
Meiosis in Polyploids and Implications for Genetic Mapping: A Review


 ,
,
Abstract
:1. Overview
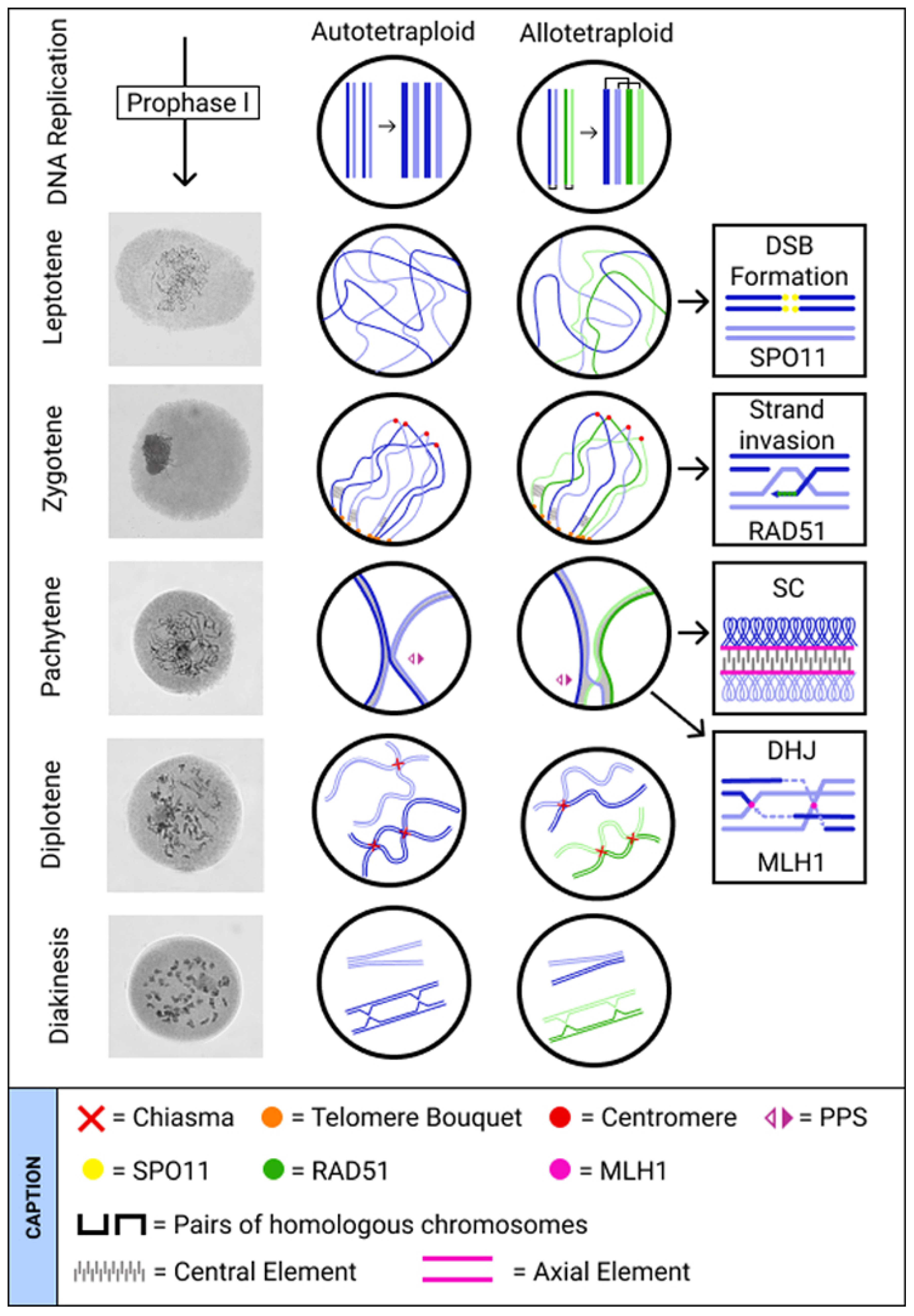
2. Revisiting Early Meiosis
3. Meiosis in Autopolyploids
4. Frequency of Crossovers
5. Multivalents and Cytological Diploidization
6. Genetic Control of Meiosis in Autopolyploids
7. Meiosis in Allopolyploids
8. Homoeologous Exchanges
9. Neoallopolyploids
10. Genetic Regulatory Systems in Allopolyploids
11. Wheat

12. The Ph1 Locus
13. The ZIP4 Gene
14. Ph2
15. Brassica and the Prevalence of Bivalent Pairing
16. PrBn Molecular Characterization and Function
17. BnaPh1
18. Meiotic Proteins and Crossover Formation
19. The Consequences of Meiosis for Genetic Mapping in Auto- and Allopolyploids
- Random chromosome segregation [259], where the gametes are formed by p/2 homologous chromosomes selected from p chromosomes; for example, in an autotetraploid genotype where p = 4, the genotype (A1, A2, A3, A4) can yield six different balanced gametes (A1A2, A1A3, A1A4, A2A3, A2A4, and A3A4) with same expected proportions of 1/6.
- Random chromatid segregation [262], where the gamete is formed by p/2 homologous chromosomes selected at random from 2p possible chromatids due to double reductional segregation; thus, in addition to the heterozygous classes presented in the previous autotetraploid example, four extra homozygotic types are expected (A1A1, A2A2, A3A3, A4A4), with proportions of 1/7 for heterozygous classes and 1/28 for homozygous classes.
20. Case Studies
20.1. Sugarcane
20.2. Sweet Potato
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newton, W.C.F.; Darlington, C.D. Meiosis in Polyploids: Part I. Triploid and Pentaploid Tulips. J. Genet. 1929, 21, 1–15. [Google Scholar] [CrossRef]
- Gillies, C.B. Chromosome Pairing and Fertility in Polyploids; CRC Press: Boca Raton, FL, USA, 1989. [Google Scholar]
- Jenkins, G. Chromosome Pairing and Fertility in Plant Hybrids. In Fertility and Chromosome Pairing: Recent Studies in Plants and Animals; Taylor & Francis: Oxfordshire, UK, 1989; pp. 109–135. ISBN 9781003068433. [Google Scholar]
- Zickler, D.; Kleckner, N. Meiotic Chromosomes: Integrating Structure and Function. Annu. Rev. Genet. 1999, 33, 603–754. [Google Scholar] [CrossRef]
- Cifuentes, M.; Grandont, L.; Moore, G.; Marie, A.; Jenczewski, E.; Chèvre, A.M.; Jenczewski, E. Genetic Regulation of Meiosis in Polyploid Species: New Insights into an Old Question. New Phytol. 2010, 186, 29–36. [Google Scholar] [CrossRef]
- Grandont, L.; Jenczewski, E.; Lloyd, A. Meiosis and Its Deviations in Polyploid Plants. Cytogenet. Genome Res. 2013, 140, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Bomblies, K.; Jones, G.; Franklin, C.; Zickler, D.; Kleckner, N. The Challenge of Evolving Stable Polyploidy: Could an Increase in “Crossover Interference Distance” Play a Central Role? Chromosoma 2016, 125, 287–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bomblies, K.; Higgins, J.D.; Yant, L. Meiosis Evolves: Adaptation to External and Internal Environments. New Phytol. 2015, 208, 306–323. [Google Scholar] [CrossRef]
- Lloyd, A.; Bomblies, K. Meiosis in Autopolyploid and Allopolyploid Arabidopsis. Curr. Opin. Plant Biol. 2016, 30, 116–122. [Google Scholar] [CrossRef]
- Pelé, A.; Rousseau-Gueutin, M.; Chèvre, A.-M.M. Speciation Success of Polyploid Plants Closely Relates to the Regulation of Meiotic Recombination. Front. Plant Sci. 2018, 9, 907. [Google Scholar] [CrossRef]
- Comai, L. The Advantages and Disadvantages of Being Polyploid. Nat. Rev. Genet. 2005, 6, 836–846. [Google Scholar] [CrossRef] [PubMed]
- McCollum, G.D. Comparative Studies of Chromosome Pairing in Natural and Induced Tetraploid Dactylis. Chromosoma 1957, 9, 571–605. [Google Scholar] [CrossRef]
- Yant, L.; Hollister, J.D.; Wright, K.M.; Arnold, B.J.; Higgins, J.D.; Franklin, F.C.H.; Bomblies, K. Meiotic Adaptation to Genome Duplication in Arabidopsis arenosa. Curr. Biol. 2013, 23, 2151–2156. [Google Scholar] [CrossRef] [Green Version]
- Higgins, E.E.; Howell, E.C.; Armstrong, S.J.; Parkin, I.A.P. A Major Quantitative Trait Locus on Chromosome A9, BnaPh1, Controls Homoeologous Recombination in Brassica napus. New Phytol. 2021, 229, 3281–3293. [Google Scholar] [CrossRef]
- Szadkowski, E.; Eber, F.; Huteau, V.; Lodé, M.; Huneau, C.; Belcram, H.; Coriton, O.; Manzanares-Dauleux, M.J.; Delourme, R.; King, G.J.; et al. The First Meiosis of Resynthesized Brassica napus, a Genome Blender. New Phytol. 2010, 186, 102–112. [Google Scholar] [CrossRef]
- Zamariola, L.; Tiang, C.L.; De Storme, N.; Pawlowski, W.; Geelen, D. Chromosome Segregation in Plant Meiosis. Front. Plant Sci. 2014, 5, 279. [Google Scholar] [CrossRef] [Green Version]
- Schubert, I.; Shaw, P. Organization and Dynamics of Plant Interphase Chromosomes. Trends Plant Sci. 2011, 16, 273–281. [Google Scholar] [CrossRef]
- Avivi, L.; Feldman, M. Arrangement of Chromosomes in the Interphase Nucleus of Plants. Hum. Genet. 1980, 55, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Cowan, C.R.; Carlton, P.M.; Cande, W.Z. The Polar Arrangement of Telomeres in Interphase and Meiosis. Rabl Organization and the Bouquet. Plant Physiol. 2001, 125, 532–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idziak, D.; Robaszkiewicz, E.; Hasterok, R. Spatial Distribution of Centromeres and Telomeres at Interphase Varies among Brachypodium Species. J. Exp. Bot. 2015, 66, 6623–6634. [Google Scholar] [CrossRef] [Green Version]
- Fransz, P.; De Jong, J.H.; Lysak, M.; Castiglione, M.R.; Schubert, I. Interphase Chromosomes in Arabidopsis Are Organized as Well Defined Chromocenters from Which Euchromatin Loops Emanate. Proc. Natl. Acad. Sci. USA 2002, 99, 14584–14589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cremer, T.; Cremer, M. Chromosome Territories. Cold Spring Harb. Perspect. Biol. 2010, 2, a003889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, J.R.; Gorkin, D.U.; Ren, B. Chromatin Domains: The Unit of Chromosome Organization. Mol. Cell 2016, 62, 668–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bass, H.W.; Marshall, W.F.; Sedat, J.W.; Agard, D.A.; Cande, W.Z. Telomeres Cluster De Novo before the Initiation of Synapsis: A Three-Dimensional Spatial Analysis of Telomere Positions before and during Meiotic Prophase. J. Cell Biol. 1997, 137, 5–18. [Google Scholar] [CrossRef]
- Tiang, C.L.; He, Y.; Pawlowski, W.P. Chromosome Organization and Dynamics during Interphase, Mitosis, and Meiosis in Plants. Plant Physiol. 2012, 158, 26–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sybenga, J. Chromosome Pairing Affinity and Quadrivalent Formation in Polyploids: Do Segmental Allopolyploids Exist? Genome 1996, 39, 1176–1184. [Google Scholar] [CrossRef]
- Clancy, S. Genetic Recombination. Nat. Educ. 2008, 1, 40. [Google Scholar]
- Zielinski, M.-L.; Mittelsten Scheid, O. Meiosis in Polyploid Plants. In Polyploidy and Genome Evolution; Springer: Berlin/Heidelberg, Germany, 2012; ISBN 9783642314421. [Google Scholar]
- Martinez-Perez, E.; Shaw, P.J.; Moore, G. Polyploidy Induces Centromere Association. J. Cell Biol. 2000, 148, 233–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keeney, S.; Giroux, C.N.; Kleckner, N. Meiosis-Specific DNA Double-Strand Breaks Are Catalyzed by Spo11, a Member of a Widely Conserved Protein Family. Cell 1997, 88, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Neale, M.J.; Keeney, S. Clarifying the Mechanics of DNA Strand Exchange in Meiotic Recombination. Nature 2006, 442, 153–158. [Google Scholar] [CrossRef]
- Hunter, N.; Kleckner, N. The Single-End Invasion: An Asymmetric Intermediate at the Double-Strand Break to Double-Holliday Junction Transition of Meiotic Recombination. Cell 2001, 106, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Lohmiller, L.D.; De Muyt, A.; Howard, B.; Offenberg, H.H.; Heyting, C.; Grelon, M.; Anderson, L.K. Cytological Analysis of MRE11 Protein during Early Meiotic Prophase I in Arabidopsis and Tomato. Chromosoma 2008, 117, 277–288. [Google Scholar] [CrossRef]
- Anderson, L.K.; Stack, S.M. Recombination Nodules in Plants. Cytogenet. Genome Res. 2005, 109, 198–204. [Google Scholar] [CrossRef]
- Roeder, G.S. Meiotic Chromosomes: It Takes Two to Tango. Genes Dev. 1997, 11, 2600–2621. [Google Scholar] [CrossRef] [Green Version]
- Page, S.L.; Hawley, R.S. The Genetics and Molecular Biology of the Synaptonemal Complex. Annu. Rev. Cell Dev. Biol. 2004, 20, 525–558. [Google Scholar] [CrossRef] [PubMed]
- Zickler, D.; Kleckner, N. Recombination, Pairing, and Synapsis of Homologs during Meiosis. Cold Spring Harb. Perspect. Biol. 2015, 7, a016626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, J.E.; Jones, G.H. Meiosis in Autopolyploid Crepis capillaris I. Triploids and Trisomics; Implications for Models of Chromosome Pairing. Chromosoma 1993, 102, 195–206. [Google Scholar] [CrossRef]
- Prieto, P.; Naranjo, T. Analytical Methodology of Meiosis in Autopolyploid and Allopolyploid Plants. In Plant Meiosis; Pradillo, M., Heckmann, S., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2020; Volume 2061, pp. 141–168. ISBN 978-1-4939-9817-3. [Google Scholar]
- Loidl, J. Synaptonemal Complex Spreading in Allium. II. Tetraploid A. vineale. Can. J. Genet. Cytol. 1986, 28, 754–761. [Google Scholar] [CrossRef]
- Loidl, J.; Jones, G.H. Synaptonemal Complex Spreading in Allium. Chromosoma 1986, 93, 420–428. [Google Scholar] [CrossRef]
- John, B.; Henderson, S.A. Asynapsis and Polyploidy in Schistocerca paranensis. Chromosoma 1962, 13, 111–147. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, J.; Schemske, D.W. Neopolyploidy in Flowering Plants. Annu. Rev. Ecol. Syst. 2002, 33, 589–639. [Google Scholar] [CrossRef] [Green Version]
- Pecinka, A.; Fang, W.; Rehmsmeier, M.; Levy, A.A.; Mittelsten Scheid, O. Polyploidization Increases Meiotic Recombination Frequency in Arabidopsis. BMC Biol. 2012, 9, 24, Erratum in 2012, 10, 33. [Google Scholar] [CrossRef] [Green Version]
- Hamant, O.; Ma, H.; Cande, W.Z. Genetics of Meiotic Prophase I in Plants. Annu. Rev. Plant Biol. 2006, 57, 267–302. [Google Scholar] [CrossRef]
- Mercier, R.; Grelon, M. Meiosis in Plants: Ten Years of Gene Discovery. Cytogenet. Genome Res. 2008, 120, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Moran, E.; Osman, K.; Higgins, J.D.; Pradillo, M.; Cuñado, N.; Jones, G.H.; Franklin, F.C.H. ASY1 Coordinates Early Events in the Plant Meiotic Recombination Pathway. Cytogenet. Genome Res. 2008, 120, 302–312. [Google Scholar] [CrossRef] [PubMed]
- De Muyt, A.; Mercier, R.; Mézard, C.; Grelon, M. Meiotic Recombination and Crossovers in Plants. Genome Dyn. 2009, 5, 14–25. [Google Scholar] [CrossRef]
- Edlinger, B.; Schlögelhofer, P. Have a Break: Determinants of Meiotic DNA Double Strand Break (DSB) Formation and Processing in Plants. J. Exp. Bot. 2011, 62, 1545–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, A.; Delgado, M.; Barão, A.; Frescatada, M.; Ribeiro, E.; Pikaard, C.S.; Viegas, W.; Neves, N. Chromosome and DNA Methylation Dynamics during Meiosis in the Autotetraploid Arabidopsis arenosa. Sex. Plant Reprod. 2010, 23, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavania, U.C. Polyploid Breeding: Meiosis in the Diploid Progenitor and Its Predictive Value for Fertility in the Autotetraploid. Proc. Indian Nat. Sci. Acad. B 1991, 57, 17–24. [Google Scholar]
- Lavania, U.C. High Bivalent Frequencies in Artificial Autopolyploids of Hyoscyamus muticus L. Can. J. Genet. Cytol. 1986, 28, 7–11. [Google Scholar] [CrossRef]
- Brubaker, C.L.; Paterson, A.H.; Wendel, J.F. Comparative Genetic Mapping of Allotetraploid Cotton and Its Diploid Progenitors. Genome 1999, 42, 184–203. [Google Scholar] [CrossRef]
- Mézard, C. Meiotic Recombination Hotspots in Plants. Biochem. Soc. Trans. 2006, 34, 531–534. [Google Scholar] [CrossRef]
- Drouaud, J.; Camilleri, C.; Bourguignon, P.Y.; Canaguier, A.; Bérard, A.; Vezon, D.; Giancola, S.; Brunel, D.; Colot, V.; Prum, B.; et al. Variation in Crossing-over Rates across Chromosome 4 of Arabidopsis thaliana Reveals the Presence of Meiotic Recombination “Hot Spots”. Genome Res. 2006, 16, 106–114. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Plagnol, V.; Hu, T.T.; Toomajian, C.; Clark, R.M.; Ossowski, S.; Ecker, J.R.; Weigel, D.; Nordborg, M. Recombination and Linkage Disequilibrium in Arabidopsis thaliana. Nat. Genet. 2007, 39, 1151–1155. [Google Scholar] [CrossRef]
- Santos, J.L.; Alfaro, D.; Sanchez-Moran, E.; Armstrong, S.J.; Franklin, F.C.H.; Jones, G.H. Partial Diploidization of Meiosis in Autotetraploid Arabidopsis thaliana. Genetics 2003, 165, 1533–1540. [Google Scholar] [CrossRef]
- Weiss, H.; Maluszynska, J. Chromosomal Rearrangement in Autotetraploid Plants of Arabidopsis thaliana. Hereditas 2001, 133, 255–261. [Google Scholar] [CrossRef]
- Hollister, J.D.; Arnold, B.J.; Svedin, E.; Xue, K.S.; Dilkes, B.P.; Bomblies, K. Genetic Adaptation Associated with Genome-Doubling in Autotetraploid Arabidopsis arenosa. PLoS Genet. 2012, 8, e1003093. [Google Scholar] [CrossRef] [Green Version]
- Stebbins, G.L. Types of Polyploids: Their Classification and Significance. Adv. Genet. 1947, 1, 403–429. [Google Scholar] [CrossRef]
- Crowley, J.G.; Rees, H. Fertility and Selection in Tetraploid Lolium. Chromosoma 1968, 24, 300–308. [Google Scholar] [CrossRef]
- Abraham, K.; Nemorin, A.; Lebot, V.; Arnau, G. Meiosis and Sexual Fertility of Autotetraploid Clones of Greater Yam Dioscorea alata L. Genet. Resour. Crop Evol. 2013, 60, 819–823. [Google Scholar] [CrossRef]
- Jones, G.H.; Khazanehdari, K.A.; Ford-Lloyd, B.V. Meiosis in the Leek (Allium porrum L.) Revisited. II. Metaphase I Observations. Heredity 1996, 76, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.H.; Datson, P.M.; Manako, K.I.; Murray, B.G. Meiotic Chromosome Pairing Behaviour of Natural Tetraploids and Induced Autotetraploids of Actinidia chinensis. Theor. Appl. Genet. 2014, 127, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Kollmann, F. Allium ampeloprasum—A Polyploid Complex II. Meiosis and Relationships between the Ploidy Types. Caryologia 1972, 25, 295–312. [Google Scholar] [CrossRef]
- Wolf, P.G.; Soltis, P.S.; Soltis, D.E. Tetrasomic Inheritance and Chromosome Pairing Behaviour in the Naturally Occurring Autotetraploid Heuchera grossulariifolia (Saxifragaceae). Genome 1989, 32, 655–659. [Google Scholar] [CrossRef]
- Jannoo, N.; Grivet, L.; David, J.; D’Hont, A.; Glaszmann, J.C. Differential Chromosome Pairing Affinities at Meiosis in Polyploid Sugarcane Revealed by Molecular Markers. Heredity 2004, 93, 460–467. [Google Scholar] [CrossRef]
- Parisod, C.; Alix, K.; Just, J.; Petit, M.; Sarilar, V.; Mhiri, C.; Ainouche, M.; Chalhoub, B.; Grandbastien, M.A. Impact of Transposable Elements on the Organization and Function of Allopolyploid Genomes. New Phytol. 2010, 186, 37–45. [Google Scholar] [CrossRef]
- Lam, W.S.; Yang, X.; Makaroff, C.A. Characterization of Arabidopsis thaliana SMC1 and SMC3: Evidence That AtSMC3 May Function beyond Chromosome Cohesion. J. Cell Sci. 2005, 118, 3037–3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradillo, M.; Knoll, A.; Oliver, C.; Varas, J.; Corredor, E.; Puchta, H.; Santos, J.L. Involvement of the Cohesin Cofactor PDS5 (SPO76) During Meiosis and DNA Repair in Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 1034. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, A.M.; Lister, C.; Page, T.; Fransz, P.; Findlay, K.; Jones, G.H.; Dickinson, H.G.; Dean, C. The DIF1 Gene of Arabidopsis Is Required for Meiotic Chromosome Segregation and Belongs to the REC8/RAD21 Cohesin Gene Family. Plant J. 1999, 19, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Dong, F.; Edelmann, R.E.; Makaroff, C.A. The Arabidopsis SYN1 Cohesin Protein Is Required for Sister Chromatid Arm Cohesion and Homologous Chromosome Pairing. J. Cell Sci. 2003, 116, 2999–3007. [Google Scholar] [CrossRef] [Green Version]
- Chelysheva, L.; Diallo, S.; Vezon, D.; Gendrot, G.; Vrielynck, N.; Belcram, K.; Rocques, N.; Márquez-Lema, A.; Bhatt, A.M.; Horlow, C.; et al. AtREC8 and AtSCC3 Are Essential to the Monopolar Orientation of the Kinetochores during Meiosis. J. Cell Sci. 2005, 118, 4621–4632. [Google Scholar] [CrossRef] [Green Version]
- Lambing, C.; Tock, A.J.; Topp, S.D.; Choi, K.; Kuo, P.C.; Zhao, X.; Osman, K.; Higgins, J.D.; Franklin, F.C.H.; Henderson, I.R. Interacting Genomic Landscapes of REC8-Cohesin, Chromatin, and Meiotic Recombination in Arabidopsis. Plant Cell 2020, 32, 1218–1239. [Google Scholar] [CrossRef] [Green Version]
- Kleckner, N. Chiasma Formation: Chromatin/Axis Interplay and the Role(s) of the Synaptonemal Complex. Chromosoma 2006, 115, 175–194. [Google Scholar] [CrossRef]
- Armstrong, S.J.; Caryl, A.P.; Jones, G.H.; Franklin, F.C.H. Asy1, a Protein Required for Meiotic Chromosome Synapsis, Localizes to Axis-Associated Chromatin in Arabidopsis and Brassica. J. Cell Sci. 2002, 115, 3645–3655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferdous, M.; Higgins, J.D.; Osman, K.; Lambing, C.; Roitinger, E.; Mechtler, K.; Armstrong, S.J.; Perry, R.; Pradillo, M.; Cuñado, N.; et al. Inter-Homolog Crossing-Over and Synapsis in Arabidopsis Meiosis Are Dependent on the Chromosome Axis Protein AtASY3. PLoS Genet. 2012, 8, e1002507. [Google Scholar] [CrossRef] [Green Version]
- Chambon, A.; West, A.; Vezon, D.; Horlow, C.; De Muyt, A.; Chelysheva, L.; Ronceret, A.; Darbyshire, A.; Osman, K.; Heckmann, S.; et al. Identification of ASYNAPTIC4, a Component of the Meiotic Chromosome Axis. Plant Physiol. 2018, 178, 233–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caryl, A.P.; Armstrong, S.J.; Jones, G.H.; Franklin, F.C.H. A Homologue of the Yeast HOP1 Gene Is Inactivated in the Arabidopsis Meiotic Mutant Asy1. Chromosoma 2000, 109, 62–71. [Google Scholar] [CrossRef] [PubMed]
- West, A.M.; Rosenberg, S.C.; Ur, S.N.; Lehmer, M.K.; Ye, Q.; Hagemann, G.; Caballero, I.; Usón, I.; MacQueen, A.J.; Herzog, F.; et al. A Conserved Filamentous Assembly Underlies the Structure of the Meiotic Chromosome Axis. eLife 2019, 8, e40372. [Google Scholar] [CrossRef]
- Morgan, C.; Zhang, H.; Henry, C.E.; Franklin, F.C.H.; Bomblies, K. Derived Alleles of Two Axis Proteins Affect Meiotic Traits in Autotetraploid Arabidopsis arenosa. Proc. Natl. Acad. Sci. USA 2020, 117, 8980–8988. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Moran, E.; Santos, J.L.; Jones, G.H.; Franklin, F.C.H. ASY1 Mediates AtDMC1-Dependent Interhomolog Recombination during Meiosis in Arabidopsis. Genes Dev. 2007, 21, 2220–2233. [Google Scholar] [CrossRef] [Green Version]
- Moran, E.S.; Armstrong, S.J.; Santos, J.L.; Franklin, F.C.H.; Jones, G.H. Chiasma Formation in Arabidopsis thaliana Accession Wassileskija and in Two Meiotic Mutants. Chromosome Res. 2001, 9, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.; Zhao, X.; Tock, A.J.; Lambing, C.; Underwood, C.J.; Hardcastle, T.J.; Serra, H.; Kim, J.; Cho, H.S.; Kim, J.; et al. Nucleosomes and DNA Methylation Shape Meiotic DSB Frequency in Arabidopsis thaliana Transposons and Gene Regulatory Regions. Genome Res. 2018, 28, 532–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Underwood, C.J.; Choi, K.; Lambing, C.; Zhao, X.; Serra, H.; Borges, F.; Simorowski, J.; Ernst, E.; Jacob, Y.; Henderson, I.R.; et al. Epigenetic Activation of Meiotic Recombination near Arabidopsis thaliana Centromeres via Loss of H3K9me2 and Non-CG DNA Methylation. Genome Res. 2018, 28, 519–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Wang, M.; Dukowic-Schulze, S.; Zhou, A.; Tiang, C.-L.; Shilo, S.; Sidhu, G.K.; Eichten, S.; Bradbury, P.; Springer, N.M.; et al. Genomic Features Shaping the Landscape of Meiotic Double-Strand-Break Hotspots in Maize. Proc. Natl. Acad. Sci. USA 2017, 114, 12231–12236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, S.J.; Franklin, F.C.H.; Jones, G.H. Nucleolus-Associated Telomere Clustering and Pairing Precede Meiotic Chromosome Synapsis in Arabidopsis thaliana. J. Cell Sci. 2001, 114, 4207–4217. [Google Scholar] [CrossRef] [PubMed]
- Lambing, C.; Kuo, P.C.; Tock, A.J.; Topp, S.D.; Henderson, I.R. ASY1 Acts as a Dosage-Dependent Antagonist of Telomere-Led Recombination and Mediates Crossover Interference in Arabidopsis. Proc. Natl. Acad. Sci. USA 2020, 117, 13647–13658. [Google Scholar] [CrossRef] [PubMed]
- Yelina, N.E.; Lambing, C.; Hardcastle, T.J.; Zhao, X.; Santos, B.; Henderson, I.R. DNA Methylation Epigenetically Silences Crossover Hot Spots and Controls Chromosomal Domains of Meiotic Recombination in Arabidopsis. Genes Dev. 2015, 29, 2183–2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seear, P.J.; France, M.G.; Gregory, C.L.; Heavens, D.; Schmickl, R.; Yant, L.; Higgins, J.D. A Novel Allele of ASY3 Is Associated with Greater Meiotic Stability in Autotetraploid Arabidopsis lyrata. PLoS Genet. 2020, 16, e1008900. [Google Scholar] [CrossRef]
- Hazarika, M.H.; Rees, H. Genotypic Control of Chromosome Behaviour in Rye X. Chromosome Pairing and Fertility in Autotetraploids. Heredity 1967, 22, 317–332. [Google Scholar] [CrossRef] [Green Version]
- Marburger, S.; Monnahan, P.; Seear, P.J.; Martin, S.H.; Koch, J.; Paajanen, P.; Bohutínská, M.; Higgins, J.D.; Schmickl, R.; Yant, L. Interspecific Introgression Mediates Adaptation to Whole Genome Duplication. Nat. Commun. 2019, 10, 5218. [Google Scholar] [CrossRef] [Green Version]
- Jenczewski, E.; Alix, K. From Diploids to Allopolyploids: The Emergence of Efficient Pairing Control Genes in Plants. Crit. Rev. Plant Sci. 2004, 23, 21–45. [Google Scholar] [CrossRef]
- Sears, E.R. Genetic Control of Chromosome Pairing in Wheat. Annu. Rev. Genet. 1976, 10, 31–51. [Google Scholar] [CrossRef]
- Grandont, L.; Cuñado, N.; Coriton, O.; Huteau, V.; Eber, F.; Chèvre, A.M.; Grelon, M.; Chelysheva, L.; Jenczewski, E. Homoeologous Chromosome Sorting and Progression of Meiotic Recombination in Brassica napus: Ploidy Does Matter! Plant Cell 2014, 26, 1448–1463. [Google Scholar] [CrossRef] [Green Version]
- Mason, A.S.; Wendel, J.F. Homoeologous Exchanges, Segmental Allopolyploidy, and Polyploid Genome Evolution. Front. Genet. 2020, 11, 1014. [Google Scholar] [CrossRef] [PubMed]
- Svačina, R.; Sourdille, P.; Kopecký, D.; Bartoš, J. Chromosome Pairing in Polyploid Grasses. Front. Plant Sci. 2020, 11, 1056. [Google Scholar] [CrossRef]
- Cuñado, N.; Callejas, S.; García, M.J.; Fernández, A.; Santos, J.L. The Pattern of Zygotene and Pachytene Pairing in Allotetraploid Aegilops Species Sharing the U Genome. Theor. Appl. Genet. 1996, 93, 1152–1155. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, G.; Rees, H. Strategies of Bivalent Formation in Allopolyploid Plants. Proc. R. Soc. B Biol. Sci. 1991, 243, 209–214. [Google Scholar] [CrossRef]
- Bennett, M.D. Centromere Arrangements in Triticum aestivum and Their Relationship to Synapsis. Heredity 1979, 43, 157. [Google Scholar]
- Moore, G. Meiosis in Allopolyploids—The Importance of ‘Teflon’ Chromosomes. Trends Genet. 2002, 18, 456–463. [Google Scholar] [CrossRef]
- Trelles-Sticken, E.; Loidl, J.; Scherthan, H. Bouquet Formation in Budding Yeast: Initiation of Recombination Is Not Required for Meiotic Telomere Clustering. J. Cell Sci. 1999, 112, 651–658. [Google Scholar] [CrossRef]
- Chikashige, Y.; Ding, D.Q.; Imai, Y.; Yamamoto, M.; Haraguchi, T.; Hiraoka, Y. Meiotic Nuclear Reorganization: Switching the Position of Centromeres and Telomeres in the Fission Yeast Schizosaccharomyces pombe. EMBO J. 1997, 16, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Niwa, O.; Shimanuki, M.; Miki, F. Telomere-Led Bouquet Formation Facilitates Homologous Chromosome Pairing and Restricts Ectopic Interaction in Fission Yeast Meiosis. EMBO J. 2000, 19, 3831–3840. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Perez, E.; Shaw, P.; Reader, S.; Aragon-Alcaide, L.; Miller, T.; Moore, G. Homologous Chromosome Pairing in Wheat. J. Cell Sci. 1999, 112, 1761–1769. [Google Scholar] [CrossRef] [PubMed]
- Mikhailova, E.I.; Sosnikhina, S.P.; Kirillova, G.A.; Tikholiz, O.A.; Smirnov, V.G.; Jones, R.N.; Jenkins, G. Nuclear Dispositions of Subtelomeric and Pericentromeric Chromosomal Domains during Meiosis in Asynaptic Mutants of Rye (Secale cereale L.). J. Cell Sci. 2001, 114, 1875–1882. [Google Scholar] [CrossRef] [PubMed]
- Gillies, C.B. An Electron Microscopic Study of Synaptonemal Complex Formation at Zygotene in Rye. Chromosoma 1985, 92, 165–175. [Google Scholar] [CrossRef]
- Gillies, C.B. Synaptonemal Complex and Chromosome Structure. Annu. Rev. Genet. 1975, 9, 91–109. [Google Scholar] [CrossRef]
- Bass, H.W.; Riera-Lizarazu, O.; Ananiev, E.V.; Bordoli, S.J.; Rines, H.W.; Phillips, R.L.; Sedat, J.W.; Agard, D.A.; Cande, W.Z. Evidence for the Coincident Initiation of Homolog Pairing and Synapsis during the Telomere-Clustering (Bouquet) Stage of Meiotic Prophase. J. Cell Sci. 2000, 113, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Loidl, J.; Ehrendorfer, F.; Schweizer, D. EM Analysis of Meiotic Chromosome Pairing in a Pentaploid Achillea Hybrid. Heredity 1990, 65, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Sybenga, J. Meiotic Configurations; Springer: Berlin, Germany, 1975; ISBN 3540073477. [Google Scholar]
- Grant, V. Genetics of Flowering Plants; Columbia University Press: New York, NY, USA, 1978. [Google Scholar]
- Hobolth, P. Chromosome Pairing in Allohexaploid Wheat Var. Chinese Spring. Transformation of Multivalents into Bivalents, a Mechanism for Exclusive Bivalent Formation. Carlsberg Res. Commun. 1981, 46, 129–173. [Google Scholar] [CrossRef] [Green Version]
- Holm, P.B. Chromosome Pairing and Chiasma Formation in Allohexaploid Wheat, Triticum aestivum Analyzed by Spreading of Meiotic Nuclei. Carlsberg Res. Commun. 1986, 51, 239–294. [Google Scholar] [CrossRef] [Green Version]
- Desai, A.; Chee, P.W.; Rong, J.; May, O.L.; Paterson, A.H. Chromosome Structural Changes in Diploid and Tetraploid A Genomes of Gossypium. Genome 2006, 49, 336–345. [Google Scholar] [CrossRef]
- Bingham, E.T.; Burnham, C.R.; Gates, C.E. Double and Single Backcross Linkage Estimates in Autotetraploid Maize. Genetics 1968, 59, 399. [Google Scholar] [CrossRef] [PubMed]
- Leflon, M.; Grandont, L.; Eber, F.; Huteau, V.; Coriton, O.; Chelysheva, L.; Jenczewski, E.; Chèvre, A.-M. Crossovers Get a Boost in Brassica Allotriploid and Allotetraploid Hybrids. Plant Cell 2010, 22, 2253–2264. [Google Scholar] [CrossRef] [Green Version]
- Pelé, A.; Falque, M.; Trotoux, G.; Eber, F.; Nègre, S.; Gilet, M.; Huteau, V.; Lodé, M.; Jousseaume, T.; Dechaumet, S.; et al. Amplifying Recombination Genome-Wide and Reshaping Crossover Landscapes in Brassicas. PLoS Genet. 2017, 13, e1006794. [Google Scholar] [CrossRef]
- Jenczewski, E.; Chèvre, A.M.; Alix, K. Chromosomal and Gene Expression Changes in Brassica Allopolyploids. In Polyploid and Hybrid Genomics; Wiley: New York, NY, USA, 2013; pp. 171–186. [Google Scholar] [CrossRef]
- Suay, L.; Zhang, D.; Eber, F.; Jouy, H.; Lodé, M.; Huteau, V.; Coriton, O.; Szadkowski, E.; Leflon, M.; Martin, O.C.; et al. Crossover Rate between Homologous Chromosomes and Interference Are Regulated by the Addition of Specific Unpaired Chromosomes in Brassica. New Phytol. 2014, 201, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Comai, L.; Tyagi, A.P.; Lysak, M.A. FISH Analysis of Meiosis in Arabidopsis Allopolyploids. Chromosome Res. 2003, 11, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Gaeta, R.T.; Chris Pires, J. Homoeologous Recombination in Allopolyploids: The Polyploid Ratchet. New Phytol. 2010, 186, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.; Gaeta, R.T.; Pires, J.C. Homoeologous Shuffling and Chromosome Compensation Maintain Genome Balance in Resynthesized Allopolyploid Brassica napus. Proc. Natl. Acad. Sci. USA 2011, 108, 7908–7913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkin, I.A.P.; Sharpe, A.G.; Lydiate, D.J. Patterns of Genome Duplication within the Brassica napus Genome. Genome 2003, 46, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Schuermann, D.; Molinier, J.; Fritsch, O.; Hohn, B. The Dual Nature of Homologous Recombination in Plants. Trends Genet. 2005, 21, 172–181. [Google Scholar] [CrossRef]
- Leitch, A.R.; Leitch, I.J. Genomic Plasticity and the Diversity of Polyploid Plants. Science 2008, 320, 481–484. [Google Scholar] [CrossRef]
- Stebbins, G.L. Chromosomal Evolution in Higher Plants; Edward Arnold Ltd.: London, UK, 1971; ISBN 9780713122879. [Google Scholar]
- Otto, S.P.; Whitton, J. Polyploid Incidence and Evolution. Annu. Rev. Genet. 2000, 34, 401–437. [Google Scholar] [CrossRef] [Green Version]
- Schiessl, S.; Katche, E.; Ihien, E.; Chawla, H.S.; Mason, A.S. The Role of Genomic Structural Variation in the Genetic Improvement of Polyploid Crops. Crop J. 2019, 7, 127–140. [Google Scholar] [CrossRef]
- Gaeta, R.T.; Pires, J.C.; Iniguez-Luy, F.; Leon, E.; Osborn, T.C. Genomic Changes in Resynthesized Brassica napus and Their Effect on Gene Expression and Phenotype. Plant Cell 2007, 19, 3403–3417. [Google Scholar] [CrossRef] [Green Version]
- Song, K.; Lu, P.; Tang, K.; Osborn, T.C. Rapid Genome Change in Synthetic Polyploids of Brassica and Its Implications for Polyploid Evolution. Proc. Natl. Acad. Sci. USA 1995, 92, 7719–7723. [Google Scholar] [CrossRef] [Green Version]
- Henry, I.M.; Dilkes, B.P.; Tyagi, A.; Gao, J.; Christensen, B.; Comai, L. The BOY NAMED SUE Quantitative Trait Locus Confers Increased Meiotic Stability to an Adapted Natural Allopolyploid of Arabidopsis. Plant Cell 2014, 26, 181–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oleszczuk, S.; Lukaszewski, A.J. The Origin of Unusual Chromosome Constitutions among Newly Formed Allopolyploids. Am. J. Bot. 2014, 101, 318–326. [Google Scholar] [CrossRef] [Green Version]
- Ramanna, M.S.; Jacobsen, E. Relevance of Sexual Polyploidization for Crop Improvement—A Review. Euphytica 2003, 133, 3–8. [Google Scholar] [CrossRef]
- Chester, M.; Gallagher, J.P.; Symonds, V.V.; Cruz da Silva, A.V.; Mavrodiev, E.V.; Leitch, A.R.; Soltis, P.S.; Soltis, D.E. Extensive Chromosomal Variation in a Recently Formed Natural Allopolyploid Species, Tragopogon miscellus (Asteraceae). Proc. Natl. Acad. Sci. USA 2012, 109, 1176–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.Q.; Yen, Y.; Zheng, Y.L.; Liu, D.C. Meiotic Restriction in Emmer Wheat Is Controlled by One or More Nuclear Genes That Continue to Function in Derived Lines. Sex. Plant Reprod. 2007, 20, 159–166. [Google Scholar] [CrossRef]
- Silkova, O.G.; Adonina, I.G.; Krivosheina, E.A.; Shchapova, A.I.; Shumny, V.K. Chromosome Pairing in Meiosis of Partially Fertile Wheat/Rye Hybrids. Plant Reprod. 2013, 26, 33–41. [Google Scholar] [CrossRef]
- Zhang, H.; Bian, Y.; Gou, X.; Zhu, B.; Xu, C.; Qi, B.; Li, N.; Rustgi, S.; Zhou, H.; Han, F.; et al. Persistent Whole-Chromosome Aneuploidy Is Generally Associated with Nascent Allohexaploid Wheat. Proc. Natl. Acad. Sci. USA 2013, 110, 3447–3452. [Google Scholar] [CrossRef] [Green Version]
- Tate, J.A.; Joshi, P.; Soltis, K.A.; Soltis, P.S.; Soltis, D.E. On the Road to Diploidization? Homoeolog Loss in Independently Formed Populations of the Allopolyploid Tragopogon miscellus (Asteraceae). BMC Plant Biol. 2009, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buggs, R.J.A.; Chamala, S.; Wu, W.; Tate, J.A.; Schnable, P.S.; Soltis, D.E.; Soltis, P.S.; Barbazuk, W.B. Rapid, Repeated, and Clustered Loss of Duplicate Genes in Allopolyploid Plant Populations of Independent Origin. Curr. Biol. 2012, 22, 248–252. [Google Scholar] [CrossRef] [Green Version]
- Martín, A.C.; Rey, M.D.; Shaw, P.; Moore, G.; Moore, G. Dual Effect of the Wheat Ph1 Locus on Chromosome Synapsis and Crossover. Chromosoma 2017, 126, 669–680. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, F.M.; McGinnis, R.C. The Meiotic Behavior of a Nulli-Haploid Plant in Avena sativa L. Can. J. Genet. Cytol. 1968, 10, 186–189. [Google Scholar] [CrossRef]
- Jauhar, P.P. Genetic Control of Diploid-like Meiosis in Hexaploid Tall Fescue. Nature 1975, 254, 595–597. [Google Scholar] [CrossRef]
- Jenczewski, E.; Eber, F.; Grimaud, A.; Huet, S.; Lucas, M.O.; Monod, H.; Chèvre, A.M. PrBn, a Major Gene Controlling Homeologous Pairing in Oilseed Rape (Brassica napus) Haploids. Genetics 2003, 164, 645–653. [Google Scholar] [CrossRef]
- Nonomura, K.I.; Nakano, M.; Eiguchi, M.; Suzuki, T.; Kurata, N. PAIR2 Is Essential for Homologous Chromosome Synapsis in Rice Meiosis I. J. Cell Sci. 2006, 119, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Evans, G.M.; Davies, E.W. The Genetics of Meiotic Chromosome Pairing in Lolium temulentum × Lolium perenne Tetraploids. Theor. Appl. Genet. 1985, 71, 185–192. [Google Scholar] [CrossRef]
- Armstead, I.P.; Bollard, A.; King, I.P.; Forster, J.W.; Hayward, M.D.; Evans, G.M.; Thomas, H.M. Chromosome Pairing in Lolium perenne × L. temulentum Diploid Hybrids: Genetic and Cytogenetic Evaluation. Heredity 1999, 83, 298–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, H. Sid: A Mendelian Locus Controlling Thylakoid Membrane Disassembly in Senescing Leaves of Festuca pratensis. Theor. Appl. Genet. 1987, 73, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Holm, P.B.; Wang, X. The Effect of Chromosome 5B on Synapsis and Chiasma Formation in Wheat, Triticum aestivum Cv. Chinese Spring. Carlsberg Res. Commun. 1988, 53, 191–208. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, G. Synaptonemal Complex Formation in Hybrids of Lolium temulentum × Lolium perenne (L.)—II. Triploid. Chromosoma 1985, 92, 387–339. [Google Scholar] [CrossRef]
- Waines, J.G. A Model for the Origin of Diploidizing Mechanisms in Polyploid Species. Am. Nat. 1976, 110, 415–430. [Google Scholar] [CrossRef]
- Grusz, A.L.; Sigel, E.M.; Witherup, C. Homoeologous Chromosome Pairing across the Eukaryote Phylogeny. Mol. Phylogenet. Evol. 2017, 117, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, Y. Evolution of Polyploid Triticum Wheats under Cultivation: The Role of Domestication, Natural Hybridization and Allopolyploid Speciation in Their Diversification. Plant Cell Physiol. 2011, 52, 750–764. [Google Scholar] [CrossRef] [Green Version]
- Riley, R.; Chapman, V. Genetic Control of the Cytologically Diploid Behaviour of Hexaploid Wheat. Nature 1958, 182, 713–715. [Google Scholar] [CrossRef]
- Sears, E.R.; Okamoto, M. Intergenomic Chromosome Pairing in Hexploid Wheat. Proc. Int. Congr. Genet. 1958, 2, 258–259. [Google Scholar]
- Prieto, P.; Shaw, P.; Moore, G. Homologue Recognition during Meiosis Is Associated with a Change in Chromatin Conformation. Nat. Cell Biol. 2004, 6, 906–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhaliwal, H.S.; Gill, B.S.; Waines, J.G. Analysis of Induced Homoeologous Pairing in a Ph Mutant Wheat × Rye Hybrid. J. Hered. 1977, 68, 207–209. [Google Scholar] [CrossRef]
- Martín, A.C.; Shaw, P.; Phillips, D.; Reader, S.; Moore, G.; Martı, A.C. Licensing MLH1 Sites for Crossover during Meiosis. Nat. Commun. 2014, 5, 4580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, M.A.; Reader, S.M.; Dalgliesh, C.; Miller, T.E.; Foote, T.N.; Fish, L.J.; Snape, J.W.; Moore, G. Induction and Characterization of Ph1 Wheat Mutants. Genetics 1999, 153, 1909–1918. [Google Scholar] [CrossRef]
- Al-Kaff, N.; Knight, E.; Bertin, I.; Foote, T.; Hart, N.; Griffiths, S.; Moore, G. Detailed Dissection of the Chromosomal Region Containing the Ph1 Locus in Wheat Triticum aestivum: With Deletion Mutants and Expression Profiling. Ann. Bot. 2008, 101, 863–872. [Google Scholar] [CrossRef] [Green Version]
- King, J.; Grewal, S.; Yang, C.Y.; Hubbart, S.; Scholefield, D.; Ashling, S.; Edwards, K.J.; Allen, A.M.; Burridge, A.; Bloor, C.; et al. A Step Change in the Transfer of Interspecific Variation into Wheat from Amblyopyrum muticum. Plant Biotechnol. J. 2017, 15, 217–226. [Google Scholar] [CrossRef]
- Griffiths, S.; Sharp, R.; Foote, T.N.; Bertin, I.; Wanous, M.; Reader, S.; Colas, I.; Moore, G. Molecular Characterization of Ph1 as a Major Chromosome Pairing Locus in Polyploid Wheat. Nature 2006, 439, 749–752. [Google Scholar] [CrossRef]
- Bhullar, R.; Nagarajan, R.; Bennypaul, H.; Sidhu, G.K.; Sidhu, G.; Rustgi, S.; von Wettstein, D.; Gill, K.S. Silencing of a Metaphase I-Specific Gene Results in a Phenotype Similar to That of the Pairing Homeologous 1 (Ph1) Gene Mutations. Proc. Natl. Acad. Sci. USA 2014, 111, 14187–14192. [Google Scholar] [CrossRef] [Green Version]
- Alabdullah, A.K.; Borrill, P.; Martin, A.C.; Ramirez-Gonzalez, R.H.; Hassani-Pak, K.; Uauy, C.; Shaw, P.; Moore, G. A Co-Expression Network in Hexaploid Wheat Reveals Mostly Balanced Expression and Lack of Significant Gene Loss of Homeologous Meiotic Genes upon Polyploidization. Front. Plant Sci. 2019, 10, 1325. [Google Scholar] [CrossRef]
- Martín, A.C.; Borrill, P.; Higgins, J.; Alabdullah, A.; Ramírez-González, R.H.; Swarbreck, D.; Uauy, C.; Shaw, P.; Moore, G. Genome-Wide Transcription during Early Wheat Meiosis Is Independent of Synapsis, Ploidy Level, and the Ph1 Locus. Front. Plant Sci. 2018, 9, 1791. [Google Scholar] [CrossRef]
- Rey, M.D.; Martín, A.C.; Higgins, J.; Swarbreck, D.; Uauy, C.; Shaw, P.; Moore, G. Exploiting the ZIP4 Homologue within the Wheat Ph1 Locus Has Identified Two Lines Exhibiting Homoeologous Crossover in Wheat-Wild Relative Hybrids. Mol. Breed. 2017, 37. [Google Scholar] [CrossRef] [PubMed]
- Krasinska, L.; Besnard, E.; Cot, E.; Dohet, C.; Méchali, M.; Lemaitre, J.-M.; Fisher, D. Cdk1 and Cdk2 Activity Levels Determine the Efficiency of Replication Origin Firing in Xenopus. EMBO J. 2008, 27, 758–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viera, A.; Rufas, J.S.; Martínez, I.; Barbero, J.L.; Ortega, S.; Suja, J.A. CDK2 Is Required for Proper Homologous Pairing, Recombination and Sex-Body Formation during Male Mouse Meiosis. J. Cell Sci. 2009, 122, 2149–2159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marston, A.L.; Amon, A. Meiosis: Cell-Cycle Controls Shuffle and Deal. Nat. Rev. Mol. Cell Biol. 2004, 5, 983–997. [Google Scholar] [CrossRef]
- Cohen, P.E.; Pollack, S.E.; Pollard, J.W. Genetic Analysis of Chromosome Pairing, Recombination, and Cell Cycle Control during First Meiotic Prophase in Mammals. Endocr. Rev. 2006, 27, 398–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight, E.; Greer, E.; Draeger, T.; Thole, V.; Reader, S.; Shaw, P.; Moore, G. Inducing Chromosome Pairing through Premature Condensation: Analysis of Wheat Interspecific Hybrids. Funct. Integr. Genom. 2010, 10, 603–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandrow, M.G.; Hamlin, J.L. Chromatin Decondensation in S-Phase Involves Recruitment of Cdk2 by Cdc45 and Histone H1 Phosphorylation. J. Cell Biol. 2005, 168, 875–886. [Google Scholar] [CrossRef]
- Greer, E.; Martı, A.C.; Moore, G.; Shaw, P. The Ph1 Locus Suppresses Cdk2-Type Activity during Premeiosis and Meiosis in Wheat. Plant Cell 2012, 24, 152–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, T.; Nibau, C.; Phillips, D.W.; Jenkins, G.; Armstrong, S.J.; Doonan, J.H. CDKG1 Protein Kinase Is Essential for Synapsis and Male Meiosis at High Ambient Temperature in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2014, 111, 2182–2187. [Google Scholar] [CrossRef] [Green Version]
- Mézard, C.; Vignard, J.; Drouaud, J.; Mercier, R. The Road to Crossovers: Plants Have Their Say. Trends Genet. 2007, 23, 91–99. [Google Scholar] [CrossRef]
- Lhuissier, F.G.P.; Offenberg, H.H.; Wittich, P.E.; Vischer, N.O.E.; Heyting, C. The Mismatch Repair Protein MLH1 Marks a Subset of Strongly Interfering Crossovers in Tomato. Plant Cell 2007, 19, 862–876. [Google Scholar] [CrossRef] [Green Version]
- Ashley, T.; Walpita, D.; De Rooij, D.G. Localization of Two Mammalian Cyclin Dependent Kinases during Mammalian Meiosis. J. Cell Sci. 2001, 114, 685–693. [Google Scholar] [CrossRef]
- Shen, Y.; Tang, D.; Wang, K.; Wang, M.; Huang, J.; Luo, W.; Luo, Q.; Hong, L.; Li, M.; Cheng, Z. ZIP4 in Homologous Chromosome Synapsis and Crossover Formation in Rice Meiosis. J. Cell Sci. 2012, 125, 2581–2591. [Google Scholar] [CrossRef] [Green Version]
- Chelysheva, L.; Gendrot, G.; Vezon, D.; Doutriaux, M.P.; Mercier, R.; Grelon, M. Zip4/Spo22 Is Required for Class I CO Formation but Not for Synapsis Completion in Arabidopsis thaliana. PLoS Genet. 2007, 3, 802–813. [Google Scholar] [CrossRef]
- Blanc, G.; Wolfe, K.H. Functional Divergence of Duplicated Genes Formed by Polyploidy during Arabidopsis Evolution. Plant Cell 2004, 16, 1679–1691. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, A.H.; Ranoux, M.; Vautrin, S.; Glover, N.; Fourment, J.; Charif, D.; Choulet, F.; Lassalle, G.; Marande, W.; Tran, J.; et al. Meiotic Gene Evolution: Can You Teach a New Dog New Tricks? Mol. Biol. Evol. 2014, 31, 1724–1727. [Google Scholar] [CrossRef] [Green Version]
- Sidhu, G.K.; Warzecha, T.; Pawlowski, W.P. Evolution of Meiotic Recombination Genes in Maize and Teosinte. BMC Genom. 2017, 18, 106. [Google Scholar] [CrossRef] [Green Version]
- Gonzalo, A.; Lucas, M.O.; Charpentier, C.; Sandmann, G.; Lloyd, A.; Jenczewski, E. Reducing MSH4 Copy Number Prevents Meiotic Crossovers between Non-Homologous Chromosomes in Brassica napus. Nat. Commun. 2019, 10, 2354. [Google Scholar] [CrossRef]
- de Muyt, A.; Pyatnitskaya, A.; Andréani, J.; Ranjha, L.; Ramus, C.; Laureau, R.; Fernandez-Vega, A.; Holoch, D.; Girard, E.; Govin, J.; et al. A Meiotic XPF–ERCC1-like Complex Recognizes Joint Molecule Recombination Intermediates to Promote Crossover Formation. Genes Dev. 2018, 32, 283–296. [Google Scholar] [CrossRef] [Green Version]
- Rey, M.; Martín, A.C.; Smedley, M.; Hayta, S.; Harwood, W.; Shaw, P.; Moore, G. Magnesium Increases Homoeologous Crossover Frequency During Meiosis in ZIP4 (Ph1 Gene) Mutant Wheat-Wild Relative Hybrids. Front. Plant Sci. 2018, 9, 509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alabdullah, A.K.; Moore, G.; Martín, A.C. A Duplicated Copy of the Meiotic Gene ZIP4 Preserves up to 50% Pollen Viability and Grain Number in Polyploid Wheat. Biology 2021, 10, 290. [Google Scholar] [CrossRef] [PubMed]
- Colas, I.; Shaw, P.; Prieto, P.; Wanous, M.; Spielmeyer, W.; Mago, R.; Moore, G. Effective Chromosome Pairing Requires Chromatin Remodeling at the Onset of Meiosis. Proc. Natl. Acad. Sci. USA 2008, 105, 6075–6080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, D.-Q.; Matsuda, A.; Okamasa, K.; Nagahama, Y.; Haraguchi, T.; Hiraoka, Y. Meiotic Cohesin-Based Chromosome Structure Is Essential for Homologous Chromosome Pairing in Schizosaccharomyces pombe. Chromosoma 2016, 125, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golubovskaya, I.N.; Hamant, O.; Timofejeva, L.; Wang, C.-J.R.; Braun, D.; Meeley, R.; Cande, W.Z. Alleles of Afd1 Dissect REC8 Functions during Meiotic Prophase I. J. Cell Sci. 2006, 119, 3306–3315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jauhar, P.P. Inter- and Intragenomic Chromosome Pairing in Haploids of Durum Wheat. J. Hered. 1999, 90, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Mello-Sampayo, T. Homoeologous Chromosome Pairing in Pentaploid Hybrids of Wheat. In Proceedings of the Third International Wheat Genetics Symposium, Canberra, Australia, 5–9 August 1968; pp. 179–184. [Google Scholar]
- Mello-Sampayo, T. Genetic Regulation of Meiotic Chromosome Pairing by Chromosome 3D of Triticum aestivum. Nat. New Biol. 1971, 230, 22–23. [Google Scholar] [CrossRef]
- Sears, E.R. A Wheat Mutation Conditioning an Intermediate Level of Homoeologous Chromosome Pairing. Can. J. Genet. Cytol. 1982, 24, 715–719. [Google Scholar] [CrossRef]
- Wall, A.M.; Riley, R.; Chapman, V. Wheat Mutants Permitting Homoeologous Meiotic Chromosome Pairing. Genet. Res. 1971, 18, 311–328. [Google Scholar] [CrossRef] [Green Version]
- Sutton, T.; Whitford, R.; Baumann, U.; Dong, C.; Able, J.A.; Langridge, P. The Ph2 Pairing Homoeologous Locus of Wheat (Triticum aestivum): Identification of Candidate Meiotic Genes Using a Comparative Genetics Approach. Plant J. 2003, 36, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.-H.; Langridge, P. An Early Meiosis cDNA Clone from Wheat. Mol. Gen. Genet. 1994, 243, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Whitford, R. From Intimate Chromosome Associations to Wild Sex in Wheat (Triticum aestivum); University of Adelaide: Adelaide, Australia, 2002. [Google Scholar]
- Letarte, J. Identification and Characterisation of Early Meiotic Genes in Wheat; University of Adelaide: Adelaide, Australia, 1996. [Google Scholar]
- Dong, C.; Thomas, S.; Becker, D.; Lörz, H.; Whitford, R.; Sutton, T.; Able, J.A.; Langridge, P. WM5: Isolation and Characterisation of a Gene Expressed during Early Meiosis and Shoot Meristem Development in Wheat. Funct. Plant Biol. 2005, 32, 249. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Whitford, R.; Langridge, P. A DNA Mismatch Repair Gene Links to the Ph2 Locus in Wheat. Genome 2002, 45, 116–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, A.H.; Milligan, A.S.; Langridge, P.; Able, J.A. TaMSH7: A Cereal Mismatch Repair Gene That Affects Fertility in Transgenic Barley (Hordeum vulgare L.). BMC Plant Biol. 2007, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Serra, H.; Svačina, R.; Baumann, U.; Whitford, R.; Sutton, T.; Bartoš, J.; Sourdille, P. Ph2 Encodes the Mismatch Repair Protein MSH7-3D That Inhibits Wheat Homoeologous Recombination. Nat. Commun. 2021, 12, 803. [Google Scholar] [CrossRef]
- Benavente, E.; Orellana, J.; Fernández-Calvín, B. Comparative Analysis of the Meiotic Effects of Wheat Ph1b and Ph2b Mutations in Wheat×rye Hybrids. Theor. Appl. Genet. 1998, 96, 1200–1204. [Google Scholar] [CrossRef]
- Martinez, M.; Cuñado, N.; Carcelén, N.; Romero, C. The Ph1 and Ph2 Loci Play Different Roles in the Synaptic Behaviour of Hexaploid Wheat Triticum aestivum. Theor. Appl. Genet. 2001, 103, 398–405. [Google Scholar] [CrossRef]
- Sánchez-Morán, E.; Benavente, E.; Orellana, J. Analysis of Karyotypic Stability of Homoeologous-Pairing (Ph) Mutants in Allopolyploid Wheats. Chromosoma 2001, 110, 371–377. [Google Scholar] [CrossRef]
- Sears, E.R. An Induced Mutant with Homoeologous Pairing in Common Wheat. Can. J. Genet. Cytol. 1977, 19, 585–593. [Google Scholar] [CrossRef]
- Prieto, P.; Moore, G.; Reader, S. Control of Conformation Changes Associated with Homologue Recognition during Meiosis. Theor. Appl. Genet. 2005, 111, 505–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boden, S.A.; Langridge, P.; Spangenberg, G.; Able, J.A. TaASY1 Promotes Homologous Chromosome Interactions and Is Affected by Deletion of Ph1. Plant J. 2009, 57, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Reyes, G.X.; Schmidt, T.T.; Kolodner, R.D.; Hombauer, H. New Insights into the Mechanism of DNA Mismatch Repair. Chromosoma 2015, 124, 443–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, U.; Alani, E. Understanding How Mismatch Repair Proteins Participate in the Repair/Anti-Recombination Decision. FEMS Yeast Res. 2016, 16, fow071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceoloni, C.; Donini, P. Combining Mutations for the Two Homoeologous Pairing Suppressor Genes Ph1 and Ph2 in Common Wheat and in Hybrids with alien Triticeae. Genome 1993, 36, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Lysak, M.A.; Koch, M.A.; Pecinka, A.; Schubert, I. Chromosome Triplication Found across the Tribe Brassiceae. Genome Res. 2005, 15, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Nagaharu, U. Genome Analysis in Brassica with Special Reference to the Experimental Formation of B. napus and Peculiar Mode of Fertilization. Jpn. J. Bot. 1935, 7, 389–452. [Google Scholar]
- Inaba, R.; Nishio, T. Phylogenetic Analysis of Brassiceae Based on the Nucleotide Sequences of the S-Locus Related Gene, SLR1. Theor. Appl. Genet. 2002, 105, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Inomata, N. Hybrid Progenies of the Cross, Brassica campestris x B. oleracea. I. Cytogenetical Studies on F1 Hybrids. Jpn. J. Genet. 1980, 55, 189–202. [Google Scholar] [CrossRef] [Green Version]
- Attia, T.; Röbbelen, G. Meiotic Pairing in Haploids and Amphidiploids of Spontaneous versus Synthetic Origin in Rape, Brassica napus L. Can. J. Genet. Cytol. 1986, 28, 330–334. [Google Scholar] [CrossRef]
- Parkin, I.A.P.; Gulden, S.M.; Sharpe, A.G.; Lukens, L.; Trick, M.; Osborn, T.C.; Lydiate, D.J. Segmental Structure of the Brassica napus Genome Based on Comparative Analysis with Arabidopsis thaliana. Genetics 2005, 171, 765–781. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.J.; Kim, J.S.; Kwon, S.J.; Lim, K.B.; Choi, B.S.; Kim, J.A.; Jin, M.; Park, J.Y.; Lim, M.H.; Kim, H.I.; et al. Sequence-Level Analysis of the Diploidization Process in the Triplicated Flowering Locus C Region of Brassica rapa. Plant Cell 2006, 18, 1339–1347. [Google Scholar] [CrossRef] [Green Version]
- Parkin, I.A.; Sharpe, A.G.; Keith, D.J.; Lydiate, D.J. Identification of the A and C Genomes of amphidiploid Brassica napus (Oilseed rape). Genome 1995, 38, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Udall, J.A.; Quijada, P.A.; Osborn, T.C. Detection of Chromosomal Rearrangements Derived from Homeologous Recombination in Four Mapping Populations of Brassica napus L. Genetics 2005, 169, 967–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, K.C.; Keller, W.A. Chromosome Pairing in Haploids of Brassica oleracea. Theor. Appl. Genet. 1982, 24, 49–52. [Google Scholar] [CrossRef]
- Prakash, S. Taxonomy, Cytogenetics and Origin of Crop Brassica, a Review; Swedish Natural Science Research Council: Stockholm, Sweden, 1980; Volume 55, pp. 1–57. [Google Scholar]
- Schmidt, R.; Acarkan, A.; Boivin, K. Comparative Structural Genomics in the Brassicaceae Family. Plant Physiol. Biochem. 2001, 39, 253–262. [Google Scholar] [CrossRef]
- Liu, Z.; Adamczyk, K.; Manzanares-Dauleux, M.; Eber, F.; Lucas, M.O.; Delourme, R.; Chèvre, A.M.; Jenczewski, E. Mapping PrBn and Other Quantitative Trait Loci Responsible for the Control of Homeologous Chromosome Pairing in Oilseed Rape (Brassica napus L.) Haploids. Genetics 2006, 174, 1583–1596. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, S.D.; Leflon, M.; Monod, H.; Eber, F.; Coriton, O.; Huteau, V.; Chevre, A.-M.; Jenczewski, E. Genetic Regulation of Meiotic Cross-Overs between Related Genomes in Brassica napus Haploids and Hybrids. Plant Cell 2009, 21, 373–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cifuentes, M.; Eber, F.; Lucas, M.O.; Lode, M.; Chevre, A.M.; Jenczewskia, E. Repeated Polyploidy Drove Different Levels of Crossover Suppression between Homoeologous Chromosomes in Brassica napus Allohaploids. Plant Cell 2010, 22, 2265–2276. [Google Scholar] [CrossRef] [Green Version]
- Cui, C.; Ge, X.; Gautam, M.; Kang, L.; Li, Z. Cytoplasmic and Genomic Effects on Meiotic Pairing in Brassica Hybrids and Allotetraploids from Pair Crosses of Three Cultivated Diploids. Genetics 2012, 191, 725–738. [Google Scholar] [CrossRef] [Green Version]
- Mason, A.S.; Huteau, V.; Eber, F.; Coriton, O.; Yan, G.; Nelson, M.N.; Cowling, W.A.; Chèvre, A.M. Genome Structure Affects the Rate of Autosyndesis and Allosyndesis in AABC, BBAC and CCAB Brassica Interspecific Hybrids. Chromosome Res. 2010, 18, 655–666. [Google Scholar] [CrossRef]
- Mason, A.S.; Nelson, M.N.; Yan, G.; Cowling, W.A. Production of Viable Male Unreduced Gametes in Brassica Interspecific Hybrids Is Genotype Specific and Stimulated by Cold Temperatures. BMC Plant Biol. 2011, 11, 103. [Google Scholar] [CrossRef] [Green Version]
- Carlton, P.M.; Farruggio, A.P.; Dernburg, A.F. A Link between Meiotic Prophase Progression and Crossover Control. PLoS Genet. 2006, 2, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Perez, E.; Moore, G. To Check or Not to Check? The Application of Meiotic Studies to Plant Breeding. Curr. Opin. Plant Biol. 2008, 11, 222–227. [Google Scholar] [CrossRef]
- Gaebelein, R.; Mason, A.S. Allohexaploids in the Genus Brassica. Crit. Rev. Plant Sci. 2018, 37, 422–437. [Google Scholar] [CrossRef]
- Prakash, S.; Takahata, Y.; Kirti, P.B.; Chopra, V.L. Cytogenetics. In Biology of Brassica coenospecies; Gomez-Campo, C., Ed.; Elsevier Science: Amsterdam, The Netherlands, 1999; pp. 59–106. ISBN 9780080528021. [Google Scholar]
- Tian, E.; Jiang, Y.; Chen, L.; Zou, J.; Liu, F.; Meng, J. Synthesis of a Brassica Trigenomic Allohexaploid (B. carinata × B. rapa) de Novo and Its Stability in Subsequent Generations. Theor. Appl. Genet. 2010, 121, 1431–1440. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Tan, C.; Cui, C.; Ge, X.; Li, Z. Distinct Subgenome Stabilities in Synthesized Brassica Allohexaploids. Theor. Appl. Genet. 2016, 129, 1257–1271. [Google Scholar] [CrossRef] [PubMed]
- Mwathi, M.W.; Gupta, M.; Atri, C.; Banga, S.S.; Batley, J.; Mason, A.S. Segregation for Fertility and Meiotic Stability in Novel Brassica Allohexaploids. Theor. Appl. Genet. 2017, 130, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Gaebelein, R.; Schiessl, S.V.; Samans, B.; Batley, J.; Mason, A.S. Inherited Allelic Variants and Novel Karyotype Changes Influence Fertility and Genome Stability in Brassica Allohexaploids. New Phytol. 2019, 223, 965–978. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.P.; Zhao, M.; Ma, J.; Yu, J.; Huang, S.; et al. The Brassica oleracea Genome Reveals the Asymmetrical Evolution of Polyploid Genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef] [PubMed]
- Parkin, I.A.P.; Koh, C.; Tang, H.; Robinson, S.J.; Kagale, S.; Clarke, W.E.; Town, C.D.; Nixon, J.; Krishnakumar, V.; Bidwell, S.L.; et al. Transcriptome and Methylome Profiling Reveals Relics of Genome Dominance in the Mesopolyploid Brassica oleracea. Genome Biol. 2014, 15, R77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodhouse, M.R.; Cheng, F.; Pires, J.C.; Lisch, D.; Freeling, M.; Wang, X. Origin, Inheritance, and Gene Regulatory Consequences of Genome Dominance in Polyploids. Proc. Natl. Acad. Sci. USA 2014, 111, 6527–6527. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Sun, R.; Hou, X.; Zheng, H.; Zhang, F.; Zhang, Y.; Liu, B.; Liang, J.; Zhuang, M.; Liu, Y.; et al. Subgenome Parallel Selection Is Associated with Morphotype Diversification and Convergent Crop Domestication in Brassica rapa and Brassica oleracea. Nat. Genet. 2016, 48, 1218–1224. [Google Scholar] [CrossRef]
- Song, Q.; Chen, J.Z. Epigenetic and Developmental Regulation in Plant Polyploids. Curr. Opin. Plant Biol. 2015, 24, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Schiessl, S.; Huettel, B.; Kuehn, D.; Reinhardt, R.; Snowdon, R. Post-Polyploidisation Morphotype Diversification Associates with Gene Copy Number Variation. Sci. Rep. 2017, 7, 41845. [Google Scholar] [CrossRef]
- Clarke, W.E.; Higgins, E.E.; Plieske, J.; Wieseke, R.; Sidebottom, C.; Khedikar, Y.; Batley, J.; Edwards, D.; Meng, J.; Li, R.; et al. A High-Density SNP Genotyping Array for Brassica napus and Its Ancestral Diploid Species Based on Optimised Selection of Single-Locus Markers in the Allotetraploid Genome. Theor. Appl. Genet. 2016, 129, 1887–1899. [Google Scholar] [CrossRef] [Green Version]
- Aklilu, B.B.; Soderquist, R.S.; Culligan, K.M. Genetic Analysis of the Replication Protein A Large Subunit Family in Arabidopsis Reveals Unique and Overlapping Roles in DNA Repair, Meiosis and DNA Replication. Nucleic Acids Res. 2014, 42, 3104–3118. [Google Scholar] [CrossRef] [Green Version]
- Berchowitz, L.E.; Francis, K.E.; Bey, A.L.; Copenhaver, G.P. The Role of AtMUS81 in Interference-Insensitive Crossovers in A. Thaliana. PLoS Genet. 2007, 3, 1355–1364. [Google Scholar] [CrossRef]
- Higgins, J.D.; Buckling, E.F.; Franklin, F.C.H.; Jones, G.H. Expression and Functional Analysis of AtMUS81 in Arabidopsis Meiosis Reveals a Role in the Second Pathway of Crossing-Over. Plant J. 2008, 54, 152–162. [Google Scholar] [CrossRef]
- Blary, A. Towards a Functional Characterization of Meiotic Recombination in Rapeseed: Analysis of the Meiotic Transcriptome and Hyper-Recombinant Mutants; Université Paris-Saclay: Saint-Aubin, France, 2016. [Google Scholar]
- Kunkel, T.A.; Erie, D.A. DNA Mismatch Repair. Annu. Rev. Biochem. 2005, 74, 681–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, J.D.; Armstrong, S.J.; Franklin, F.C.H.; Jones, G.H. The Arabidopsis MutS Homolog AtMSH4 Functions at an Early Step in Recombination: Evidence for Two Classes of Recombination in Arabidopsis. Genes Dev. 2004, 18, 2557–2570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, J.D.; Sanchez-Moran, E.; Armstrong, S.J.; Jones, G.H.; Franklin, F.C.H. The Arabidopsis Synaptonemal Complex Protein ZYP1 Is Required for Chromosome Synapsis and Normal Fidelity of Crossing Over. Genes Dev. 2005, 19, 2488–2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culligan, K.M.; Hays, J.B. Arabidopsis MutS Homologs—AtMSH2, AtMSH3, AtMSH6, and a Novel AtMSH7—Form Three Distinct Protein Heterodimers with Different Specificities for Mismatched DNA. Plant Cell 2000, 12, 991–1002. [Google Scholar] [CrossRef] [Green Version]
- Sourdille, P.; Jenczewski, E. Homoeologous Exchanges in Allopolyploids: How Brassica napus Established Self-control! New Phytol. 2021, 229, 3041–3043. [Google Scholar] [CrossRef]
- Dubois, E.; De Muyt, A.; Soyer, J.L.; Budin, K.; Legras, M.; Piolot, T.; Debuchy, R.; Kleckner, N.; Zickler, D.; Espagne, E. Building Bridges to Move Recombination Complexes. Proc. Natl. Acad. Sci. USA 2019, 116, 12400–12409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelysheva, L.; Vezon, D.; Chambon, A.; Gendrot, G.; Pereira, L.; Lemhemdi, A.; Vrielynck, N.; Le Guin, S.; Novatchkova, M.; Grelon, M. The Arabidopsis HEI10 Is a New ZMM Protein Related to Zip3. PLoS Genet. 2012, 8, e1002799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Wang, M.; Tang, D.; Shen, Y.; Miao, C.; Hu, Q.; Lu, T.; Cheng, Z. The Role of Rice HEI10 in the Formation of Meiotic Crossovers. PLoS Genet. 2012, 8, e1002809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziolkowski, P.A.; Underwood, C.J.; Lambing, C.; Martinez-Garcia, M.; Lawrence, E.J.; Ziolkowska, L.; Griffin, C.; Choi, K.; Franklin, F.C.H.; Martienssen, R.A.; et al. Natural Variation and Dosage of the HEI10 Meiotic E3 Ligase Control Arabidopsis Crossover Recombination. Genes Dev. 2017, 31, 306–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cifuentes, M.; Rivard, M.; Pereira, L.; Chelysheva, L.; Mercier, R. Haploid Meiosis in Arabidopsis: Double-Strand Breaks Are Formed and Repaired but Without Synapsis and Crossovers. PLoS ONE 2013, 8, e72431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, H.J. A New Mode of Segregation in Gregory’s Tetraploid Primulas. Am. Nat. 1914, 48, 508–512. [Google Scholar] [CrossRef]
- Gregory, R.P. On the Genetics of Tetraploid Plants in Primula sinensis. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1914, 87, 484–492. [Google Scholar] [CrossRef]
- Lawrence, W.J.C. The Genetics and Cytology of Dahlia Species. J. Genet. 1929, 21, 125–159. [Google Scholar] [CrossRef]
- Haldane, J.B.S. Theoretical Genetics of Autopolyploids. J. Genet. 1930, 22, 359–372. [Google Scholar] [CrossRef]
- De Winton, D.; Haldane, J.B.S. Linkage in the Tetraploid Primula sinensis. J. Genet. 1931, 24, 121–144. [Google Scholar] [CrossRef]
- Mather, K. Reductional and Equational Separation of the Chromosomes in Bivalents and Multivalents. J. Genet. 1935, 30, 53–78. [Google Scholar] [CrossRef]
- Mather, K. Segregation and Linkage in Autotetraploids. J. Genet. 1936, 32, 287–314. [Google Scholar] [CrossRef]
- Fisher, R.A.; Mather, K. Polyploid Inheritance in Lythrum salicaria. Nature 1942, 150, 430–430. [Google Scholar] [CrossRef]
- Fisher, B.A.; Mather, K. Non-Lethality of the Mid Factor in Lythrum salicaria. Nature 1940, 146, 521–521. [Google Scholar] [CrossRef]
- Fisher, R.A.; Mather, K. The Inheritance of Style Length in Lythrum salicaria. Ann. Eugen. 1943, 12, 1–23. [Google Scholar] [CrossRef]
- Dawson, C.D.R. Tetrasomic Inheritance in Lotus corniculatus L. J. Genet. 1941, 42, 49–72. [Google Scholar] [CrossRef]
- Fisher, R.A. The Theoretical Consequences of Polyploid Inheritance for The Mid Style Form of Lythrum salicaria. Ann. Eugen. 1941, 11, 31–38. [Google Scholar] [CrossRef]
- Fisher, R.A. The Theory of Linkage in Polysomic Inheritance. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1947, 233, 55–87. [Google Scholar] [CrossRef]
- Fisher, R.A. Statistical Methods for Research Workers; Oliver and Boyd Ltd.: London, UK, 1954. [Google Scholar]
- Wu, K.K.; Burnquist, W.; Sorrells, M.E.; Tew, T.L.; Moore, P.H.; Tanksley, S.D. The Detection and Estimation of Linkage in Polyploids Using Single-Dose Restriction Fragments. Theor. Appl. Genet. 1992, 83, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Sorrells, M.E. Development and Application of RFLPs in Polyploids. Crop Sci. 1992, 32, 1086–1091. [Google Scholar] [CrossRef]
- Doerge, R.W.; Craig, B.A. Model Selection for Quantitative Trait Locus Analysis in Polyploids. Proc. Natl. Acad. Sci. USA 2000, 97, 7951–7956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackett, C.A.; McLean, K.; Bryan, G.J. Linkage Analysis and QTL Mapping Using SNP Dosage Data in a Tetraploid Potato Mapping Population. PLoS ONE 2013, 8, e63939. [Google Scholar] [CrossRef] [Green Version]
- Bourke, P.M.; Voorrips, R.E.; Visser, R.G.F.; Maliepaard, C. The Double-Reduction Landscape in Tetraploid Potato as Revealed by a High-Density Linkage Map. Genetics 2015, 201, 853–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Geest, G.; Bourke, P.M.; Voorrips, R.E.; Marasek-Ciolakowska, A.; Liao, Y.; Post, A.; van Meeteren, U.; Visser, R.G.F.; Maliepaard, C.; Arens, P. An Ultra-Dense Integrated Linkage Map for Hexaploid Chrysanthemum Enables Multi-Allelic QTL Analysis. Theor. Appl. Genet. 2017, 130, 2527–2541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollinari, M.; Olukolu, B.A.; Da Pereira, G.S.; Khan, A.; Gemenet, D.; Craig Yencho, G.; Zeng, Z.B. Unraveling the Hexaploid Sweetpotato Inheritance Using Ultra-Dense Multilocus Mapping. G3 Genes Genomes Genet. 2020, 10, 281–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappai, F.; Amadeu, R.R.; Benevenuto, J.; Cullen, R.; Garcia, A.; Grossman, A.; Ferrão, L.F.V.; Munoz, P. High-Resolution Linkage Map and QTL Analyses of Fruit Firmness in Autotetraploid Blueberry. Front. Plant Sci. 2020, 11, 1767. [Google Scholar] [CrossRef] [PubMed]
- Voorrips, R.E.; Gort, G.; Vosman, B. Genotype Calling in Tetraploid Species from Bi-Allelic Marker Data Using Mixture Models. BMC Bioinform. 2011, 12, 172. [Google Scholar] [CrossRef] [Green Version]
- Serang, O.; Mollinari, M.; Garcia, A.A.F. Efficient Exact Maximum a Posteriori Computation for Bayesian SNP Genotyping in Polyploids. PLoS ONE 2012, 7, e30906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerard, D.; Ferrão, L.F.V.; Garcia, A.A.F.; Stephens, M. Genotyping Polyploids from Messy Sequencing Data. Genetics 2018, 210, 789–807. [Google Scholar] [CrossRef] [Green Version]
- Clark, L.V.; Lipka, A.E.; Sacks, E.J. PolyRAD: Genotype Calling with Uncertainty from Sequencing Data in Polyploids and Diploids. G3 Genes Genomes Genet. 2019, 9, 663–673. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Van Eck, H.J.; Rouppe Van Der Voort, J.N.A.M.; Huigen, D.J.; Stam, P.; Jacobsen, E. Autotetraploids and Genetic Mapping Using Common AFLP Markers: The R2 Allele Conferring Resistance to Phytophthora infestans Mapped on Potato Chromosome 4. Theor. Appl. Genet. 1998, 96, 1121–1128. [Google Scholar] [CrossRef]
- Al-Janabi, S.M.; Honeycutt, R.J.; McClelland, M.; Sobral, B.W.S. A Genetic Linkage Map of Saccharum spontaneum L. “SES 208”. Genetics 1993, 134, 1249–1260. [Google Scholar] [CrossRef]
- Ming, R.; Liu, S.C.; Bowers, J.E.; Moore, P.H.; Irvine, J.E.; Paterson, A.H. Construction of a Saccharum Consensus Genetic Map from Two Interspecific Crosses. Crop Sci. 2002, 42, 570–583. [Google Scholar] [CrossRef]
- Aitken, K.S.; Jackson, P.A.; McIntyre, C.L. A Combination of AFLP and SSR Markers Provides Extensive Map Coverage and Identification of Homo(Eo)Logous Linkage Groups in a Sugarcane Cultivar. Theor. Appl. Genet. 2005, 110, 789–801. [Google Scholar] [CrossRef]
- Aitken, K.S.; McNeil, M.D.; Hermann, S.; Bundock, P.C.; Kilian, A.; Heller-Uszynska, K.; Henry, R.J.; Li, J. A Comprehensive Genetic Map of Sugarcane That Provides Enhanced Map Coverage and Integrates High-Throughput Diversity Array Technology (DArT) Markers. BMC Genom. 2014, 15, 152. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.A.F.; Kido, E.A.; Meza, A.N.; Souza, H.M.B.; Pinto, L.R.; Pastina, M.M.; Leite, C.S.; da Silva, J.A.G.; Ulian, E.C.; Figueira, A.; et al. Development of an Integrated Genetic Map of a Sugarcane (Saccharum spp.) Commercial Cross, Based on a Maximum-Likelihood Approach for Estimation of Linkage and Linkage Phases. Theor. Appl. Genet. 2006, 112, 298–314. [Google Scholar] [CrossRef]
- Shirasawa, K.; Tanaka, M.; Takahata, Y.; Ma, D.; Cao, Q.; Liu, Q.; Zhai, H.; Kwak, S.S.; Cheol Jeong, J.; Yoon, U.H.; et al. A High-Density SNP Genetic Map Consisting of a Complete Set of Homologous Groups in Autohexaploid Sweetpotato (Ipomoea batatas). Sci. Rep. 2017, 7, 44207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, E.; Shirasawa, K.; Kimura, T.; Monden, Y.; Tanaka, M.; Isobe, S. Genetic Mapping in Autohexaploid Sweet Potato with Low-Coverage NGS-Based Genotyping Data. G3 Genes Genomes Genet. 2020, 10, 2661–2670. [Google Scholar]
- Tennessen, J.A.; Govindarajulu, R.; Ashman, T.L.; Liston, A. Evolutionary Origins and Dynamics of Octoploid Strawberry Subgenomes Revealed by Dense Targeted Capture Linkage Maps. Genome Biol. Evol. 2014, 6, 3295–3313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.R.; Natarajan, S.; Kim, H.T.; Jesse, D.M.I.; Lee, C.G.; Park, J.I.; Nou, I.S. High Density Linkage Map Construction and QTL Mapping for Runner Production in Allo-Octoploid Strawberry Fragaria × Ananassa Based on DdRAD-Seq Derived SNPs. Sci. Rep. 2019, 9, 3275. [Google Scholar] [CrossRef]
- Rajapakse, S.; Byrne, D.H.; Zhang, L.; Anderson, N.; Arumuganathan, K.; Ballard, R.E. Two Genetic Linkage Maps of Tetraploid Roses. Theor. Appl. Genet. 2001, 103, 575–583. [Google Scholar] [CrossRef]
- Bourke, P.M.; Voorrips, R.E.; Visser, R.G.F.; Maliepaard, C. Tools for Genetic Studies in Experimental Populations of Polyploids. Front. Plant Sci. 2018, 9, 513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Hont, A.; Ison, D.; Alix, K.; Roux, C.; Glaszmann, J.C. Determination of Basic Chromosome Numbers in the Genus Saccharum by Physical Mapping of Ribosomal RNA Genes. Genome 1998, 41, 221–225. [Google Scholar] [CrossRef]
- Grivet, L.; Glaszmann, J.C.; Vincentz, M.; Da Silva, F.; Arruda, P. ESTs as a Source for Sequence Polymorphism Discovery in Sugarcane: Example of the Adh Genes. Theor. Appl. Genet. 2003, 106, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Grivet, L.; Arruda, P. Sugarcane Genomics: Depicting the Complex Genome of an Important Tropical Crop. Curr. Opin. Plant Biol. 2002, 5, 122–127. [Google Scholar] [CrossRef]
- Garcia, A.A.F.; Mollinari, M.; Marconi, T.G.; Serang, O.R.; Silva, R.R.; Vieira, M.L.C.; Vicentini, R.; Costa, E.A.; Mancini, M.C.; Garcia, M.O.S.; et al. SNP Genotyping Allows an In-Depth Characterisation of the Genome of Sugarcane and Other Complex Autopolyploids. Sci. Rep. 2013, 3, 3399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimarães, C.T.; Honeycutt, R.J.; Sills, G.R.; Sobral, B.W.S. Genetic Maps of Saccharum officinarum L. and Saccharum robustum Brandes & Jew. Ex Grassl. Genet. Mol. Biol. 1999, 22, 125–132. [Google Scholar] [CrossRef]
- Balsalobre, T.W.A.; da Silva Pereira, G.; Margarido, G.R.A.; Gazaffi, R.; Barreto, F.Z.; Anoni, C.O.; Cardoso-Silva, C.B.; Costa, E.A.; Mancini, M.C.; Hoffmann, H.P.; et al. GBS-Based Single Dosage Markers for Linkage and QTL Mapping Allow Gene Mining for Yield-Related Traits in Sugarcane. BMC Genom. 2017, 18, 72. [Google Scholar] [CrossRef] [Green Version]
- You, Q.; Yang, X.; Peng, Z.; Islam, M.S.; Sood, S.; Luo, Z.; Comstock, J.; Xu, L.; Wang, J. Development of an Axiom Sugarcane100K SNP Array for Genetic Map Construction and QTL Identification. Theor. Appl. Genet. 2019, 132, 2829–2845. [Google Scholar] [CrossRef] [PubMed]
- Loebenstein, G. Origin, Distribution and Economic Importance. In The Sweetpotato; Loebenstein, G., Thottappilly, G., Eds.; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Yang, J.; Moeinzadeh, M.H.; Kuhl, H.; Helmuth, J.; Xiao, P.; Haas, S.; Liu, G.; Zheng, J.; Sun, Z.; Fan, W.; et al. Haplotype-Resolved Sweet Potato Genome Traces Back Its Hexaploidization History. Nat. Plants 2017, 3, 696–703. [Google Scholar] [CrossRef]
- Gustafsson, Å.; Gadd, I. Mutations and Crop Improvement. Iii. Ipomoea batatas (L.) Poir. (Convolvulaceae). Hereditas 1965, 53, 77–89. [Google Scholar] [CrossRef]
- Magoon, M.L.; Krishnan, R.; Vijaya Bai, K. Cytological Evidence on the Origin of Sweet Potato. Theor. Appl. Genet. 1970, 40, 360–366. [Google Scholar] [CrossRef]
- Ukoskit, K.; Thompson, P.G. Autopolyploidy versus Allopolyploidy and Low-Density Randomly Amplified Polymorphic DNA Linkage Maps of Sweetpotato. J. Am. Soc. Hortic. Sci. 1997, 122, 822–828. [Google Scholar] [CrossRef] [Green Version]
- Kriegner, A.; Cervantes, J.C.; Burg, K.; Mwanga, R.O.M.; Zhang, D. A Genetic Linkage Map of Sweetpotato [Ipomoea batatas (L.) Lam.] Based on AFLP Markers. Mol. Breed. 2003, 11, 169–185. [Google Scholar] [CrossRef]
- Cervantes-Flores, J.C.; Yencho, G.C.; Kriegner, A.; Pecota, K.V.; Faulk, M.A.; Mwanga, R.O.M.; Sosinski, B.R. Development of a Genetic Linkage Map and Identification of Homologous Linkage Groups in Sweetpotato Using Multiple-Dose AFLP Markers. Mol. Breed. 2008, 21, 511–532. [Google Scholar] [CrossRef]
- Monden, Y.; Tahara, M. Genetic Linkage Analysis Using DNA Markers in Sweetpotato. Breed. Sci. 2017, 67, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollinari, M.; Garcia, A.A.F. Linkage Analysis and Haplotype Phasing in Experimental Autopolyploid Populations with High Ploidy Level Using Hidden Markov Models. G3 Genes Genomes Genet. 2019, 9, 3297–3314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, G.S.; Mollinari, M.; Schumann, M.J.; Clough, M.E.; Zeng, Z.-B.; Yencho, G.C. The Recombination Landscape and Multiple QTL Mapping in a Solanum tuberosum cv. ‘Atlantic’-Derived F1 Population. Heredity 2021, 126, 817–830. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Ploidy Level | Number of Gametes | Number of Genotypes |
|---|---|---|
| 2 | 2 | 4 |
| 4 | 6 | 36 |
| 6 | 20 | 400 |
| 8 | 70 | 4900 |
| 10 | 252 | 63,504 |
| 12 | 924 | 853,776 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, N.R.; Mollinari, M.; Oliveira, G.K.; Pereira, G.S.; Vieira, M.L.C. Meiosis in Polyploids and Implications for Genetic Mapping: A Review. Genes 2021, 12, 1517. https://doi.org/10.3390/genes12101517
Soares NR, Mollinari M, Oliveira GK, Pereira GS, Vieira MLC. Meiosis in Polyploids and Implications for Genetic Mapping: A Review. Genes. 2021; 12(10):1517. https://doi.org/10.3390/genes12101517
Chicago/Turabian StyleSoares, Nina Reis, Marcelo Mollinari, Gleicy K. Oliveira, Guilherme S. Pereira, and Maria Lucia Carneiro Vieira. 2021. "Meiosis in Polyploids and Implications for Genetic Mapping: A Review" Genes 12, no. 10: 1517. https://doi.org/10.3390/genes12101517