Molecular Cloning, Structure and Phylogenetic Analysis of a Hemocyanin Subunit from the Black Sea Crustacean Eriphia verrucosa (Crustacea, Malacostraca)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. cDNA Cloning and Analyses of Hemocyanin from Eriphia verrucosa
2.3. 3D-Model of E. verrucosa Hemocyanin Subunit
2.4. Phylogenetic Analysis
3. Results
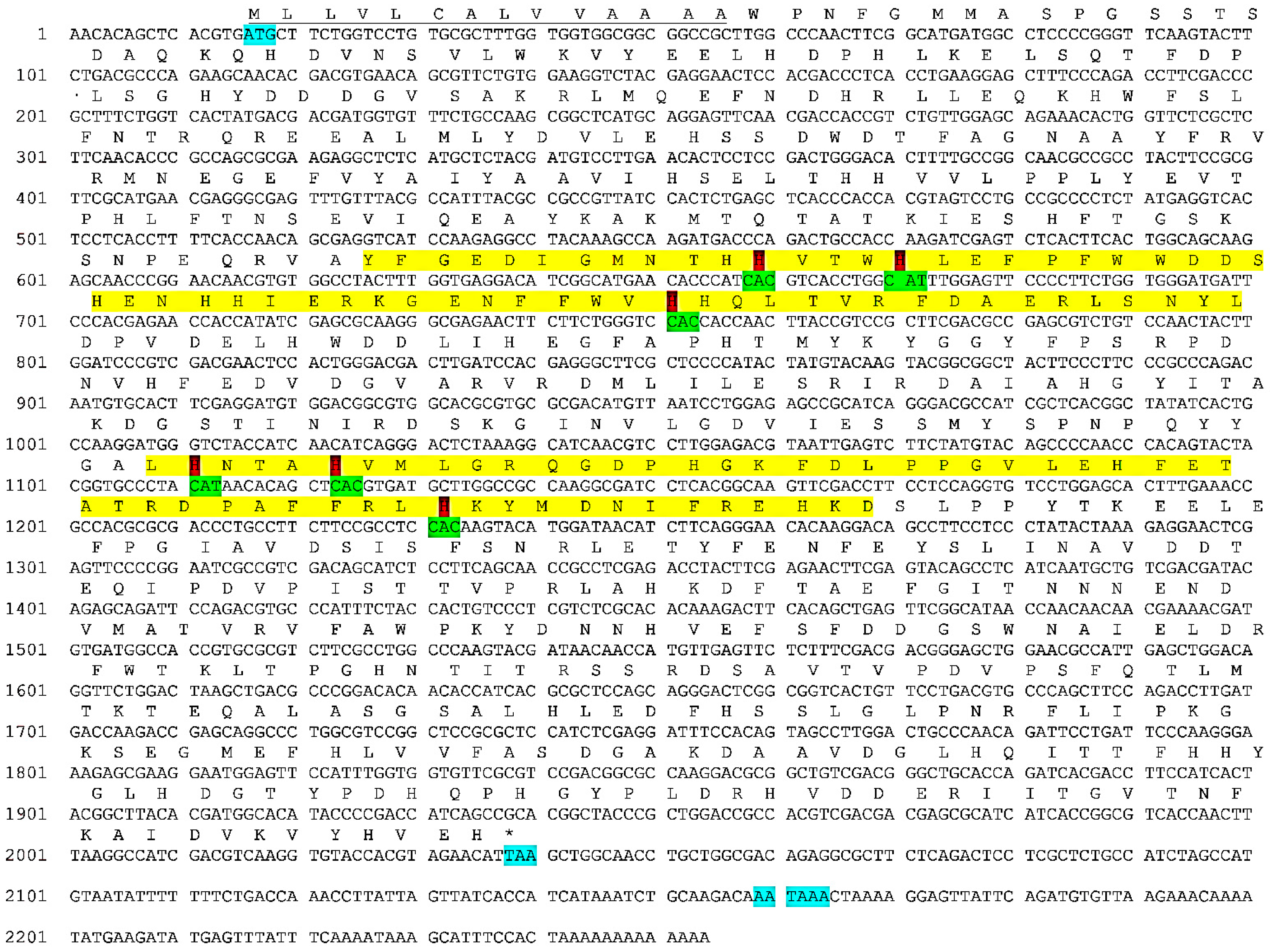
3.1. cDNA Sequence of E. verrucosa Hemocyanin Subunit
3.2. Homology and Phylogenetic Analysis of E. verrucosa Hemocyanin Subunit
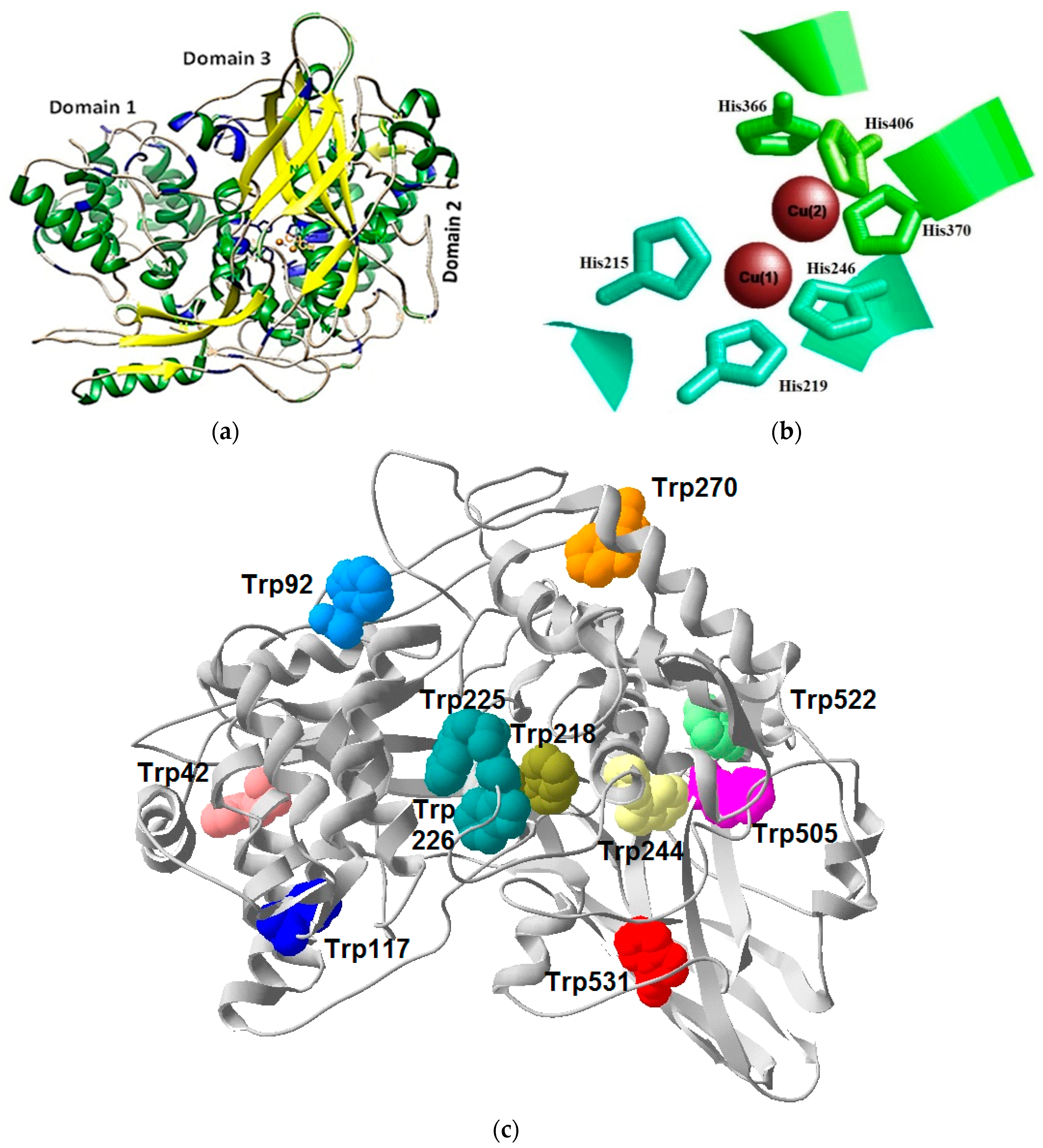
3.3. Structural Features of the E. verrucosa Hemocyanin Subunit 5
3.4. Glycosylation Sites in the E. verrucosa Hemocyanin Subunit 5
4. Discussion
4.1. Isolation, Structural, and Functional Properties of E. verrucosa Hemocyanin Subunit 5
4.2. Glycosylation of E. verrucosa Hemocyanin Subunit 5 and Possible Role in Immune Response
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Coates, C.J.; Nairn, J. Diverse immune functions of hemocyanins. Dev. Comp. Immunol. 2014, 45, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Markl, J. Evolution of molluscan hemocyanin structures. Biochim. Biophys. Acta 2013, 1834, 1840–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieb, B.; Markl, J. Evolution of molluscan hemocyanins as deduced from DNA sequencing. Micron 2004, 35, 117–119. [Google Scholar] [CrossRef] [PubMed]
- Burmester, T. Origin and evolution of arthropod hemocyanins and related proteins. J. Comp. Physiol. B 2002, 172, 95–117. [Google Scholar] [PubMed]
- Mangum, C.P.; Scott, J.L.; Black, R.E.L.; Miller, K.I.; van Holde, K.E. Centipedal hemocyanins: Its structure and implication for arthropod phylogeny. Proc. Natl. Acad. Sci. USA 1985, 82, 3721–3725. [Google Scholar] [CrossRef] [Green Version]
- Jaenicke, E.; Decker, H.; Gebauer, W.; Markl, J.; Burmester, T. Identification, structure and properties of hemocyanins from diplopod Myriapoda. J. Biol. Chem. 1999, 274, 29071–29074. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, D.; Ganfornina, M.D.; Gutierrez, G.; Bastiani, M.J. Molecular characterization and phylogenetic relationship of a protein with oxygen-binding capabilities in the grasshopper embryo. A hemocyanin in insects? Mol. Biol. Evol. 1998, 15, 415–426. [Google Scholar] [CrossRef] [Green Version]
- Hagner-Holler, S.; Schoen, A.; Erker, W.; Marden, J.H.; Rupprecht, R.; Decker, H.; Burmester, T. A respiratory hemocyanin from an insect. Proc. Natl. Acad. Sci. USA 2004, 101, 871–874. [Google Scholar] [CrossRef] [Green Version]
- Ertas, B.; von Reumont, B.M.; Wägele, J.W.; Misof, B.; Burmester, T. Hemocyanin suggests a close relationship of Remipedia and Hexapoda. Mol. Biol. Evol. 2009, 26, 2711–2718. [Google Scholar] [CrossRef] [Green Version]
- Meissner, U.; Stohr, M.; Kusche, K.; Burmester, T.; Stark, H.; Harris, J.R.; Orlova, E.; Markl, J. Quaternary structure of the European spiny lobster (Palinurus elephas) 1×6-mer hemocyanin from cryoEM and amino acid sequence data. J. Mol. Biol. 2003, 325, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Markl, J.; Decker, H. Molecular structure of the arthropod hemocyanins. Adv. Comp. Environ. Phys. 1992, 13, 325–376. [Google Scholar]
- van Holde, K.; Miller, K. Hemocyanins. Adv. Protein Chem. 1995, 47, 1–81. [Google Scholar] [PubMed]
- Riciluca, K.C.T.; Borges, A.C.; Mello, J.F.R.; de Oliveira, U.C.; Serdan, D.C.; Florez-Ariza, A.; Chaparro, E.; Nishiyama, M.Y.; Cassago, A.; Junqueira-de-Azevedo, I.L.M.; et al. Myriapod haemocyanin: The first three-dimensional reconstruction of Scolopendra subspinipes and preliminary structural analysis of S. Viridicornis. Open Biol. 2020, 10, 190258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, A.G.; Depoix, F.; Stohr, M.; Meissner, U.; Hagner-Holler, S.; Hammouti, K.; Burmester, T.; Heyd, J.; Wriggers, W.; Markl, J. Å Cryo-EM Structure, Sequence Analysis, Molecular Modelling and Rigid-body Fitting Reveal the Interfaces between the eight intefaces. J. Mol. Biol. 2007, 366, 1332–1350. [Google Scholar] [CrossRef] [PubMed]
- Linzen, B.; Soeter, N.M.; Riggs, A.F.; Schnieder, H.J.; Schartau, W. The structure of arthropod hemocyanins. Science 1985, 229, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Neuteboom, B.; Jekel, P.A.; Beintema, J.J. Primary structure of hemocyanin subunit c from Panulirus interruptus. Eur. J. Biochem. 1992, 206, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Decker, H.; Hellmann, N.; Jaenicke, E.; Lieb, B.; Meissner, U.; Markl, J. Minireview: Recent progress in hemocyanin research. Integr. Comp. Biol. 2007, 47, 631–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havanapan, P.; Kanlaya, R.; Bourchookarn, A.; Krittanai, C.; Thongboonkerd, V. C-Terminal hemocyanin from hemocytes of Penaeus vannamei interacts with ERK1/2 and undergoes serine phosphorylation. J. Proteome Res. 2009, 8, 2476–2483. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, F.; Hu, Z.; Zhao, X.; Min, S.; Du, Z.; Zhao, S.; Ye, X.; Li, Y. Hemocyanin from shrimp Litopenaeus vannamei shows hemolytic activity. Fish. Shellfish Immunol. 2009, 27, 330–335. [Google Scholar] [CrossRef]
- Markl, J. Evolution and function of structurally diverse subunits in the respiratory protein hemocyanin from arthropods. Biol. Bull. 1986, 171, 90–115. [Google Scholar] [CrossRef]
- Burmester, T. Molecular evolution of the arthropod hemocyanin superfamily. Mol. Biol. Evol. 2001, 18, 184–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durstewitz, G.; Terwilliger, N.B. cDNA cloning of a developmentally regulated hemocyanin subunit in the crustacean Cancer magister and phylogenetic analysis of the hemocyanin gene family. Mol. Biol. Evol. 1997, 14, 266–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durstewitz, G.; Terwilliger, N.B. Developmental changes in hemocyanin expression in the Dungeness crab, Cancer magister. J. Biol. Chem. 1997, 272, 4347–4350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terwilliger, N.; Ryan, M.; Phyllips, M.R. Crustacean hemocyanin gene family and microarray studies of expression change during eco-physiological stress. Integr. Comp. Biol. 2006, 46, 991–999. [Google Scholar] [CrossRef] [Green Version]
- Kusche, K.; Burmester, T. Molecular cloning and evolution of lobster hemocyanin. Biochem. Biophys. Res. Commun. 2001, 282, 887–892. [Google Scholar] [CrossRef]
- Sellos, D.; Lemoine, S.; van Wormhoudt, A. Molecular cloning of hemocyanin cDNA from Penaeus vannamei (Crustacea, Decapoda): Structure, evolution and physiological aspects. Febs Lett. 1997, 407, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Dolashka-Angelova, P.; Dolashki, A.; Savvides, S.N.; Hristova, R.; Van Beeumen, J.; Voelter, W.; Devreese, B.; Weser, U.; Di Muro, P.; Salvato, B.; et al. Structure of hemocyanin subunit CaeSS2 of the crustacean Mediterranean crab Carcinus aestuarii. J. Biochem. 2005, 138, 303–312. [Google Scholar] [CrossRef]
- Sun, J.; Wang, B.J.; Sun, Z.J.; Li, X.H. cDNA cloning and sequence analysis of hemocyanin in Fenneropenaeus chinensis. Prog. Fish. Sci. 2010, 31, 80–89. [Google Scholar]
- Sun, S.M.; Chen, L.Q.; Qin, J.G.; Ye, J.Y. Molecular cloning, characterization and mRNA expression of copper-binding protein hemocyanin subunit in Chinese mitten crab, Eriocheir sinensis. Fish. Shellfish. Immunol. 2012, 33, 1222–1228. [Google Scholar] [CrossRef]
- Marxen, J.C.; Pick, C.; Kwiatkowski, M.; Burmester, T. Molecular characterization and evolution of haemocyanin from the two freshwater shrimps Caridina multidentata (Stimpson, 1860) and Atyopsis moluccensis (De Haan, 1849). J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2013, 183, 613–624. [Google Scholar] [CrossRef]
- Kong, Y.; Chen, L.; Ding, Z.; Qin, J.; Sun, S.; Wang, L.; Ye, J. Molecular cloning, characterization, and mRNA expression of hemocyanin subunit in oriental river prawn Macrobrachium nipponense. Int. J. Genom. 2016, 2019, 6404817. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.F.; Xia, X.C.; Liu, F.; Chen, X.; Yang, H.; Ning, Q. Cloning and characterization of the hemocyanin gene of prawn Macrobrachium nipponense. Turk. J. Biochem. 2012, 37, 348–355. [Google Scholar] [CrossRef]
- Wang, D.L.; Zuo, D.; Wang, L.-M.; Wang, Q.; Sun, T.; Zhao, Y.-L. Cloning and tissue expression of hemocyanin gene in Cherax quadricarinatus during white spot syndrome virus infection. Aquaculture 2013, 410, 216–224. [Google Scholar] [CrossRef]
- Wang, K.Z.; Wen, Y.L.; Ye, Z.C.; Wu, H.G.; Pan, J.Y. Hemocyanin-derived phenoloxidase activity is dependent on dodecameric structure in shrimp Litopenaeus vannamei. Arch. Biol. Sci. 2015, 67, 19–23. [Google Scholar] [CrossRef]
- Hagner-Holler, S.; Kusche, K.; Hembach, A.; Burmester, T. Biochemical and molecular characterisation of hemocyanin from the amphipod Gammarus roeseli: Complex pattern of hemocyanin subunit evolution in Crustacea. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2005, 175, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Besser, K.; Malyon, G.P.; Eborall, W.S.; Paro da Cunha, G.; Filgueiras, J.G.; Dowle, A.; Cruz Garcia, L.; Page, S.J.; Dupree, R.; Kern, M.; et al. Hemocyanin facilitates lignocellulose digestion by wood-boring marine crustaceans. Nat. Commun. 2018, 9, 5125. [Google Scholar] [CrossRef]
- Nicholas, K.B.; Nicholas, H.B., Jr.; Deerfield, D.W., II. GeneDoc: Analysis and visualization of genetic variation. Embnew. News 1997, 4, 14. [Google Scholar]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421–430. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, H.J.; Bernstein, F.C. RASMOL—Molecular Graphics Visualization Tool, ver.2.7.5.1, (17 July 2009); Philippe Valadon, RASTOP ver.1. Available online: http://www.RasMol.org (accessed on 3 August 2000).
- Wang, Q.; Canutescu, A.A.; Dunbrack, R.L., Jr. SCWRL and MolIDE: Computer programs for side-chain conformation prediction and homology modeling. Nat. Protoc. 2008, 3, 1832–1847. [Google Scholar] [CrossRef] [Green Version]
- Krivov, G.G.; Shapovalov, M.V.; Dunbrack, R.L., Jr. Improved prediction of protein side-chain conformations with SCWRL4. Proteins 2009, 77, 778–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling Using MODELLER. In Current Protocols in Bioinformatics; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; Volume 54, pp. 5.6.1–5.6.37. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF CHIMERA—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guex, N.; Peitsch, M.C. DeepView—Swiss-Pdb Viewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Bendtsen, J.D.; Nielsen, H.; von Heijne, G.; Brunak, S. Improved prediction of signal peptides-Signal P 3.0. J. Mol. Biol. 2004, 340, 783–795. [Google Scholar] [CrossRef]
- Terwilliger, N.B.; Ryan, M.; Towle, D. Evolution of novel functions: Cryptocyanin helps build new exoskeleton in Cancer magister. J. Exp. Biol. 2005, 208, 2467–2474. [Google Scholar] [CrossRef] [Green Version]
- Pinnow, P.; Fabrizius, A.; Pick, C.; Burmester, T. Identification and characterisation of hemocyanin of the fish louse Argulus (Crustacea: Branchiura). J. Comp. Physiol. B 2016, 186, 161–168. [Google Scholar] [CrossRef]
- Hazes, B.; Magnus, K.A.; Bonaventura, C.; Bonaventura, J.; Dauter, Z.; Kalk, K.H.; Hol, W.G.J. Crystal structure of deoxygenated Limulus polyphemus subunit II hemocyanin at 2.18A resolution: Clues for a mechanism for allosteric regulation. Protein Sci. 1993, 2, 597–619. [Google Scholar] [CrossRef] [Green Version]
- Dolashka-Angelova, P.; Beltramini, M.; Dolashki, A.; Salvato, B.; Hristova, R.; Voelter, W. Carbohydrate composition of Carcinus aestuarii hemocyanin. Arch. Biochem. Biophys. 2001, 389, 153–158. [Google Scholar] [CrossRef]
- Kizheva, Y.K.; Rasheva, I.K.; Petrova, M.N.; Milosheva-Ivanova, A.V.; Velkova, L.G.; Dolashka, P.A.; Dolashki, A.K.; Hristova, P.K. Antibacterial activity of crab haemocyanin against clinical pathogens. Biotech. Biotechnol. Equip. 2019, 33, 873–880. [Google Scholar] [CrossRef] [Green Version]
- Tseneklidou-Stoeter, D.; Gerwig, G.J.; Kamerling, J.P.; Spindler, K.D. Characterization of N-linked carbohydrate chains of the crayfish, Astacus leptodactylus hemocyanin. Biol. Chem. Hoppe Seyler 1995, 376, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Dolashki, A.; Radkova, M.; Todorovska, E.; Ivanov, M.; Stevanovic, S.; Molin, L.; Traldi, P.; Voelter, W.; Dolashka, P. Structure and Characterization of Eriphia verrucosa Hemocyanin. Mar. Biotechnol. 2015, 17, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Decker, H.; Terwilliger, N. Cops and robbers: Putative evolution of copper oxygen-binding proteins. J. Exp. Biol. 2000, 203, 1777–1782. [Google Scholar] [PubMed]
- van Holde, K.E.; Miller, K.I.; Decker, H. Hemocyanins and invertebrate evolution. J. Biol. Chem. 2001, 276, 15563–15566. [Google Scholar] [CrossRef] [Green Version]
- Scherbaum, S.; Ertas, B.; Gebauer, W.; Burmester, T. Characterization of hemocyanin from the peacock mantis shrimp Odontodactylus scyllarus (Malacostraca: Hoplocarida). J. Comp. Physiol. B 2010, 180, 1235–1245. [Google Scholar] [CrossRef]
- Kusche, K.; Hembach, A.; Milke, C.; Burmester, T. Molecular characterization and evolution of the hemocyanin from the European spiny lobster, Palinurus elephas. J. Comp. Physiol. B 2003, 173, 319–325. [Google Scholar] [CrossRef]
- Volbeda, A.; Hol, W.G.J. Crystal structure of hexameric haemocyanin from Panulirus interruptus refined at 3.2 Å resolution. J. Mol. Biol. 1989, 209, 249–279. [Google Scholar] [CrossRef]
- Magnus, K.A.; Ton-That, H.; Carpenter, J.E. Recent structural work on the oxygen transport protein hemocyanin. Chem. Rev. 1994, 94, 727–735. [Google Scholar] [CrossRef]
- Hazes, B.; Hol, W.G. Comparison of the hemocyanin beta-barrel with other Greek key beta-barrels: Possible importance of the “beta-zipper” in protein structure and folding. Proteins 1992, 12, 278–298. [Google Scholar] [CrossRef]
- Zhang, Y.-L.; Peng, B.; Li, H.; Yan, F.; Wu, H.-K.; Zhao, X.-L.; Lin, X.-M.; Min, S.-Y.; Gao, Y.-Y.; Wang, S.-Y.; et al. Terminal domain of hemocyanin, a major antimicrobial protein from Litopenaeus vannamei: Structural homology with immunoglobulins and molecular diversity. Front. Immunol. 2017, 8, 611. [Google Scholar] [CrossRef]
- Dolashka-Angelova, P.; Lieb, B.; Velkova, L.; Heilen, N.; Sandra, K.; Nikolaeva-Glomb, L.; Dolashki, A.; Galabov, A.S.; Van Beeumen, J.; Stevanovic, S.; et al. Identification of glycosylated sites in Rapana hemocyanin by mass spectrometry and gene sequence, and their antiviral effect. Bioconjug Chem. 2009, 20, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Dolashka, P.; Velkova, L.; Shishkov, S.; Kostova, K.; Dolashki, A.; Dimitrov, I.; Atanasov, B.; Devreese, B.; Voelter, W.; Van Beeumen, J. Glycan structures and antiviral effect of the structural subunit RvH2 of Rapana hemocyanin. Carbohydr. Res. 2010, 345, 2361–2367. [Google Scholar] [CrossRef] [PubMed]
- Hart, G.W.; Copeland, R.J. Glycomics hits the big time. Cell 2010, 143, 672–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostadinova, E.; Dolashka, P.; Velkova, L.; Dolashki, A.; Stevanovic, S.; Voelter, W.J. Positions of the glycans in molluscan hemocyanin, determined by fluorescence spectroscopy. J. Fluoresc. 2013, 23, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Voit, R.; Feldmaier-Fuchs, G.; Schweikardt, T.; Decker, H.; Burmester, T. Complete sequence of the 24-mer hemocyanin of the tarantula Eurypelma californicus. Structure and intramolecular evolution of the subunits. J. Biol. Chem. 2000, 275, 39339–39344. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Li, R.; Aweya, J.J.; Wang, F.; Zhong, M.; Zhang, Y. Identification and characterization of glycosylation sites on Litopenaeus vannamei hemocyanin. FEBS Lett. 2019, 593, 820–830. [Google Scholar] [CrossRef]
- Becker, M.I.; Fuentes, A.; Del Campo, M.; Manubens, A.; Nova, E.; Oliva, H.; Faunes, F.; Valenzuela, M.A.; Campos Vallette, M.; Aliaga, A. Immunodominant role of CCHA subunit of Concholepas hemocyanin is associated with unique biochemical properties. Int. Immunopharmacol. 2009, 9, 330–339. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence 5′–3′ | References |
|---|---|---|
| Degenerate primer F | GAACTTTTTTTTTGGGTTCATCATCAACTTAC | [22] |
| Degenerate primer R | TGTGTTCTCTGAAGATGTTATCCATGTACTT | [22] |
| Gene-specific primer F | AAGTACATGGATAACATCTTCAG | In this study |
| Gene-specific primer R1 | TTATGCCGAACTCAGCTGTGAAGT | In this study |
| Gene-specific primer R2 | AAGCATCACGTGAGCTGTGTTA | In this study |
| 3′ anchor-specific primer | GACCACGCGTATCGATGTCGACTTTTTTTTTTTTTTTTV where V = A,C,G | 5′ and 3′ RACE Kit, 2nd Generation |
| 5′ anchor-specific primer | GACCACGCGTATCGATGTCGACTTTTTTTTTTTTTTTTV where V = A,C,G | 5′ and 3′ RACE Kit, 2nd Generation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Todorovska, E.; Ivanov, M.; Radkova, M.; Dolashki, A.; Dolashka, P. Molecular Cloning, Structure and Phylogenetic Analysis of a Hemocyanin Subunit from the Black Sea Crustacean Eriphia verrucosa (Crustacea, Malacostraca). Genes 2021, 12, 93. https://doi.org/10.3390/genes12010093
Todorovska E, Ivanov M, Radkova M, Dolashki A, Dolashka P. Molecular Cloning, Structure and Phylogenetic Analysis of a Hemocyanin Subunit from the Black Sea Crustacean Eriphia verrucosa (Crustacea, Malacostraca). Genes. 2021; 12(1):93. https://doi.org/10.3390/genes12010093
Chicago/Turabian StyleTodorovska, Elena, Martin Ivanov, Mariana Radkova, Alexandar Dolashki, and Pavlina Dolashka. 2021. "Molecular Cloning, Structure and Phylogenetic Analysis of a Hemocyanin Subunit from the Black Sea Crustacean Eriphia verrucosa (Crustacea, Malacostraca)" Genes 12, no. 1: 93. https://doi.org/10.3390/genes12010093
APA StyleTodorovska, E., Ivanov, M., Radkova, M., Dolashki, A., & Dolashka, P. (2021). Molecular Cloning, Structure and Phylogenetic Analysis of a Hemocyanin Subunit from the Black Sea Crustacean Eriphia verrucosa (Crustacea, Malacostraca). Genes, 12(1), 93. https://doi.org/10.3390/genes12010093