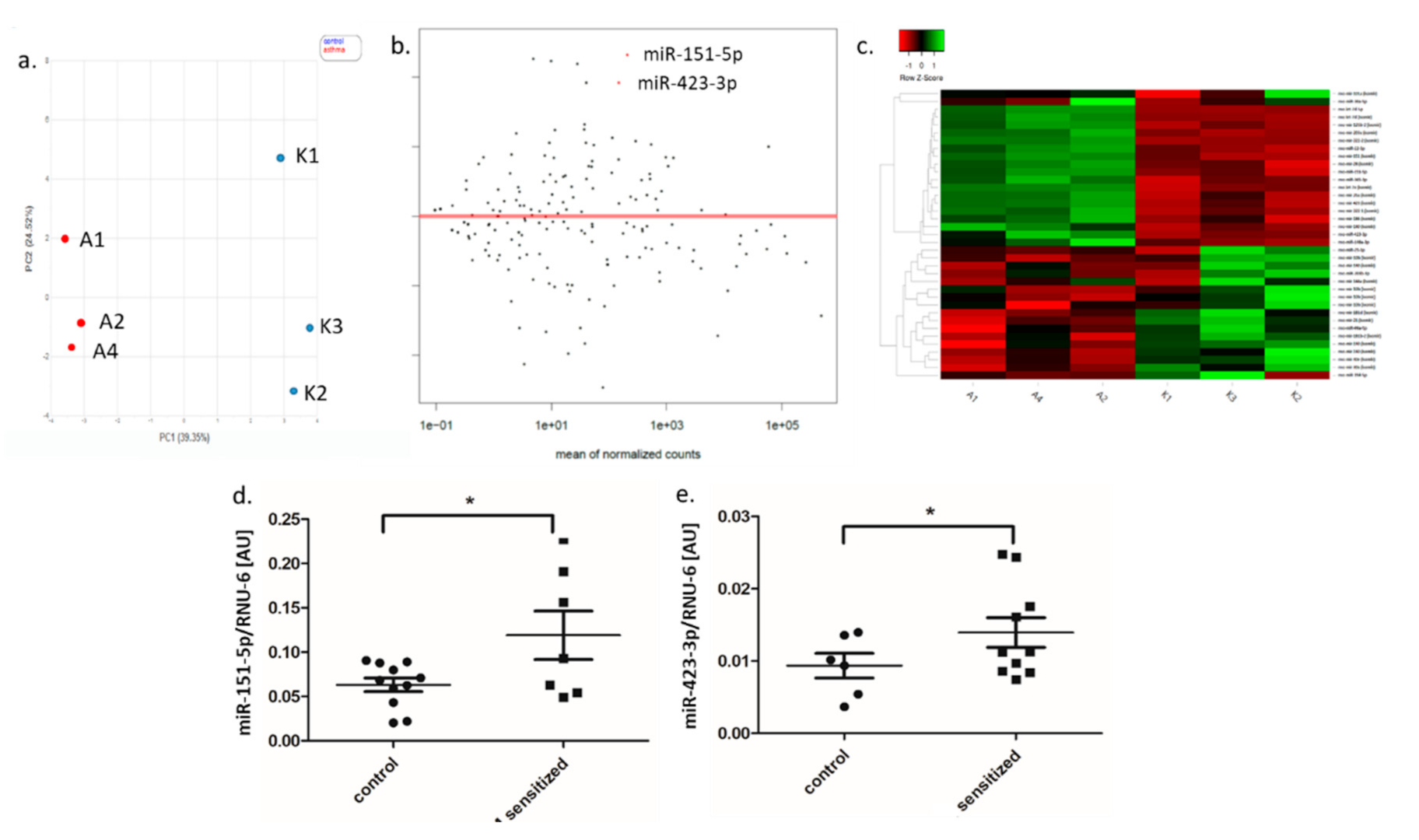
3.2. Small RNA Expression Profile in Adipose Tissue
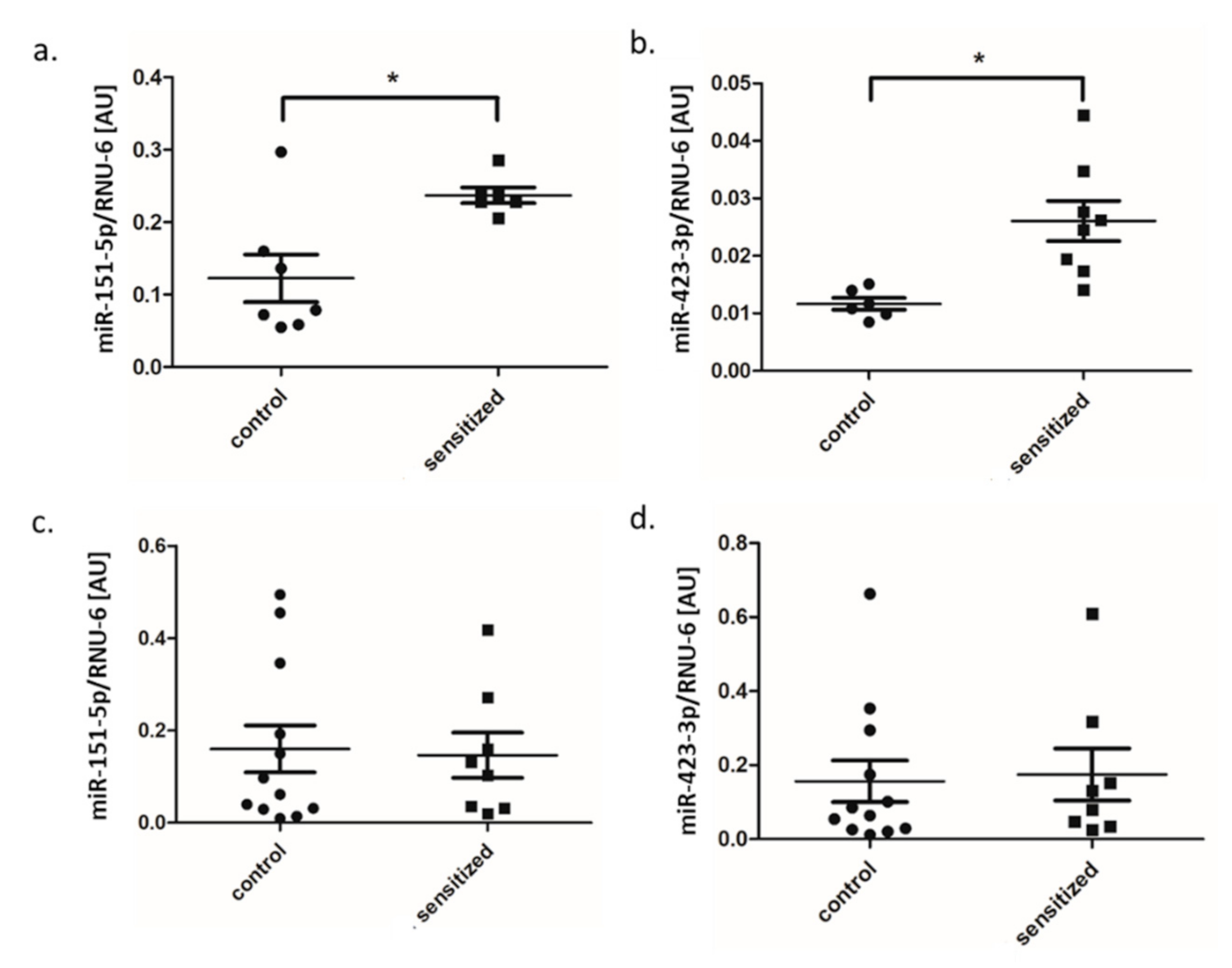
3.3. MiRNA Expression in Lung and BALF
3.4. MiRNA Pathway Prediction
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thomou, T.; Mori, M.A.; Dreyfuss, J.M.; Konishi, M.; Sakaguchi, M.; Wolfrum, C.; Rao, T.N.; Winnay, J.N.; Garcia-Martin, R.; Grinspoon, S.K.; et al. Adipose-derived circulating miRNAs regulate gene expression in other tissues. Nature 2017, 542, 450–455. [Google Scholar] [CrossRef]
- Arner, E.; Mejhert, N.; Kulyte, A.; Balwierz, P.J.; Pachkov, M.; Cormont, M.; Lorente-Cebrian, S.; Ehrlund, A.; Laurencikiene, J.; Heden, P.; et al. Adipose tissue microRNAs as regulators of CCL2 production in human obesity. Diabetes 2012, 61, 1986–1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumortier, O.; Hinault, C.; Van Obberghen, E. MicroRNAs and metabolism crosstalk in energy homeostasis. Cell Metab. 2013, 18, 312–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trajkovski, M.; Hausser, J.; Soutschek, J.; Bhat, B.; Akin, A.; Zavolan, M.; Heim, M.H.; Stoffel, M. MicroRNAs 103 and 107 regulate insulin sensitivity. Nature 2011, 474, 649–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, W.; Riopel, M.; Bandyopadhyay, G.; Dong, Y.; Birmingham, A.; Seo, J.B.; Ofrecio, J.M.; Wollam, J.; Hernandez-Carretero, A.; Fu, W.; et al. Adipose Tissue Macrophage-Derived Exosomal miRNAs Can Modulate In Vivo and In Vitro Insulin Sensitivity. Cell 2017, 171, 372–384. [Google Scholar] [CrossRef]
- Capobianco, V.; Nardelli, C.; Ferrigno, M.; Iaffaldano, L.; Pilone, V.; Forestieri, P.; Zambrano, N.; Sacchetti, L. miRNA and protein expression profiles of visceral adipose tissue reveal miR-141/YWHAG and miR-520e/RAB11A as two potential miRNA/protein target pairs associated with severe obesity. J. Proteom. Res. 2012, 11, 3358–3369. [Google Scholar] [CrossRef]
- Caroli, A.; Cardillo, M.T.; Galea, R.; Biasucci, L.M. Potential therapeutic role of microRNAs in ischemic heart disease. J. Cardiol. 2013, 61, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Scadding, G. Cytokine profiles in allergic rhinitis. Curr. Allergy Asthma Rep. 2014, 14, 435. [Google Scholar] [CrossRef]
- Kips, J.C. Cytokines in asthma. Eur. Respir. J. Suppl. 2001, 34, 24s–33s. [Google Scholar] [CrossRef] [Green Version]
- Ge, Q.; Brichard, S.; Yi, X.; Li, Q. microRNAs as a new mechanism regulating adipose tissue inflammation in obesity and as a novel therapeutic strategy in the metabolic syndrome. J. Immunol. Res. 2014, 2014, 987285. [Google Scholar] [CrossRef] [Green Version]
- Baskara-Yhuellou, I.; Tost, J. The impact of microRNAs on alterations of gene regulatory networks in allergic diseases. Adv. Protein Chem. Struct. Biol. 2020, 120, 237–312. [Google Scholar]
- Specjalski, K.; Niedoszytko, M. MicroRNAs: Future biomarkers and targets of therapy in asthma? Curr. Opin. Pulm. Med. 2020, 26, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, A.; Maurya, M.; Maurya, P.; Barthwal, M.K. Lin28B Regulates Angiotensin II-Mediated Let-7c/miR-99a MicroRNA Formation Consequently Affecting Macrophage Polarization and Allergic Inflammation. Inflammation 2020. [Google Scholar] [CrossRef] [PubMed]
- Shefler, I.; Salamon, P.; Mekori, Y.A. MicroRNA Involvement in Allergic and Non-Allergic Mast Cell Activation. Int. J. Mol. Sci. 2019, 20, 2145. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Lai, Y.; Zheng, J.; Chen, K.; Jiang, H.; Xu, G. MiR-146a Promotes Tolerogenic Properties of Dendritic Cells and through Targeting Notch1 Signaling. Immunol. Investig. 2020, 49, 555–570. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhou, Q.; Shang, Y. MiRNA-451a inhibits airway remodeling by targeting Cadherin 11 in an allergic asthma model of neonatal mice. Int. Immunopharmacol. 2020, 83, 106440. [Google Scholar] [CrossRef]
- Xie, Q.M.; Wu, X.; Wu, H.M.; Deng, Y.M.; Zhang, S.J.; Zhu, J.P.; Dong, X.W. Oral administration of allergen extracts from Dermatophagoides farinae desensitizes specific allergen-induced inflammation and airway hyperresponsiveness in rats. Int. Immunopharmacol. 2008, 8, 1639–1645. [Google Scholar] [CrossRef]
- Szczepankiewicz, D.; Skrzypski, M.; Pruszynska-Oszmalek, E.; Kolodziejski, P.A.; Sassek, M.; Stefanska, B.; Nowak, K.W.; Szczepankiewicz, A. Interleukin 4 affects lipid metabolism and the expression of pro-inflammatory factors in mature rat adipocytes. Immunobiology 2018, 223, 677–683. [Google Scholar] [CrossRef]
- Mito, N.; Kitada, C.; Hosoda, T.; Sato, K. Effect of diet-induced obesity on ovalbumin-specific immune response in a murine asthma model. Metab. Clin. Exp. 2002, 51, 1241–1246. [Google Scholar] [CrossRef]
- Calixto, M.C.; Lintomen, L.; Schenka, A.; Saad, M.J.; Zanesco, A.; Antunes, E. Obesity enhances eosinophilic inflammation in a murine model of allergic asthma. Br. J. Pharmacol. 2010, 159, 617–625. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.S.; Park, M.K.; Kang, S.A.; Park, H.Y.; Hong, S.L.; Park, H.K.; Yu, H.S.; Roh, H.J. Adipose-derived stem cells ameliorate allergic airway inflammation by inducing regulatory T cells in a mouse model of asthma. Med. Inflamm. 2014, 2014, 436476. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, N.; Mandour, Y.M.H.; Farid, A.S.; Nafie, E.; Mohamed, A.Z.; Safwat, M.; Taha, R.; Sabry, D.; Sorour, S.M.; Refae, A. Adipose Tissue-Derived Mesenchymal Stem Cell Modulates the Immune Response of Allergic Rhinitis in a Rat Model. Int. J. Mol. Sci. 2019, 20, 873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sastre, B.; Canas, J.A.; Rodrigo-Munoz, J.M.; Del Pozo, V. Novel Modulators of Asthma and Allergy: Exosomes and MicroRNAs. Front. Immunol. 2017, 8, 826. [Google Scholar] [CrossRef] [PubMed]
- Mannucci, C.; Casciaro, M.; Minciullo, P.L.; Calapai, G.; Navarra, M.; Gangemi, S. Involvement of microRNAs in skin disorders: A literature review. Allergy Asthma Proc. 2017, 38, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Joetham, A.; Schedel, M.; Ning, F.; Wang, M.; Takeda, K.; Gelfand, E.W. Dichotomous role of TGF-β controls inducible regulatory T-cell fate in allergic airway disease through Smad3 and TGF-β-activated kinase 1. J. Allergy Clin. Immunol. 2020, 145, 933–946. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Yang, C.; Yang, W.; Jiang, T. Effect of Dexamethasone on TGF-beta1/Smad3 Signalling Pathway in Airway Remodelling Model of Asthmatic Rats. J. Coll. Physicians Surg. Pak. 2019, 29, 537–540. [Google Scholar] [CrossRef]
- Chung, S.; Lee, T.J.; Reader, B.F.; Kim, J.Y.; Lee, Y.G.; Park, G.Y.; Karpurapu, M.; Ballinger, M.N.; Qian, F.; Rusu, L.; et al. FoxO1 regulates allergic asthmatic inflammation through regulating polarization of the macrophage inflammatory phenotype. Oncotarget 2016, 7, 17532–17546. [Google Scholar] [CrossRef]
- Liu, L.Y.; Wang, H.; Xenakis, J.J.; Spencer, L.A. Notch signaling mediates granulocyte-macrophage colony-stimulating factor priming-induced transendothelial migration of human eosinophils. Allergy 2015, 70, 805–812. [Google Scholar] [CrossRef]
- Pelaia, G.; Renda, T.; Gallelli, L.; Vatrella, A.; Busceti, M.T.; Agati, S.; Caputi, M.; Cazzola, M.; Maselli, R.; Marsico, S.A. Molecular mechanisms underlying airway smooth muscle contraction and proliferation: Implications for asthma. Respir. Med. 2008, 102, 1173–1181. [Google Scholar] [CrossRef]
- Yang, T.R.; Zhang, T.; Mu, N.H.; Ruan, L.B.; Duan, J.L.; Zhang, R.P.; Miao, Y.B. Resina draconis inhibits the endoplasmic-reticulum-induced apoptosis of myocardial cells via regulating miR-423-3p/ERK signaling pathway in a tree shrew myocardial ischemia- reperfusion model. J. Biosci. 2019, 44, 53. [Google Scholar] [CrossRef]
- Wang, R.; Li, G.; Zhuang, G.; Sun, S.; Song, Z. Overexpression of microRNA-423-3p indicates poor prognosis and promotes cell proliferation, migration, and invasion of lung cancer. Diagn. Pathol. 2019, 14, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Li, T.; Chen, G.; Yan, G.; Zhang, X.; Wan, Y.; Li, Q.; Zhu, B.; Zhuo, W. Identification of a serum microRNA expression signature for detection of lung cancer, involving miR-23b, miR-221, miR-148b and miR-423-3p. Lung Cancer 2017, 114, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Olioso, D.; Dauriz, M.; Bacchi, E.; Negri, C.; Santi, L.; Bonora, E.; Moghetti, P. Effects of Aerobic and Resistance Training on Circulating Micro-RNA Expression Profile in Subjects With Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2019, 104, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.; Bae, H.; Song, G. Differential expression of apolipoprotein D in male reproductive system of rats by high-fat diet. Andrology 2016, 4, 1115–1122. [Google Scholar] [CrossRef]
- Chen, C.; Wang, D.; Moshaverinia, A.; Liu, D.; Kou, X.; Yu, W.; Yang, R.; Sun, L.; Shi, S. Mesenchymal stem cell transplantation in tight-skin mice identifies miR-151-5p as a therapeutic target for systemic sclerosis. Cell Res. 2017, 27, 559–577. [Google Scholar] [CrossRef]
- Choi, E.J.; Kim, H.B.; Baek, Y.H.; Kim, E.H.; Pascua, P.N.; Park, S.J.; Kwon, H.I.; Lim, G.J.; Kim, S.; Kim, Y.I.; et al. Differential microRNA expression following infection with a mouse-adapted, highly virulent avian H5N2 virus. BMC Microbiol. 2014, 14, 252. [Google Scholar] [CrossRef] [Green Version]
- Lian, Y.; Xiong, F.; Yang, L.; Bo, H.; Gong, Z.; Wang, Y.; Wei, F.; Tang, Y.; Li, X.; Liao, Q.; et al. Long noncoding RNA AFAP1-AS1 acts as a competing endogenous RNA of miR-423-5p to facilitate nasopharyngeal carcinoma metastasis through regulating the Rho/Rac pathway. J. Exp. Clin. Cancer Res. CR 2018, 37, 253. [Google Scholar] [CrossRef] [Green Version]
- Moller, L.L.V.; Klip, A.; Sylow, L. Rho GTPases-Emerging Regulators of Glucose Homeostasis and Metabolic Health. Cells 2019, 8, 434. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Yan, W.; Wang, S.E.; Baltimore, D. Dual mechanisms of posttranscriptional regulation of Tet2 by Let-7 microRNA in macrophages. Proc. Natl. Acad. Sci. USA 2019, 116, 12416–12421. [Google Scholar] [CrossRef] [Green Version]
- Gubernatorova, E.O.; Gorshkova, E.A.; Namakanova, O.A.; Zvartsev, R.V.; Hidalgo, J.; Drutskaya, M.S.; Tumanov, A.V.; Nedospasov, S.A. Non-redundant Functions of IL-6 Produced by Macrophages and Dendritic Cells in Allergic Airway Inflammation. Front. Immunol. 2018, 9, 2718. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.L.; Chen, S.H.; Wang, J.Y. Critical role of IL-6 in dendritic cell-induced allergic inflammation of asthma. J. Mol. Med. 2016, 94, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Squillace, N.; Bresciani, E.; Torsello, A.; Bandera, A.; Sabbatini, F.; Giovannetti, C.; Giunta, G.; Rovati, L.; Del Bene, M.; Locatelli, V.; et al. Changes in subcutaneous adipose tissue microRNA expression in HIV-infected patients. J. Antimicrob. Chem. 2014, 69, 3067–3075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Chen, Z.; Tan, X.; Zhou, F.; Tan, F.; Gao, Y.; Sun, N.; Xu, X.; Shao, K.; He, J. MicroRNA-99a/100 promotes apoptosis by targeting mTOR in human esophageal squamous cell carcinoma. Med. Oncol. 2013, 30, 411. [Google Scholar] [CrossRef] [PubMed]
- Li, X.J.; Luo, X.Q.; Han, B.W.; Duan, F.T.; Wei, P.P.; Chen, Y.Q. MicroRNA-100/99a, deregulated in acute lymphoblastic leukaemia, suppress proliferation and promote apoptosis by regulating the FKBP51 and IGF1R/mTOR signalling pathways. Br. J. Cancer 2013, 109, 2189–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Mo, D.; Zhang, Y.; Zhang, Y.; Cong, P.; Xiao, S.; He, Z.; Liu, X.; Chen, Y. MicroRNAome comparison between intramuscular and subcutaneous vascular stem cell adipogenesis. PLoS ONE 2012, 7, e45410. [Google Scholar] [CrossRef]
- Li, G.; Li, Y.; Li, X.; Ning, X.; Li, M.; Yang, G. MicroRNA identity and abundance in developing swine adipose tissue as determined by Solexa sequencing. J. Cell. Biochem. 2011, 112, 1318–1328. [Google Scholar] [CrossRef]
- Viswanathan, S.R.; Daley, G.Q.; Gregory, R.I. Selective blockade of microRNA processing by Lin28. Science 2008, 320, 97–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viswanathan, S.R.; Powers, J.T.; Einhorn, W.; Hoshida, Y.; Ng, T.L.; Toffanin, S.; O’Sullivan, M.; Lu, J.; Phillips, L.A.; Lockhart, V.L.; et al. Lin28 promotes transformation and is associated with advanced human malignancies. Nat. Genet. 2009, 41, 843–848. [Google Scholar] [CrossRef] [Green Version]
- Lightfoot, H.L.; Bugaut, A.; Armisen, J.; Lehrbach, N.J.; Miska, E.A.; Balasubramanian, S. A LIN28-dependent structural change in pre-let-7g directly inhibits dicer processing. Biochemistry 2011, 50, 7514–7521. [Google Scholar] [CrossRef] [Green Version]
- Michlewski, G.; Caceres, J.F. Post-transcriptional control of miRNA biogenesis. RNA 2019, 25, 1–16. [Google Scholar]
- Kaczkowski, B.; Torarinsson, E.; Reiche, K.; Havgaard, J.H.; Stadler, P.F.; Gorodkin, J. Structural profiles of human miRNA families from pairwise clustering. Bioinformatics 2009, 25, 291–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathelier, A.; Carbone, A. Large scale chromosomal mapping of human microRNA structural clusters. Nucleic Acids Res. 2013, 41, 4392–4408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otton, R.; Bolin, A.P.; Ferreira, L.T.; Marinovic, M.P.; Rocha, A.L.S.; Mori, M.A. Polyphenol-rich green tea extract improves adipose tissue metabolism by down-regulating miR-335 expression and mitigating insulin resistance and inflammation. J. Nutr. Biochem. 2018, 57, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Chen, L.; Shi, C.M.; Xu, G.F.; Xu, L.L.; Zhu, L.L.; Guo, X.R.; Ni, Y.; Cui, Y.; Ji, C. MiR-335, an adipogenesis-related microRNA, is involved in adipose tissue inflammation. Cell Biochem. Biophys. 2014, 68, 283–290. [Google Scholar] [CrossRef]
- Conickx, G.; Avila Cobos, F.; van den Berge, M.; Faiz, A.; Timens, W.; Hiemstra, P.S.; Joos, G.F.; Brusselle, G.G.; Mestdagh, P.; Bracke, K.R. microRNA profiling in lung tissue and bronchoalveolar lavage of cigarette smoke-exposed mice and in COPD patients: A translational approach. Sci. Rep. 2017, 7, 12871. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Control | Sensitized | log2FC | p corr |
|---|---|---|---|---|
| Downregulated | ||||
| rno-mir-30a | 497.871 | 135.472 | −1.878 | 0.000 |
| rno-mir-181d | 17.772 | 5.331 | −1.737 | 0.003 |
| rno-mir-21 | 313.798 | 96.820 | −1.696 | 0.000 |
| rno-mir-30e | 51.538 | 17.866 | −1.528 | 0.001 |
| rno-miR-194-5p | 79.663 | 28.303 | −1.493 | 0.022 |
| rno-mir-181b-2 | 42.018 | 15.548 | −1.434 | 0.003 |
| rno-mir-140 | 224.135 | 83.272 | −1.428 | 0.000 |
| rno-mir-140 (isomiR) | 16.408 | 6.139 | −1.418 | 0.035 |
| rno-miR-99a-5p | 2355.335 | 920.727 | −1.355 | 0.000 |
| rno-mir-140 (isomiR) | 71.921 | 31.032 | −1.213 | 0.024 |
| rno-mir-10b (isomiR) | 8594.443 | 3779.265 | −1.185 | 0.000 |
| rno-mir-10b (isomiR) | 5186.707 | 2673.096 | −0.956 | 0.002 |
| rno-mir-10b (isomiR) | 17436.284 | 9038.696 | −0.948 | 0.000 |
| rno-mir-10b | 389984.419 | 207561.930 | −0.910 | 0.002 |
| rno-miR-25-3p | 1095.843 | 609.184 | −0.847 | 0.001 |
| rno-miR-203b-3p | 404.280 | 226.100 | −0.838 | 0.003 |
| rno-mir-148a | 1710.533 | 985.831 | −0.795 | 0.019 |
| rno-mir-101a | 592.118 | 383.474 | −0.627 | 0.042 |
| rno-miR-30a-5p | 1980.284 | 1332.122 | −0.572 | 0.032 |
| rno-miR-148a-3p | 848.973 | 632.695 | −0.424 | 0.044 |
| Upregulated | ||||
| rno-miR-22-3p | 42064.320 | 61447.574 | 0.547 | 0.047 |
| rno-mir-26a | 1845.464 | 3083.088 | 0.740 | 0.013 |
| rno-mir-203a | 80.248 | 134.653 | 0.747 | 0.025 |
| rno-miR-151-5p | 330.363 | 559.598 | 0.760 | 0.005 |
| rno-let-7e | 212.724 | 382.057 | 0.845 | 0.001 |
| rno-mir-125b-2 | 59.775 | 107.911 | 0.852 | 0.011 |
| rno-let-7d-5p | 198.981 | 361.600 | 0.862 | 0.000 |
| rno-mir-186 | 164.726 | 310.721 | 0.916 | 0.044 |
| rno-mir-28 | 33.449 | 65.084 | 0.960 | 0.034 |
| rno-miR-365-3p | 21.838 | 44.120 | 1.015 | 0.045 |
| rno-let-7d | 121.531 | 248.091 | 1.030 | 0.000 |
| rno-mir-151 (isomiR) | 63.952 | 139.470 | 1.125 | 0.000 |
| rno-mir-423 (isomiR) | 39.236 | 91.327 | 1.219 | 0.001 |
| rno-miR-423-3p | 31.664 | 76.350 | 1.270 | 0.011 |
| rno-mir-322-1 | 15.317 | 40.091 | 1.388 | 0.004 |
| rno-mir-322-2 | 11.173 | 36.720 | 1.717 | 0.000 |
| Term | Gene Count | Fold Enrichment | p corr | Genes |
|---|---|---|---|---|
| Mineral absorption | 7 | 6.293 | 0.001 | FTL1, SLC5A1, HMOX1, SLC26A9, MT1, SLC39A4, FTH1 |
| Lysosome | 11 | 3.037 | 0.003 | ATP6V0C, AP1S1, CLTB, HEXA, NAGA, GALC, ABCA2, CD164, ATP6V0A4, AP4S1, AP3B1 |
| Bacterial invasion of epithelial cells | 8 | 3.463 | 0.008 | ACTB, CAV3, CBLB, CLTB, HCLS1, PIK3R5, ARPC4, PXN |
| TGF-β signaling pathway | 8 | 3.262 | 0.011 | MAPK1, ACVR1B, ROCK1, TGFBR1, ZFYVE16, ZFYVE9, PITX2, ACVR1 |
| Tuberculosis | 12 | 2.312 | 0.014 | ATP6V0C, CIITA, MAPK1, CYP27B1, PPP3CB, CALM3, FCGR2A, ATP6V0A4, RFXANK, IFNGR1, CD74, IL10 |
| Osteoclast differentiation | 9 | 2.446 | 0.030 | MAP3K7, MAPK1, TGFBR1, PPP3CB, PIK3R5, FCGR2A, FOSL1, IFNGR1, CSF1R |
| Synaptic vesicle cycle | 6 | 3.393 | 0.031 | ATP6V0C, CLTB, CPLX1, AP2S1, ATP6V1G1, ATP6V0A4 |
| Dorso-ventral axis formation | 4 | 5.610 | 0.033 | NOTCH3, MAPK1, ETS2, NOTCH4 |
| FoxO signaling pathway | 9 | 2.320 | 0.039 | CCNB1, MAPK1, PRMT1, RBL2, ATG12, TGFBR1, PIK3R5, STAT3, IL10 |
| Endocytosis | 14 | 1.818 | 0.044 | CAV3, ARFGAP2, CLTB, USP8, AP2S1, TGFBR1, ARPC4, CBLB, DAB2, ZFYVE16, ZFYVE9, SPG21, BIN1, VPS25 |
| Leukocyte transendothelial migration | 8 | 2.397 | 0.048 | ACTB, RASSF5, ROCK1, PTK2B, CLDN1, PIK3R5, PXN, CLDN23 |
| Term | Gene Count | p corr | Fold Enrichment | Genes |
|---|---|---|---|---|
| GO:0030100~regulation of endocytosis | 3 | 0.007 | 24.021 | ARFGAP1, SYT4, RAB4B |
| GO:0021831~embryonic olfactory bulb interneuron precursor migration | 2 | 0.008 | 240.205 | ARX, RAC1 |
| GO:0043652~engulfment of apoptotic cell | 2 | 0.020 | 96.082 | BECN1, RAC1 |
| GO:0046929~negative regulation of neurotransmitter secretion | 2 | 0.020 | 96.082 | SYT4, PNKD |
| GO:0006366~transcription from RNA polymerase II promoter | 6 | 0.024 | 3.621 | HAND1, CCNH, GTF2H4, NEUROD1, DDX21, RUNX2 |
| GO:0032471~negative regulation of endoplasmic reticulum calcium ion concentration | 2 | 0.032 | 60.051 | BAK1, ATP2A1 |
| GO:0014049~positive regulation of glutamate secretion | 2 | 0.048 | 40.034 | SYT4, ADORA2A |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szczepankiewicz, D.; Langwiński, W.; Kołodziejski, P.; Pruszyńska-Oszmałek, E.; Sassek, M.; Nowakowska, J.; Chmurzyńska, A.; Nowak, K.W.; Szczepankiewicz, A. Allergic Inflammation Alters microRNA Expression Profile in Adipose Tissue in the Rat. Genes 2020, 11, 1034. https://doi.org/10.3390/genes11091034
Szczepankiewicz D, Langwiński W, Kołodziejski P, Pruszyńska-Oszmałek E, Sassek M, Nowakowska J, Chmurzyńska A, Nowak KW, Szczepankiewicz A. Allergic Inflammation Alters microRNA Expression Profile in Adipose Tissue in the Rat. Genes. 2020; 11(9):1034. https://doi.org/10.3390/genes11091034
Chicago/Turabian StyleSzczepankiewicz, Dawid, Wojciech Langwiński, Paweł Kołodziejski, Ewa Pruszyńska-Oszmałek, Maciej Sassek, Joanna Nowakowska, Agata Chmurzyńska, Krzysztof W. Nowak, and Aleksandra Szczepankiewicz. 2020. "Allergic Inflammation Alters microRNA Expression Profile in Adipose Tissue in the Rat" Genes 11, no. 9: 1034. https://doi.org/10.3390/genes11091034