
Expression of Cytokine-Coding Genes BMP8B, LEFTY1 and INSL5 Could Distinguish between Ulcerative Colitis and Crohn’s Disease



Abstract
1. Introduction
2. Materials and Methods
2.1. Microarray Data
2.2. Data Processing
2.3. Functional Analysis
2.4. Patients and Tissue Samples
2.5. RNA Isolation and Reverse Transcription
2.6. Preamplification and Quantitative Real Time PCR
2.7. Statistical Analysis of qPCR
3. Results
3.1. Differentially Expressed Genes in Ulcerative Colitis and Crohn’s Disease
3.2. Functional Enrichment Analysis
3.2.1. Biological Processes
3.2.2. Cellular Components
3.2.3. Molecular Functions
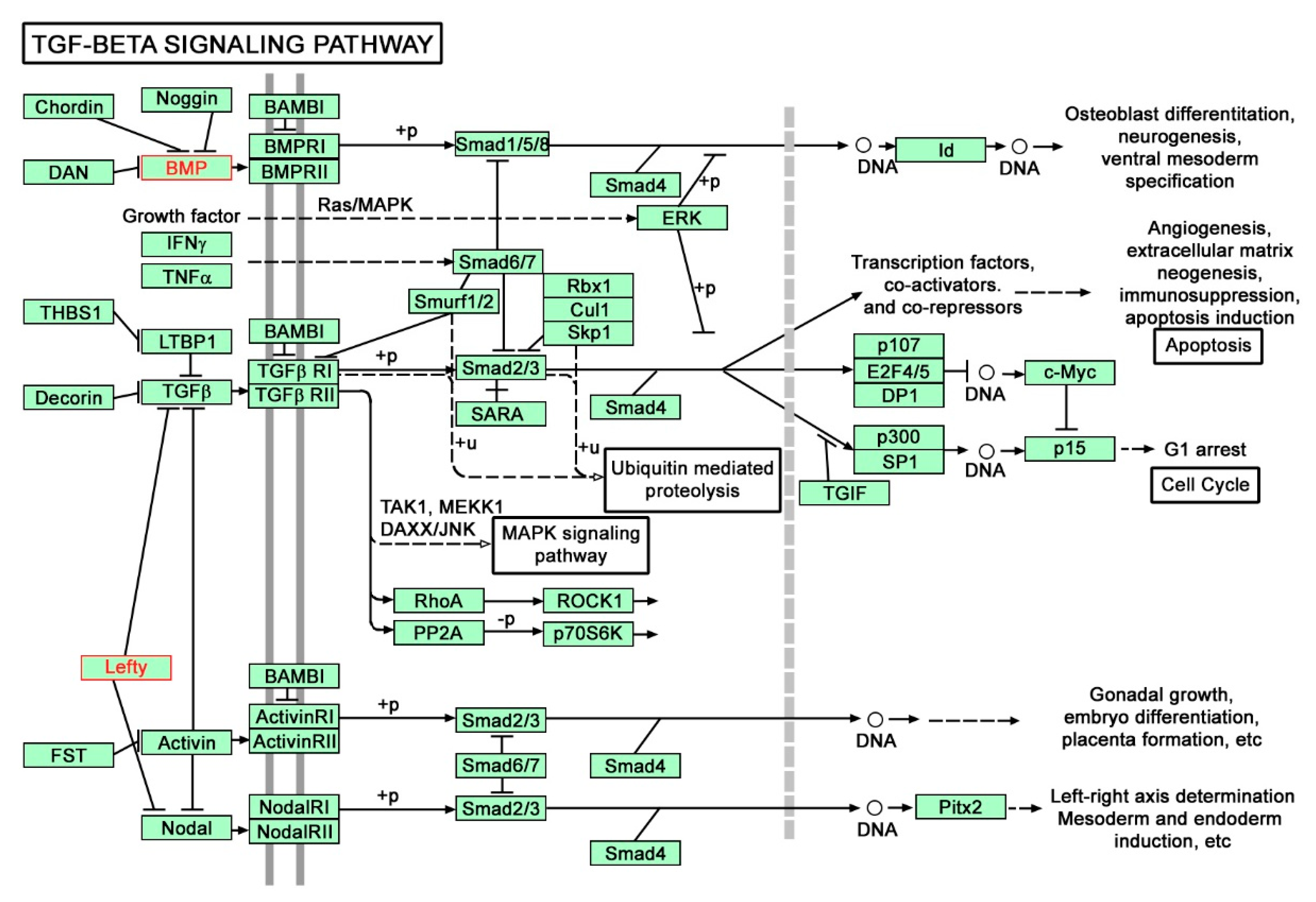
3.2.4. Pathway Analysis
3.2.5. Phenotypes
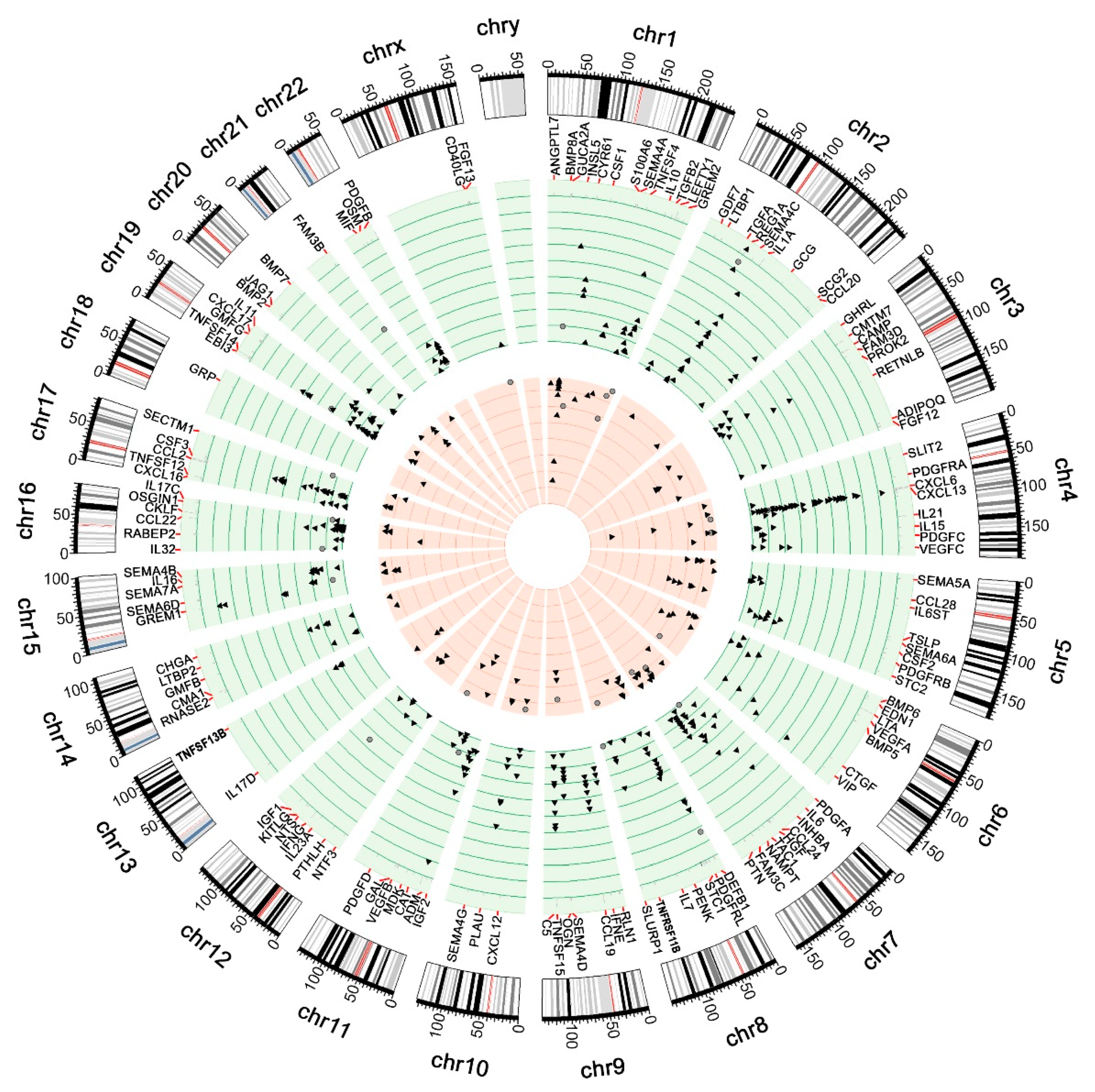
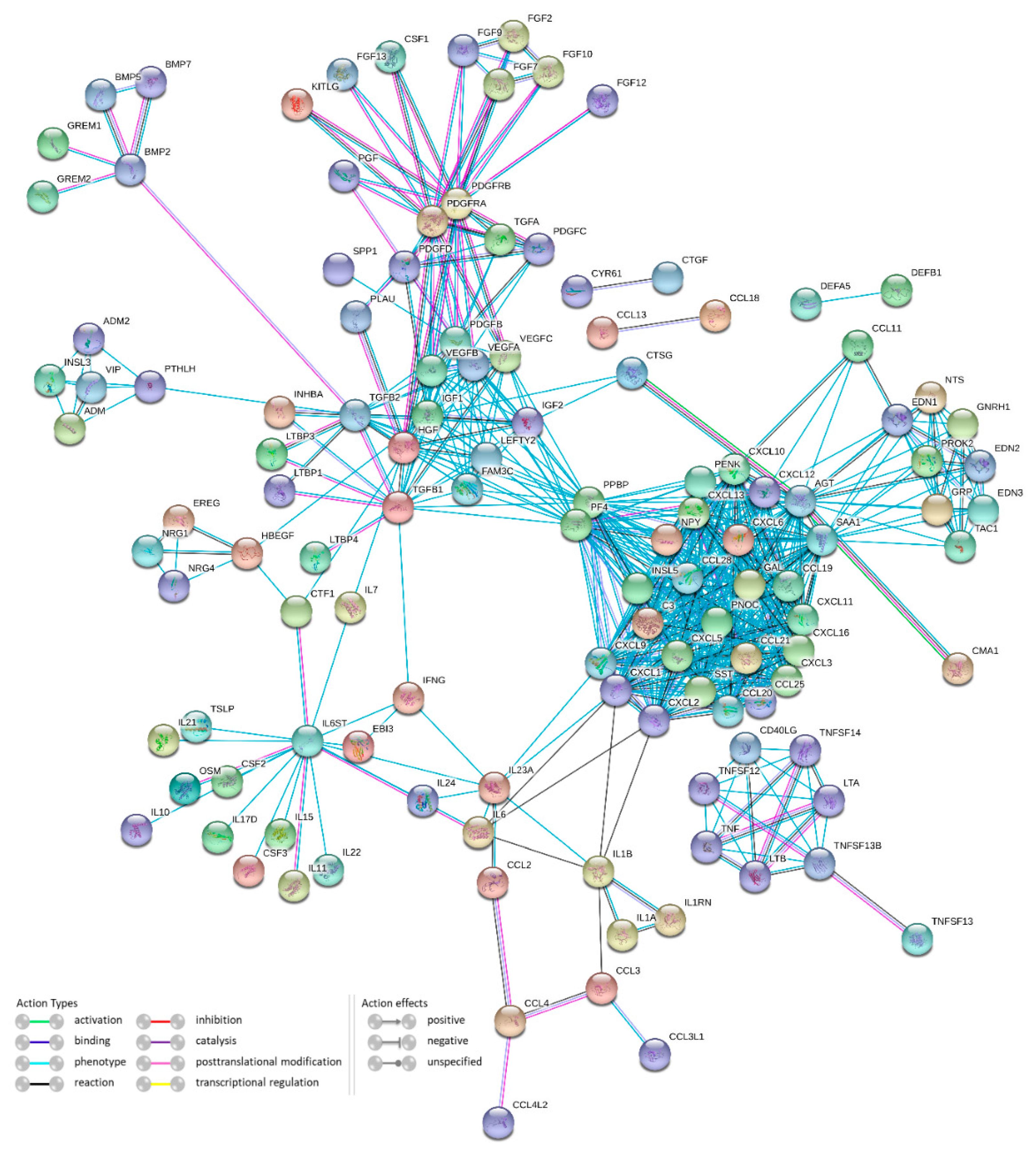
3.2.6. Protein-Protein Interactions Network
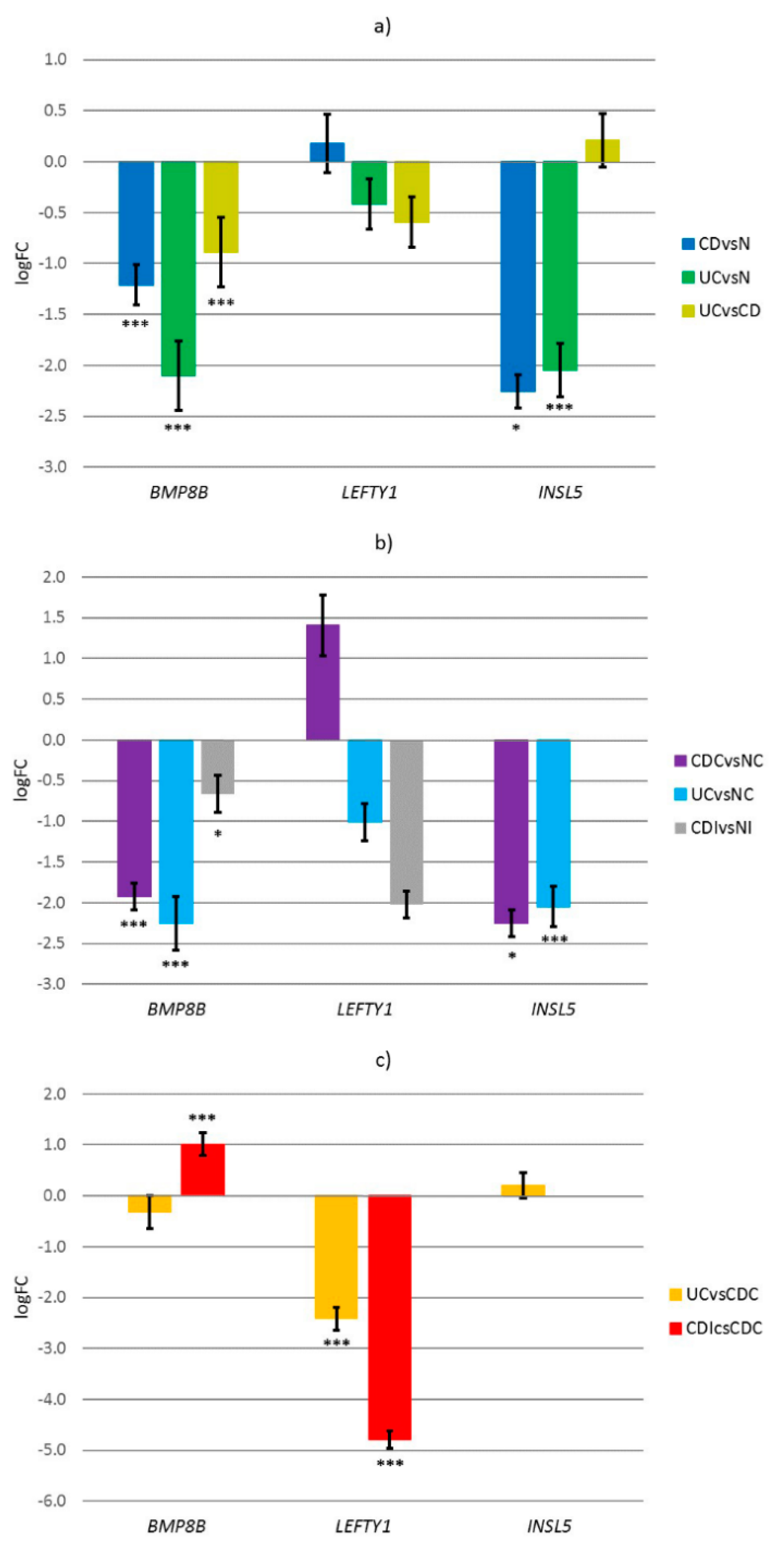
3.3. Experimental Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bouma, G.; Strober, W. The immunological and genetic basis of inflammatory bowel disease. Nat. Rev. Immunol. 2003, 3, 521–533. [Google Scholar] [CrossRef]
- Abraham, C.; Cho, J. Interleukin-23/Th17 pathways and inflammatory bowel disease. Inflamm. Bowel. Dis. 2009, 15, 1090–1100. [Google Scholar] [CrossRef]
- Neurath, M.F. Cytokines in inflammatory bowel disease. Nat. Rev. Immunol. 2014, 14, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Bamias, G.; Kaltsa, G.; Ladas, S.D. Cytokines in the pathogenesis of ulcerative colitis. Discov. Med. 2011, 11, 459–467. [Google Scholar] [PubMed]
- Bjerrum, J.T.; Hansen, M.; Olsen, J.; Nielsen, O.H. Genome-wide gene expression analysis of mucosal colonic biopsies and isolated colonocytes suggests a continuous inflammatory state in the lamina propria of patients with quiescent ulcerative colitis. Inflamm. Bowel. Dis. 2010, 16, 999–1007. [Google Scholar] [CrossRef]
- Van der Goten, J.; Vanhove, W.; Lemaire, K.; Van Lommel, L.; Machiels, K.; Wollants, W.J.; De Preter, V.; De Hertogh, G.; Ferrante, M.; Van Assche, G.; et al. Integrated miRNA and mRNA expression profiling in inflamed colon of patients with ulcerative colitis. PLoS ONE 2014, 9, e116117. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Fan, J.; Zhi, F.; Li, A.; Li, C.; Berger, A.E.; Boorgula, M.P.; Barkataki, S.; Courneya, J.P.; Chen, Y.; et al. Mobilization of epithelial mesenchymal transition genes distinguishes active from inactive lesional tissue in patients with ulcerative colitis. Hum. Mol. Genet. 2015, 24, 4615–4624. [Google Scholar] [CrossRef]
- Zhang, T.; DeSimone, R.A.; Jiao, X.; Rohlf, F.J.; Zhu, W.; Gong, Q.Q.; Hunt, S.R.; Dassopoulos, T.; Newberry, R.D.; Sodergren, E.; et al. Host genes related to paneth cells and xenobiotic metabolism are associated with shifts in human ileum-associated microbial composition. PLoS ONE 2012, 7, e30044. [Google Scholar]
- Peck, B.C.; Weiser, M.; Lee, S.E.; Gipson, G.R.; Iyer, V.B.; Sartor, R.B.; Herfarth, H.H.; Long, M.D.; Hansen, J.J.; Isaacs, K.L.; et al. MicroRNAs Classify Different Disease Behavior Phenotypes of Crohn’s Disease and May Have Prognostic Utility. Inflamm. Bowel. Dis. 2015, 21, 2178–2187. [Google Scholar] [CrossRef]
- Peters, L.A.; Perrigoue, J.; Mortha, A.; Iuga, A.; Song, W.M.; Neiman, E.M.; Llewellyn, S.R.; Di Narzo, A.; Kidd, B.A.; Telesco, S.E.; et al. A functional genomics predictive network model identifies regulators of inflammatory bowel disease. Nat. Genet. 2017, 49, 1437–1449. [Google Scholar] [CrossRef]
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. affy–analysis of Affymetrix GeneChip data at the probe level. Bioinformatics 2004, 20, 307–315. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Backes, C.; Keller, A.; Kuentzer, J.; Kneissl, B.; Comtesse, N.; Elnakady, Y.A.; Muller, R.; Meese, E.; Lenhof, H.P. GeneTrail--advanced gene set enrichment analysis. Nucleic Acids Res. 2007, 35, W186–W192. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Matsuura, M.; Okazaki, K.; Nishio, A.; Nakase, H.; Tamaki, H.; Uchida, K.; Nishi, T.; Asada, M.; Kawasaki, K.; Fukui, T.; et al. Therapeutic effects of rectal administration of basic fibroblast growth factor on experimental murine colitis. Gastroenterology 2005, 128, 975–986. [Google Scholar] [CrossRef]
- Bossard, C.; Souaze, F.; Jarry, A.; Bezieau, S.; Mosnier, J.F.; Forgez, P.; Laboisse, C.L. Over-expression of neurotensin high-affinity receptor 1 (NTS1) in relation with its ligand neurotensin (NT) and nuclear beta-catenin in inflammatory bowel disease-related oncogenesis. Peptides 2007, 28, 2030–2035. [Google Scholar] [CrossRef]
- Boyer, L.; Sidpra, D.; Jevon, G.; Buchan, A.M.; Jacobson, K. Differential responses of VIPergic and nitrergic neurons in paediatric patients with Crohn’s disease. Auton. Neurosci. 2007, 134, 106–114. [Google Scholar] [CrossRef]
- Jonsson, M.; Norrgard, O.; Hansson, M.; Forsgren, S. Decrease in binding for the neuropeptide VIP in response to marked inflammation of the mucosa in ulcerative colitis. Ann. N. Y. Acad. Sci. 2007, 1107, 280–289. [Google Scholar] [CrossRef]
- Tursi, A.; Elisei, W.; Inchingolo, C.D.; Nenna, R.; Picchio, M.; Ierardi, E.; Brandimarte, G. Chronic diverticulitis and Crohn’s disease share the same expression of basic fibroblastic growth factor, syndecan 1 and tumour necrosis factor-alpha. J. Clin. Pathol. 2014, 67, 844–846. [Google Scholar] [CrossRef]
- Tursi, A.; Elisei, W.; Principi, M.; Inchingolo, C.D.; Nenna, R.; Picchio, M.; Giorgio, F.; Ierardi, E.; Brandimarte, G. Mucosal expression of basic fibroblastic growth factor, syndecan 1 and tumour necrosis factor-alpha in Crohn’s disease in deep remission under treatment with anti-TNFalpha antibodies. J. Gastrointestin. Liver Dis. 2014, 23, 261–265. [Google Scholar] [CrossRef]
- Del Valle-Pinero, A.Y.; Sherwin, L.B.; Anderson, E.M.; Caudle, R.M.; Henderson, W.A. Altered vasoactive intestinal peptides expression in irritable bowel syndrome patients and rats with trinitrobenzene sulfonic acid-induced colitis. World J. Gastroenterol. 2015, 21, 155–163. [Google Scholar] [CrossRef]
- Wilkinson, T.N.; Speed, T.P.; Tregear, G.W.; Bathgate, R.A. Evolution of the relaxin-like peptide family. BMC Evol. Biol. 2005, 5, 14. [Google Scholar] [CrossRef]
- Liu, C.; Lovenberg, T.W. Relaxin-3, INSL5, and their receptors. Results Probl. Cell Differ. 2008, 46, 213–237. [Google Scholar]
- Conklin, D.; Lofton-Day, C.E.; Haldeman, B.A.; Ching, A.; Whitmore, T.E.; Lok, S.; Jaspers, S. Identification of INSL5, a new member of the insulin superfamily. Genomics 1999, 60, 50–56. [Google Scholar] [CrossRef]
- Bishop, A.E.; Pietroletti, R.; Taat, C.W.; Brummelkamp, W.H.; Polak, J.M. Increased populations of endocrine cells in Crohn’s ileitis. Virchows Arch. A Pathol. Anat. Histopathol. 1987, 410, 391–396. [Google Scholar] [CrossRef]
- Moran, G.W.; Pennock, J.; McLaughlin, J.T. Enteroendocrine cells in terminal ileal Crohn’s disease. J. Crohns Colitis 2012, 6, 871–880. [Google Scholar] [CrossRef]
- Hernandez-Trejo, J.A.; Suarez-Perez, D.; Gutierrez-Martinez, I.Z.; Fernandez-Vargas, O.E.; Serrano, C.; Candelario-Martinez, A.A.; Meraz-Rios, M.A.; Citalan-Madrid, A.F.; Hernandez-Ruiz, M.; Reyes-Maldonado, E.; et al. The pro-inflammatory cytokines IFNgamma/TNFalpha increase chromogranin A-positive neuroendocrine cells in the colonic epithelium. Biochem. J. 2016, 473, 3805–3818. [Google Scholar] [CrossRef]
- Oshima, S.; Fujimura, M.; Fukimiya, M. Changes in number of serotonin-containing cells and serotonin levels in the intestinal mucosa of rats with colitis induced by dextran sodium sulfate. Histochem. Cell Biol. 1999, 112, 257–263. [Google Scholar] [CrossRef]
- Linden, D.R.; Chen, J.X.; Gershon, M.D.; Sharkey, K.A.; Mawe, G.M. Serotonin availability is increased in mucosa of guinea pigs with TNBS-induced colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G207–G216. [Google Scholar] [CrossRef]
- O’Hara, J.R.; Ho, W.; Linden, D.R.; Mawe, G.M.; Sharkey, K.A. Enteroendocrine cells and 5-HT availability are altered in mucosa of guinea pigs with TNBS ileitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G998–G1007. [Google Scholar] [CrossRef]
- Thanasupawat, T.; Hammje, K.; Adham, I.; Ghia, J.E.; Del Bigio, M.R.; Krcek, J.; Hoang-Vu, C.; Klonisch, T.; Hombach-Klonisch, S. INSL5 is a novel marker for human enteroendocrine cells of the large intestine and neuroendocrine tumours. Oncol. Rep. 2013, 29, 149–154. [Google Scholar] [CrossRef]
- Hooper, L.V.; Macpherson, A.J. Immune adaptations that maintain homeostasis with the intestinal microbiota. Nat. Rev. Immunol. 2010, 10, 159–169. [Google Scholar] [CrossRef]
- Ihara, S.; Hirata, Y.; Koike, K. TGF-beta in inflammatory bowel disease: A key regulator of immune cells, epithelium, and the intestinal microbiota. J. Gastroenterol. 2017, 52, 777–787. [Google Scholar] [CrossRef]
- Monteleone, G.; Neurath, M.F.; Ardizzone, S.; Di Sabatino, A.; Fantini, M.C.; Castiglione, F.; Scribano, M.L.; Armuzzi, A.; Caprioli, F.; Sturniolo, G.C.; et al. Mongersen, an oral SMAD7 antisense oligonucleotide, and Crohn’s disease. N. Engl. J. Med. 2015, 372, 1104–1113. [Google Scholar] [CrossRef]
- Miettinen, P.J.; Ebner, R.; Lopez, A.R.; Derynck, R. TGF-beta induced transdifferentiation of mammary epithelial cells to mesenchymal cells: Involvement of type I receptors. J. Cell Biol. 1994, 127, 2021–2036. [Google Scholar] [CrossRef]
- Flier, S.N.; Tanjore, H.; Kokkotou, E.G.; Sugimoto, H.; Zeisberg, M.; Kalluri, R. Identification of epithelial to mesenchymal transition as a novel source of fibroblasts in intestinal fibrosis. J. Biol. Chem. 2010, 285, 20202–20212. [Google Scholar] [CrossRef]
- Zidar, N.; Bostjancic, E.; Jerala, M.; Kojc, N.; Drobne, D.; Stabuc, B.; Glavac, D. Down-regulation of microRNAs of the miR-200 family and up-regulation of Snail and Slug in inflammatory bowel diseases—Hallmark of epithelial-mesenchymal transition. J. Cell. Mol. Med. 2016, 20, 1813–1820. [Google Scholar] [CrossRef]
- Siegmund, B.; Feakins, R.M.; Barmias, G.; Ludvig, J.C.; Teixeira, F.V.; Rogler, G.; Scharl, M. Results of the Fifth Scientific Workshop of the ECCO (II): Pathophysiology of Perianal Fistulizing Disease. J. Crohns Colitis 2016, 10, 377–386. [Google Scholar] [CrossRef]
- Ulloa, L.; Tabibzadeh, S. Lefty inhibits receptor-regulated Smad phosphorylation induced by the activated transforming growth factor-beta receptor. J. Biol. Chem. 2001, 276, 21397–21404. [Google Scholar] [CrossRef]
- Yashiro, K.; Saijoh, Y.; Sakuma, R.; Tada, M.; Tomita, N.; Amano, K.; Matsuda, Y.; Monden, M.; Okada, S.; Hamada, H. Distinct transcriptional regulation and phylogenetic divergence of human LEFTY genes. Genes Cells 2000, 5, 343–357. [Google Scholar] [CrossRef]
- Miyata, N.; Azuma, T.; Hozawa, S.; Higuchi, H.; Yokoyama, A.; Kabashima, A.; Igarashi, T.; Saeki, K.; Hibi, T. Transforming growth factor beta and Ras/MEK/ERK signaling regulate the expression level of a novel tumor suppressor Lefty. Pancreas 2012, 41, 745–752. [Google Scholar] [CrossRef]
- Akiya, M.; Yamazaki, M.; Matsumoto, T.; Kawashima, Y.; Oguri, Y.; Kajita, S.; Kijima, D.; Chiba, R.; Yokoi, A.; Takahashi, H.; et al. Identification of LEFTY as a molecular marker for ovarian clear cell carcinoma. Oncotarget 2017, 8, 63646–63664. [Google Scholar] [CrossRef]
- Mima, K.; Fukagawa, T.; Kurashige, J.; Takano, Y.; Uchi, R.; Ueo, H.; Matsumura, T.; Ishibashi, M.; Sawada, G.; Takahashi, Y.; et al. Gene expression of bone morphogenic protein 8B in the primary site, peripheral blood and bone marrow of patients with gastric cancer. Oncol. Lett. 2013, 6, 387–392. [Google Scholar] [CrossRef]
- Wisnieski, F.; Leal, M.F.; Calcagno, D.Q.; Santos, L.C.; Gigek, C.O.; Chen, E.S.; Artigiani, R.; Demachki, S.; Assumpcao, P.P.; Lourenco, L.G.; et al. BMP8B Is a Tumor Suppressor Gene Regulated by Histone Acetylation in Gastric Cancer. J. Cell Biochem. 2017, 118, 869–877. [Google Scholar] [CrossRef]
- Davis, S.J.; Sheppard, K.E.; Anglesio, M.S.; George, J.; Traficante, N.; Fereday, S.; Intermaggio, M.P.; Menon, U.; Gentry-Maharaj, A.; Lubinski, J.; et al. Enhanced GAB2 Expression Is Associated with Improved Survival in High-Grade Serous Ovarian Cancer and Sensitivity to PI3K Inhibition. Mol. Cancer Ther. 2015, 14, 1495–1503. [Google Scholar] [CrossRef]
- Cheng, Z.; Cui, W.; Ding, Y.; Liu, T.; Liu, W.; Qin, Y.; Xia, W.; Xu, J.; Zhang, Y.; Zou, X. BMP8B mediates the survival of pancreatic cancer cells and regulates the progression of pancreatic cancer. Oncol. Rep. 2014, 32, 1861–1866. [Google Scholar] [CrossRef][Green Version]
- Zhao, G.Q.; Liaw, L.; Hogan, B.L. Bone morphogenetic protein 8A plays a role in the maintenance of spermatogenesis and the integrity of the epididymis. Development 1998, 125, 1103–1112. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GEO Project | Samples | Normal | UC | CD | Platform |
|---|---|---|---|---|---|
| GSE13367 | 17 | 9 | 8 | NA | Affymetrix Huma Genome U133 Plus 2.0 Array |
| GSE48958 | 15 | 8 | 7 | NA | Affymetrix Human Gene 1.0 ST Array |
| GSE53306 | 28 | 12 | 16 | NA | Illumina HumanHT-12 WG-DASL V4.0 R2 |
| GSE65114 | 28 | 12 | 16 | NA | Affymetrix Human Gene 2.0 ST Array |
| GSE24287 | 72 | 25 | NA | 47 | Agilent Whole Human Genome Microarray 4x44K G4112F |
| GSE66207 | 33 | 13 | NA | 20 | Illumina HiSeq 2500 |
| GSE100833 | 327 | 168 | NA | 159 | Affymetrix HT HG-U133+ PM Array Plate |
| n | F/M | Average Age | |
|---|---|---|---|
| Crohn’s disease | 27 | 14/13 | 42 ± 19 |
| 13 | 8/5 | 43 ± 19 |
| 14 | 6/8 | 42 ± 19 |
| Ulcerative colitis | |||
| 15 | 9/6 | 49 ± 17 |
| Normal mucosa | 22 | 17/5 | 44 ± 22 |
| 10 | 8/2 | 44 ± 24 |
| 12 | 9/3 | 44 ± 22 |
| Gene | Assay ID | Product (bp) |
|---|---|---|
| B2M | Hs99999907_m1 | 75 |
| IPO8 | Hs00183533_m1 | 71 |
| BMP8B | Hs01629120_s1 | 105 |
| INSL5 | Hs00193884_m1 | 119 |
| LEFTY1 | Hs04235412_sH | 109 |
| Gene | CD | UC | Difference | ||
|---|---|---|---|---|---|
| logFC | p-Value | logFC | p-Value | logFC | |
| INSL5 | −1.26 | 0.015 | 3.22 | <0.001 | 4.48 |
| VIP | −1.23 | 0.003 | 2.77 | <0.001 | 4.00 |
| FAM3B | 2.35 | 0.002 | −0.72 | 0.002 | 3.07 |
| NTS | 2.08 | 0.012 | −0.81 | 0.003 | 2.89 |
| LEFTY1 | −1.38 | 0.043 | 0.92 | 0.001 | 2.30 |
| SEMA6D | 0.83 | 0.005 | −0.77 | 0.013 | 1.60 |
| FGF2 | −0.66 | 0.010 | 0.90 | 0.003 | 1.56 |
| SEMA3A | −0.66 | 0.022 | 0.82 | <0.001 | 1.48 |
| EREG | −0.62 | 0.047 | 0.81 | 0.004 | 1.43 |
| BMP8B | 0.57 | 0.017 | −0.79 | 0.007 | 1.36 |
| Category | Number of Available Categories | UC * | CD * | Union * |
|---|---|---|---|---|
| Gene ontology (GO) | ||||
| Biological processes | 6168 | 1322 | 100 | 1323 |
| Cellular component | 758 | 4 | NA | 4 |
| Molecular functions | 1550 | 41 | 14 | 41 |
| Pathway analysis | ||||
| BioCarta pathways | 310 | 22 | NA | 22 |
| KEGG pathways | 280 | 61 | 6 | 61 |
| Reactome pathways | 1300 | 47 | 2 | 47 |
| Phenotype | ||||
| NIA-Phenotypes | 704 | 129 | NA | 129 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skok, D.J.; Hauptman, N.; Jerala, M.; Zidar, N. Expression of Cytokine-Coding Genes BMP8B, LEFTY1 and INSL5 Could Distinguish between Ulcerative Colitis and Crohn’s Disease. Genes 2021, 12, 1477. https://doi.org/10.3390/genes12101477
Skok DJ, Hauptman N, Jerala M, Zidar N. Expression of Cytokine-Coding Genes BMP8B, LEFTY1 and INSL5 Could Distinguish between Ulcerative Colitis and Crohn’s Disease. Genes. 2021; 12(10):1477. https://doi.org/10.3390/genes12101477
Chicago/Turabian StyleSkok, Daša Jevšinek, Nina Hauptman, Miha Jerala, and Nina Zidar. 2021. "Expression of Cytokine-Coding Genes BMP8B, LEFTY1 and INSL5 Could Distinguish between Ulcerative Colitis and Crohn’s Disease" Genes 12, no. 10: 1477. https://doi.org/10.3390/genes12101477
APA StyleSkok, D. J., Hauptman, N., Jerala, M., & Zidar, N. (2021). Expression of Cytokine-Coding Genes BMP8B, LEFTY1 and INSL5 Could Distinguish between Ulcerative Colitis and Crohn’s Disease. Genes, 12(10), 1477. https://doi.org/10.3390/genes12101477