Sex Chromosome Turnover in Bent-Toed Geckos (Cyrtodactylus)

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
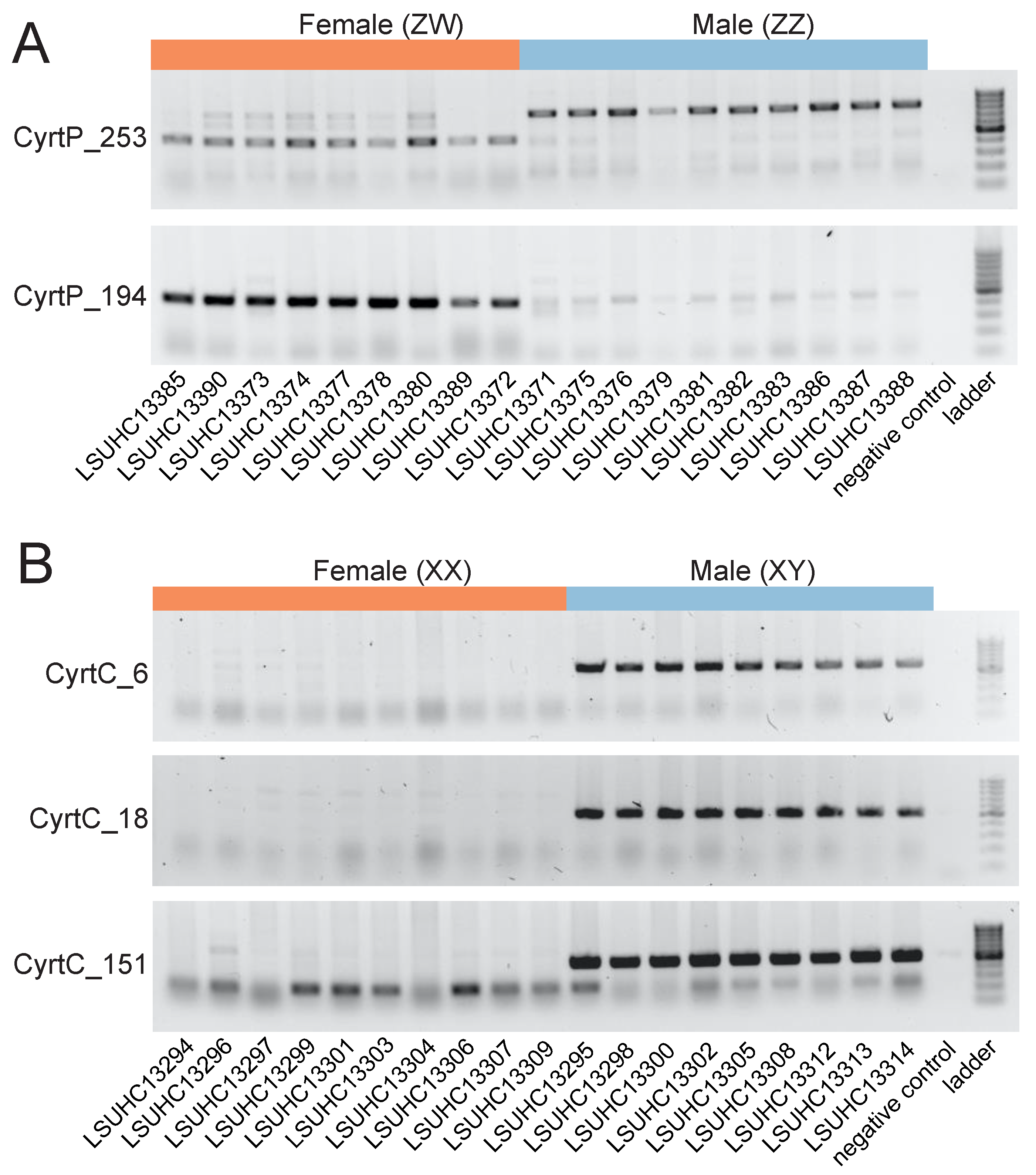
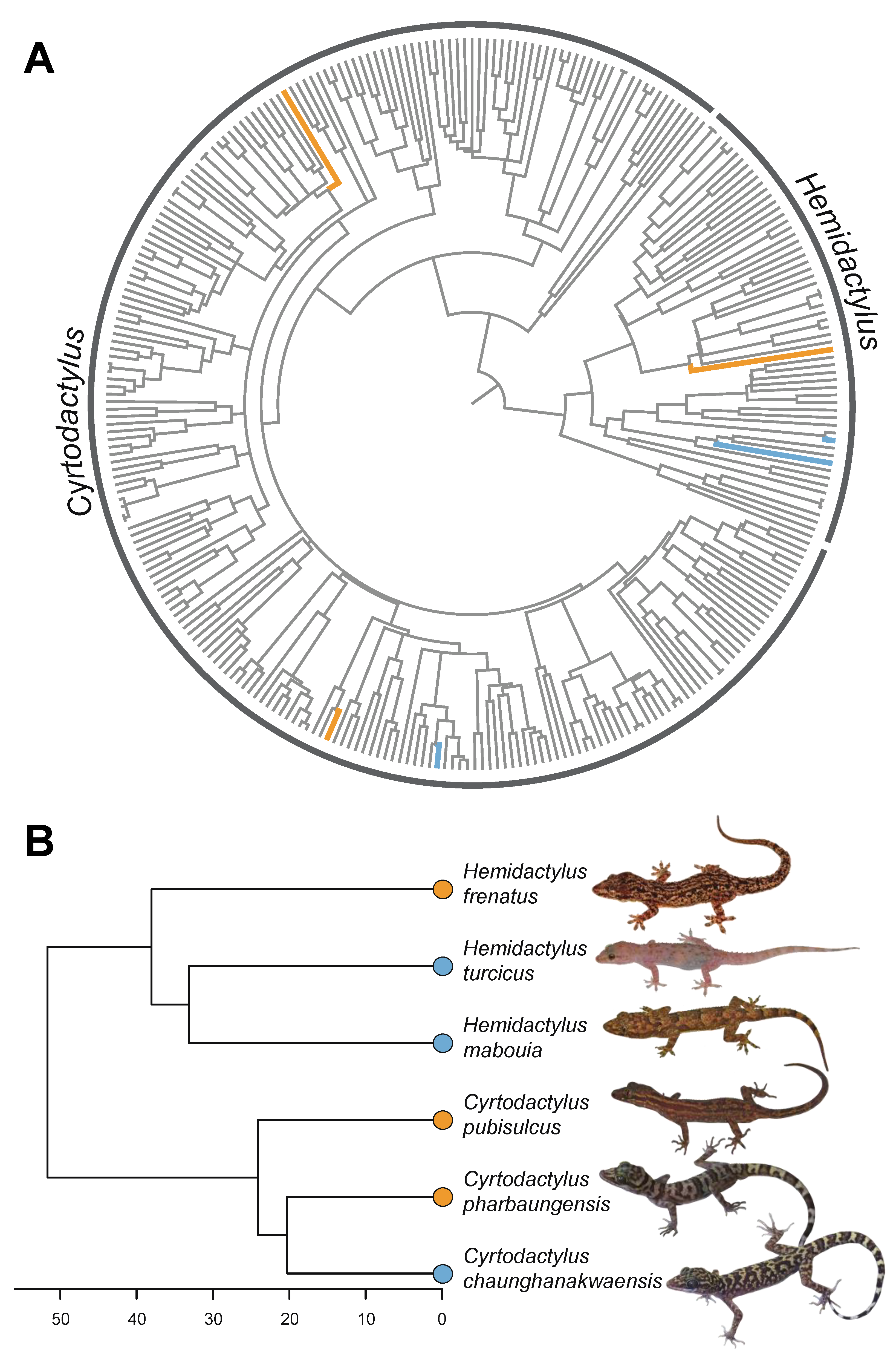
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ezaz, T.; Quinn, A.E.; Miura, I.; Sarre, S.D.; Georges, A.; Graves, J.A.M. The dragon lizard Pogona vitticeps has ZZ/ZW micro-sex chromosomes. Chromosome Res. 2005, 13, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Ezaz, T.; Sarre, S.; O’Meally, D.; Graves, J.M.; Georges, A. Sex chromosome evolution in lizards: Independent origins and rapid transitions. Cytogenet. Genome Res. 2009, 127, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Pokorna, M.; Kratochvíl, L. Phylogeny of sex-determining mechanisms in squamate reptiles: Are sex chromosomes an evolutionary trap? Zool. J. Linn. Soc. 2009, 156, 168–183. [Google Scholar] [CrossRef] [Green Version]
- Gamble, T.; Geneva, A.J.; Glor, R.E.; Zarkower, D. Anolis Sex chromosomes are derived from a single ancestral pair. Evolution 2014, 68, 1027–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamble, T.; Coryell, J.; Ezaz, T.; Lynch, J.; Scantlebury, D.P.; Zarkower, D. Restriction site-associated DNA sequencing (RAD-Seq) reveals an extraordinary number of transitions among gecko sex-determining systems. Mol. Biol. Evol. 2015, 32, 1296–1309. [Google Scholar] [CrossRef] [Green Version]
- Altmanová, M.; Rovatsos, M.; Pokorná, M.J.; Veselý, M.; Wagner, F.; Kratochvíl, L. All iguana families with the exception of basilisks share sex chromosomes. Zoology 2018, 126, 98–102. [Google Scholar] [CrossRef]
- Nielsen, S.V.; Banks, J.; Diaz, R., Jr.; Trainor, P.; Gamble, T. Dynamic sex chromosomes in Old World chameleons (Squamata: Chamaeleonidae). J. Evol. Biol. 2018, 31, 484–490. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, S.V.; Guzmán-Méndez, I.A.; Gamble, T.; Blumer, M.; Pinto, B.J.; Kratochvíl, L.; Rovatsos, M. Escaping the evolutionary trap? Sex chromosome turnover in basilisks and related lizards (Corytophanidae: Squamata). Biol. Lett. 2019, 15, 20190498. [Google Scholar] [CrossRef]
- Yoshida, M.; Itoh, M. Karyotype of the gekko Gekko Japonicus. Chrom. Inf. Serv. 1974, 17, 29–31. [Google Scholar]
- Solleder, E.; Schmid, M. XX/XY-Sex Chromosomes in Gekko gecko (Sauria, Reptilia). Amphib. Reptil. 1984, 5, 339–345. [Google Scholar] [CrossRef]
- Kawai, A.; Ishijima, J.; Nishida, C.; Kosaka, A.; Ota, H.; Kohno, S.; Matsuda, Y. The ZW sex chromosomes of Gekko hokouensis (Gekkonidae, Squamata) represent highly conserved homology with those of avian species. Chromosoma 2009, 118, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.L., Jr.; Heinicke, M.P.; Jackman, T.R.; Bauer, A.M. Phylogeny of bent-toed geckos (Cyrtodactylus) reveals a west to east pattern of diversification. Mol. Phylogenet. Evol. 2012, 65, 992–1003. [Google Scholar] [CrossRef] [PubMed]
- Das, I.; van Dijk, P.P. Species richness and endemicity of the herpetofauna of South and Southeast Asia. Raffles Bull. Zool. 2013, 29, 269–277. [Google Scholar]
- Brennan, I.G.; Bauer, A.M.; Van Tri, N.; Wang, Y.; Wang, W.; Zhang, Y.-P.; Murphy, R.W. Barcoding utility in a mega-diverse, cross-continental genus: Keeping pace with Cyrtodactylus geckos. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grismer, L.L.; Wood, P.J.; Thura, M.K.; Quah, E.S.; Grismer, M.S.; Murdoch, M.L.; Espinoza, R.E.; Lin, A. A new Cyrtodactylus Gray, 1827 (Squamata, Gekkonidae) from the Shan Hills and the biogeography of bent-toed geckos from Eastern Myanmar. Zootaxa 2018, 4446, 477–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grismer, L.L.; Wood, P.L., Jr.; Le, M.D.; Quah, E.S.; Grismer, J.L. Evolution of habitat preference in 243 species of bent-toed geckos (genus Cyrtodactylus Gray, 1827) with a discussion of karst habitat conservation. Ecol. Evol. 2020, 10. [Google Scholar] [CrossRef]
- Grismer, L.L.; Wood, P.L., Jr.; Poyarkov, N.A.; Le, M.D.; Kraus, F.; Agarwal, I.; Oliver, P.M.; Nguyen, S.N.; Nguyen, T.Q.; Karunarathna, S.; et al. Phylogenetic partitioning of the third-largest vertebrate genus in the world, Cyrtodactylus Gray, 1827 (Reptilia; Squamata; Gekkonidae) and its relevance to taxonomy and conservation. Vertebr. Zool. 2021, accepted. [Google Scholar]
- Ota, H.; Hikida, T.; Matsui, M.; Mori, A. Karyotypes of two species of the genus Cyrtodactylus (Squamata: Gekkonidae) from Sarawak, Malaysia. Caryologia 1992, 45, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Thongnetr, W.; Tanomtong, A.; Prasopsin, S.; Maneechot, N.; Pinthong, K.; Patawang, I. Cytogenetic study of the bent-toed gecko (Reptilia, Gekkonidae) in Thailand; I: Chromosomal classical features and NORs characterization of Cyrtodactylus Kunyai and C. Interdigitalis. Caryologia 2019, 72, 23–28. [Google Scholar] [CrossRef]
- Gamble, T.; Greenbaum, E.; Jackman, T.R.; Russell, A.P.; Bauer, A.M. Repeated origin and loss of adhesive toepads in geckos. PLoS ONE 2012, 7, e39429. [Google Scholar] [CrossRef] [Green Version]
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Currey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresko, W.A.; Johnson, E.A. Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef] [PubMed]
- Grismer, L.L.; Wood, P.L., Jr.; Thura, M.K.; Quah, E.S.; Murdoch, M.L.; Grismer, M.S.; Herr, M.W.; Lin, A.; Kyaw, H. Three more new species of Cyrtodactylus (Squamata: Gekkonidae) from the Salween Basin of Eastern Myanmar underscore the urgent need for the conservation of Karst Habitats. J. Nat. Hist. 2018, 52, 1243–1294. [Google Scholar] [CrossRef]
- Grismer, L.L.; Wood, P.L., Jr.; Thura, M.K.; Zin, T.; Quah, E.S.; Murdoch, M.L.; Grismer, M.S.; Lin, A.; Kyaw, H.; Lwin, N. Twelve new species of Cyrtodactylus Gray (Squamata: Gekkonidae) from isolated limestone habitats in East-Central and Southern Myanmar demonstrate high localized diversity and unprecedented microendemism. Zool. J. Linn. Soc. 2018, 182, 862–959. [Google Scholar] [CrossRef] [Green Version]
- Etter, P.D.; Bassham, S.; Hohenlohe, P.A.; Johnson, E.A.; Cresko, W.A. SNP discovery and genotyping for evolutionary genetics using RAD sequencing. In Molecular Methods for Evolutionary Genetics; Humana Press: Totowa, NJ, USA, 2012; Volume 772, pp. 157–178. [Google Scholar]
- Rohland, N.; Reich, D. Cost-effective, high-throughput DNA sequencing libraries for multiplexed target capture. Genome Res. 2012, 22, 939–946. [Google Scholar] [CrossRef] [Green Version]
- Catchen, J.M.; Amores, A.; Hohenlohe, P.; Cresko, W.; Postlethwait, J.H. Stacks: Building and genotyping loci de novo from short-read sequences. G3 2011, 1, 171–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, S.W.; Davey, J.W.; Johnston, J.S.; Shelton, A.M.; Heckel, D.G.; Jiggins, C.D.; Blaxter, M.L. Linkage mapping and comparative genomics using next-generation RAD sequencing of a non-model organism. PLoS ONE 2011, 6, e19315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Pinto, B.J.; Nielsen, S.V.; Gamble, T. Transcriptomic data support a nocturnal bottleneck in the ancestor of gecko lizards. Mol. Phylogenet. Evol. 2019, 141, 106639. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- International Chicken Genome Sequencing Consortium. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar] [CrossRef] [PubMed]
- Warren, W.C.; Hillier, L.W.; Tomlinson, C.; Minx, P.; Kremitzki, M.; Graves, T.; Markovic, C.; Bouk, N.; Pruitt, K.D.; Thibaud-Nissen, F. A new chicken genome assembly provides insight into avian genome structure. G3 2017, 7, 109–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graves, J.A.M.; Peichel, C.L. Are homologies in vertebrate sex determination due to shared ancestry or to limited options? Genome Biol. 2010, 11, 205. [Google Scholar] [CrossRef] [PubMed]
- Kawai, A.; Nishida-Umehara, C.; Ishijima, J.; Tsuda, Y.; Ota, H.; Matsuda, Y. Different origins of bird and reptile sex chromosomes inferred from comparative mapping of chicken Z-linked genes. Cytogenet. Genome Res. 2007, 117, 92–102. [Google Scholar] [CrossRef]
- Matsubara, K.; Tarui, H.; Toriba, M.; Yamada, K.; Nishida-Umehara, C.; Agata, K.; Matsuda, Y. Evidence for different origin of sex chromosomes in snakes, birds, and mammals and step-wise differentiation of snake sex chromosomes. Proc. Natl. Acad. Sci. USA 2006, 103, 18190–18195. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, Y.; Nishida-Umehara, C.; Tarui, H.; Kuroiwa, A.; Yamada, K.; Isobe, T.; Ando, J.; Fujiwara, A.; Hirao, Y.; Nishimura, O. Highly conserved linkage homology between birds and turtles: Bird and turtle chromosomes are precise counterparts of each other. Chromosome Res. 2005, 13, 601–615. [Google Scholar] [CrossRef]
- Alföldi, J.; Di Palma, F.; Grabherr, M.; Williams, C.; Kong, L.; Mauceli, E.; Russell, P.; Lowe, C.B.; Glor, R.E.; Jaffe, J.D. The genome of the green anole lizard and a comparative analysis with birds and mammals. Nature 2011, 477, 587–591. [Google Scholar] [CrossRef] [Green Version]
- Pokorná, M.; Giovannotti, M.; Kratochvíl, L.; Caputo, V.; Olmo, E.; Ferguson-Smith, M.A.; Rens, W. Conservation of chromosomes syntenic with avian autosomes in squamate reptiles revealed by comparative chromosome painting. Chromosoma 2012, 121, 409–418. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- To, T.-H.; Jung, M.; Lycett, S.; Gascuel, O. Fast dating using least-squares criteria and algorithms. Syst. Biol. 2016, 65, 82–97. [Google Scholar] [CrossRef] [PubMed]
- Gamble, T.; Greenbaum, E.; Jackman, T.R.; Bauer, A.M. Into the light: Diurnality has evolved multiple times in geckos. Biol. J. Linn. Soc 2015, 115, 896–910. [Google Scholar] [CrossRef] [Green Version]
- Gamble, T.; McKenna, E.; Meyer, W.; Nielsen, S.V.; Pinto, B.J.; Scantlebury, D.P.; Higham, T.E. XX/XY sex chromosomes in the South American dwarf gecko (Gonatodes Humeralis). J. Hered. 2018, 109, 462–468. [Google Scholar] [CrossRef]
- Kachitvichyanukul, V.; Schmeiser, B. Computer generation of hypergeometric random variates. J. Stat. Comput. Simul. 1985, 22, 127–145. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016; ISBN 3-319-24277-6. [Google Scholar]
- Wickham, H. Reshaping data with the reshape package. J. Stat. Softw. 2007, 21, 1–20. [Google Scholar] [CrossRef]
- O’Connell, K.A.; Smart, U.; Sidik, I.; Riyanto, A.; Kurniawan, N.; Smith, E.N. Diversification of bent-toed geckos (Cyrtodactylus) on Sumatra and West Java. Mol. Phylogenet. Evol. 2019, 134, 1–11. [Google Scholar] [CrossRef]
- Carranza, S.; Arnold, E. Systematics, biogeography, and evolution of Hemidactylus geckos (Reptilia: Gekkonidae) elucidated using mitochondrial DNA sequences. Mol. Phylogenet. Evol. 2006, 38, 531–545. [Google Scholar] [CrossRef]
- Moritz, C. Patterns and processes of sex chromosome evolution in gekkonid lizards (Sauria: Reptilia). In Cytogenetics of Amphibians and Reptiles; Birkhäuser Verlag: Basel, Switzerland, 1990; pp. 205–219. [Google Scholar]
- Gamble, T. A review of sex determining mechanisms in geckos (Gekkota: Squamata). Sex. Dev. 2010, 4, 88–103. [Google Scholar] [CrossRef] [Green Version]
- Grismer, L.L.; Quah, E.S.; Thura, M.K. Origin, diversity, and conservation of karst-associated bent-toed geckos (Genus Cyrtodactylus) in Myanmar (Burma). Isr. J. Ecol. Evol. 2020, 1, 1–7. [Google Scholar] [CrossRef]
- Uetz, P.; Hošek, J.; Hallermann, J. The Reptile Database. 2020. Available online: http://www.reptile-database.org/ (accessed on 1 December 2020).
- O’Meally, D.; Ezaz, T.; Georges, A.; Sarre, S.D.; Graves, J.A.M. Are some chromosomes particularly good at sex? Insights from amniotes. Chromosome Res. 2012, 20, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Brelsford, A.; Lavanchy, G.; Sermier, R.; Rausch, A.; Perrin, N. Identifying homomorphic sex chromosomes from wild-caught adults with limited genomic resources. Mol. Ecol. Res. 2017, 17, 752–759. [Google Scholar] [CrossRef] [Green Version]
- Kostmann, A.; Kratochvil, L.; Rovatsos, M. Poorly differentiated XX/XY sex chromosomes are widely shared across skink radiation. BioRxiv 2020. [Google Scholar] [CrossRef]
- Rovatsos, M.; Vukić, J.; Altmanová, M.; Johnson Pokorná, M.; Moravec, J.; Kratochvíl, L. Conservation of sex chromosomes in lacertid lizards. Mol. Ecol. 2016, 25, 3120–3126. [Google Scholar] [CrossRef] [PubMed]
- Iannucci, A.; Altmanová, M.; Ciofi, C.; Ferguson-Smith, M.; Milan, M.; Pereira, J.C.; Pether, J.; Rehák, I.; Rovatsos, M.; Stanyon, R. Conserved sex chromosomes and karyotype evolution in monitor lizards (Varanidae). Heredity 2019, 123, 215–227. [Google Scholar] [CrossRef]
- Rovatsos, M.; Rehák, I.; Velenský, P.; Kratochvíl, L. Shared ancient sex chromosomes in varanids, beaded lizards, and alligator lizards. Mol. Biol. Evol. 2019, 36, 1113–1120. [Google Scholar] [CrossRef]
- Rovatsos, M.; Pokorná, M.; Altmanová, M.; Kratochvíl, L. Cretaceous park of sex determination: Sex chromosomes are conserved across iguanas. Biol. Lett. 2014, 10, 20131093. [Google Scholar] [CrossRef]
- Vicoso, B.; Emerson, J.; Zektser, Y.; Mahajan, S.; Bachtrog, D. Comparative sex chromosome genomics in snakes: Differentiation, evolutionary strata, and lack of global dosage compensation. PLoS Biol. 2013, 11, e1001643. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Species | Primer Name | Sequence (5′ to 3′) |
|---|---|---|
| Cyrtodactylus chaunghanakwaensis | CyrtC_6-F | TCAGCCCTATATGCAACGGATC |
| CyrtC_6-R | TGTCCCTTCAGTTGGTCCAAAA | |
| CyrtC_18-F | CCCGGTTAACTCTAGTCGCATT | |
| CyrtC-18-R | TGAGGGGTAGGCAAGATAAGGA | |
| CyrtC-151-F | CAGACTTGTCACTCACCCTGAA | |
| CyrtC_151-R | TGACTTCTCCTCATCTGGCAAC | |
| Cyrtodactylus pharbaungensis | CyrtP_194-F | AATCAGGCGACCTTTAAGCTCA |
| CyrtP_194-R | TGCAGACGTGATGTAAGGGAAA | |
| CyrtP_253-F | CTCAGTGGCTCCCTCGTTAATT | |
| CyrtP_253-R | TCTGTGTGGACTTTTTGGACCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keating, S.E.; Blumer, M.; Grismer, L.L.; Lin, A.; Nielsen, S.V.; Thura, M.K.; Wood, P.L., Jr.; Quah, E.S.H.; Gamble, T. Sex Chromosome Turnover in Bent-Toed Geckos (Cyrtodactylus). Genes 2021, 12, 116. https://doi.org/10.3390/genes12010116
Keating SE, Blumer M, Grismer LL, Lin A, Nielsen SV, Thura MK, Wood PL Jr., Quah ESH, Gamble T. Sex Chromosome Turnover in Bent-Toed Geckos (Cyrtodactylus). Genes. 2021; 12(1):116. https://doi.org/10.3390/genes12010116
Chicago/Turabian StyleKeating, Shannon E., Madison Blumer, L. Lee Grismer, Aung Lin, Stuart V. Nielsen, Myint Kyaw Thura, Perry L. Wood, Jr., Evan S. H. Quah, and Tony Gamble. 2021. "Sex Chromosome Turnover in Bent-Toed Geckos (Cyrtodactylus)" Genes 12, no. 1: 116. https://doi.org/10.3390/genes12010116
APA StyleKeating, S. E., Blumer, M., Grismer, L. L., Lin, A., Nielsen, S. V., Thura, M. K., Wood, P. L., Jr., Quah, E. S. H., & Gamble, T. (2021). Sex Chromosome Turnover in Bent-Toed Geckos (Cyrtodactylus). Genes, 12(1), 116. https://doi.org/10.3390/genes12010116