
Fractionated Seminal Plasma of Boar Ejaculates Analyzed by LC–MS/MS: Its Effects on Post-Thaw Semen Quality

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Media
2.2. Animal and Semen Collections
2.3. Experiment I
2.3.1. Sample Preparation and Digestion for Nano LC–MS/MS Analysis
2.3.2. Protein Identification and Quantification
2.3.3. Gene Ontology (GO) Analysis, KEGG, and REAC Pathways
2.4. Experiment II: Semen Processing Procedure and Quality Assessment
2.4.1. Semen Cryopreservation
2.4.2. Post-Thaw Semen Treatment
2.4.3. Semen Quality Analysis
Motility Parameters Analyzed by the Computer-Assisted Sperm Analysis (CASA) System
Membrane Integrity Assessment
Viability Assessment
2.5. Statistical Analysis
3. Results
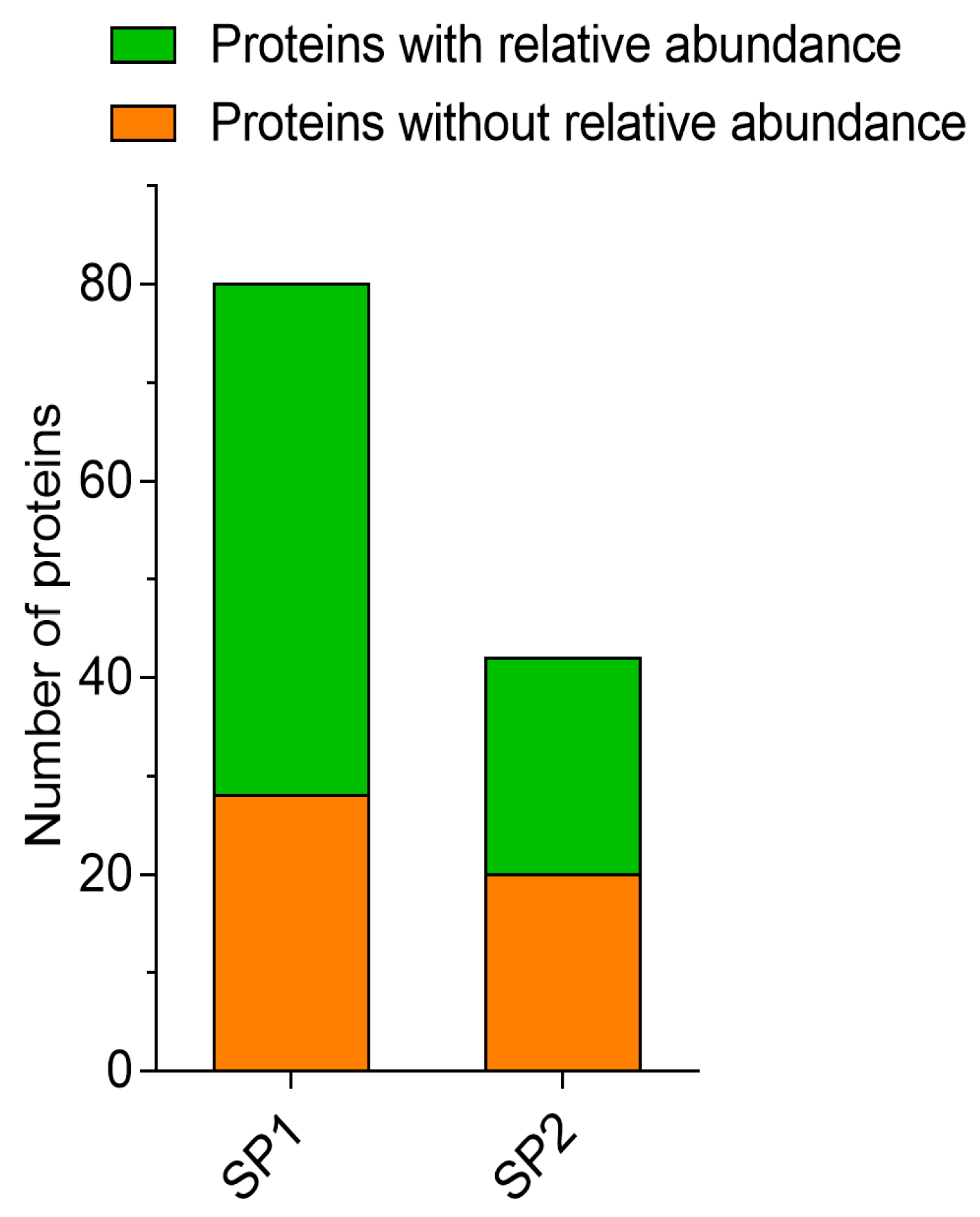
3.1. Analysis of Fractionated SP Proteins
3.1.1. LC–MS/MS Analysis
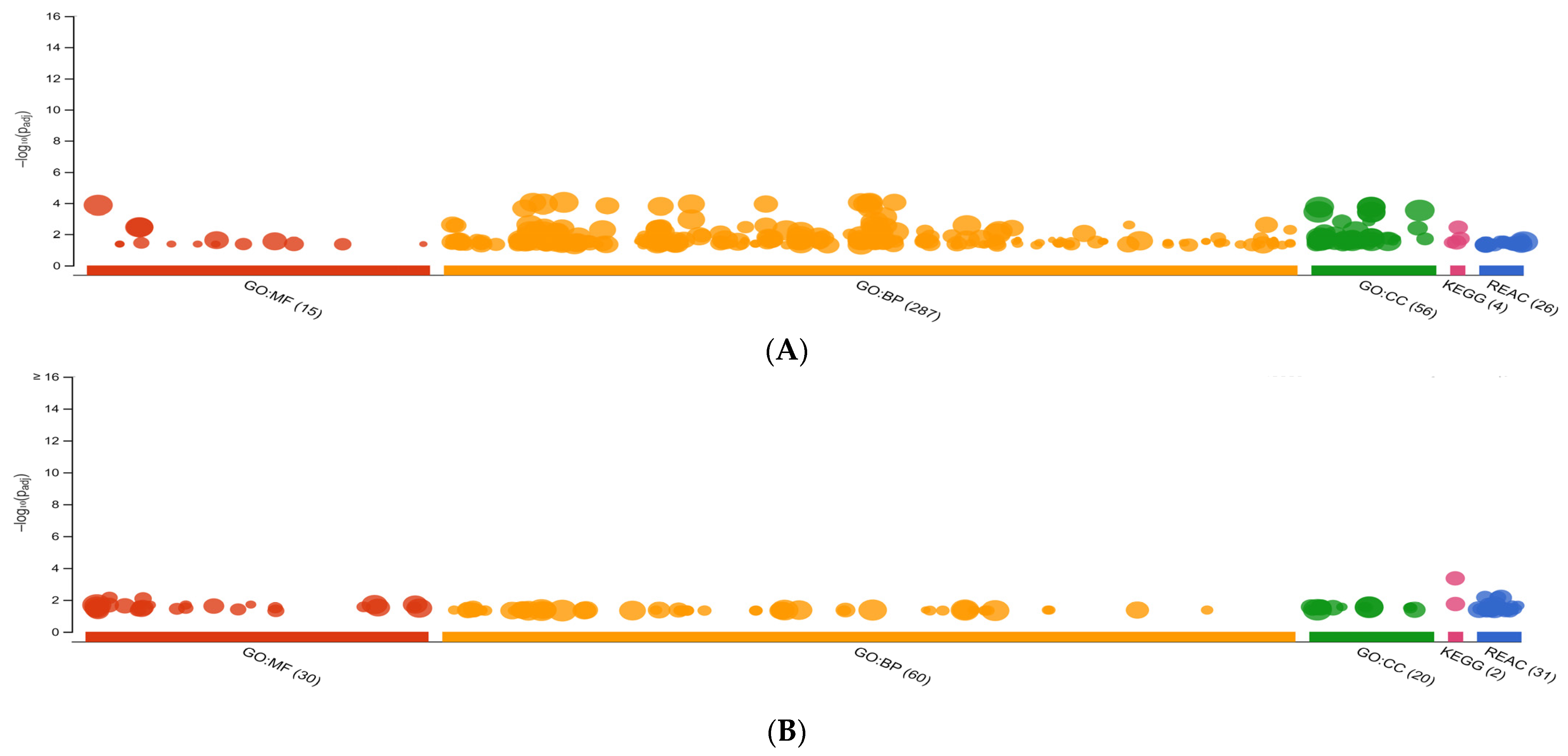
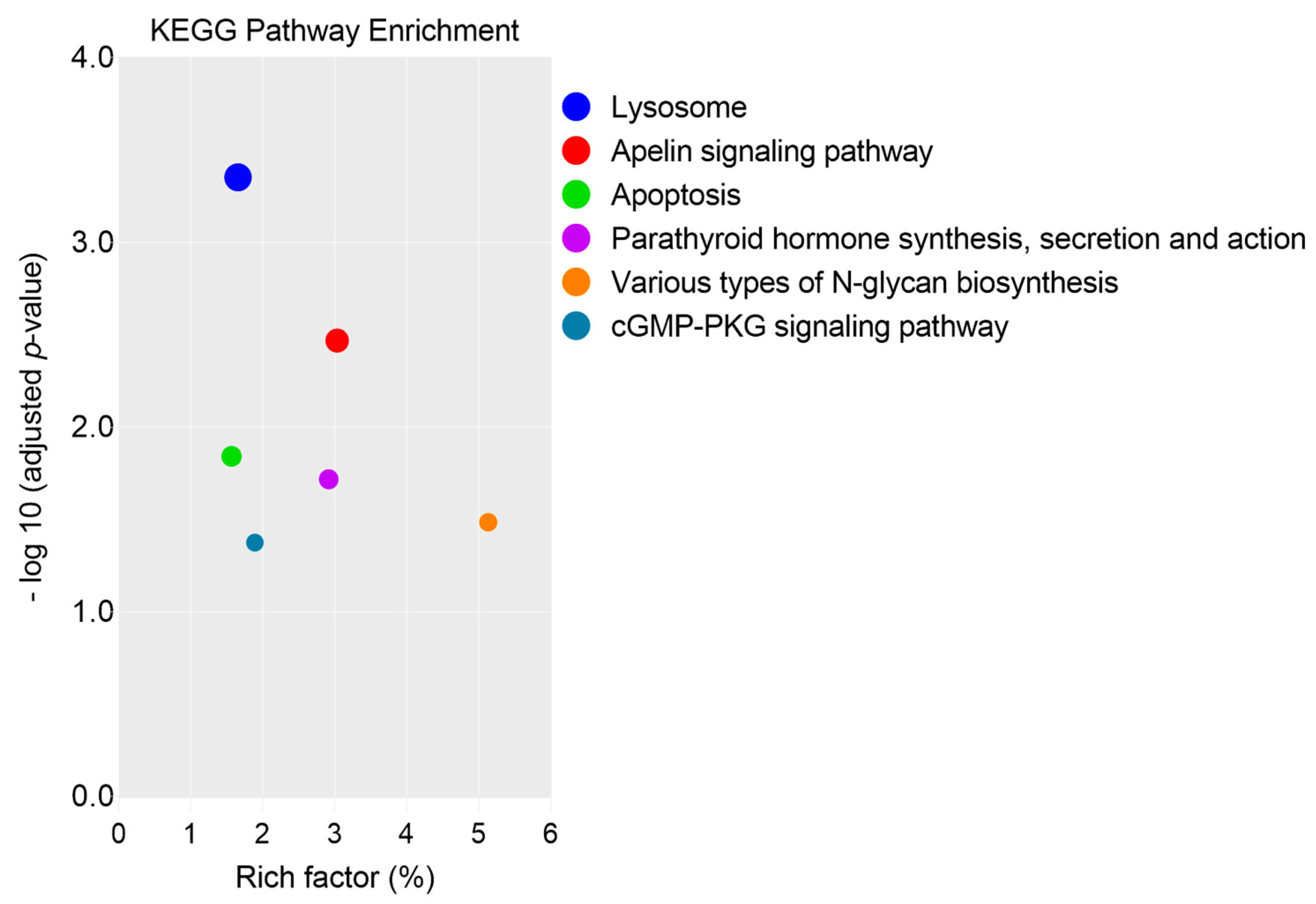
3.1.2. GO Analysis, KEGG, and REAC Pathways
3.1.3. Protein Classes of Fractionated SP
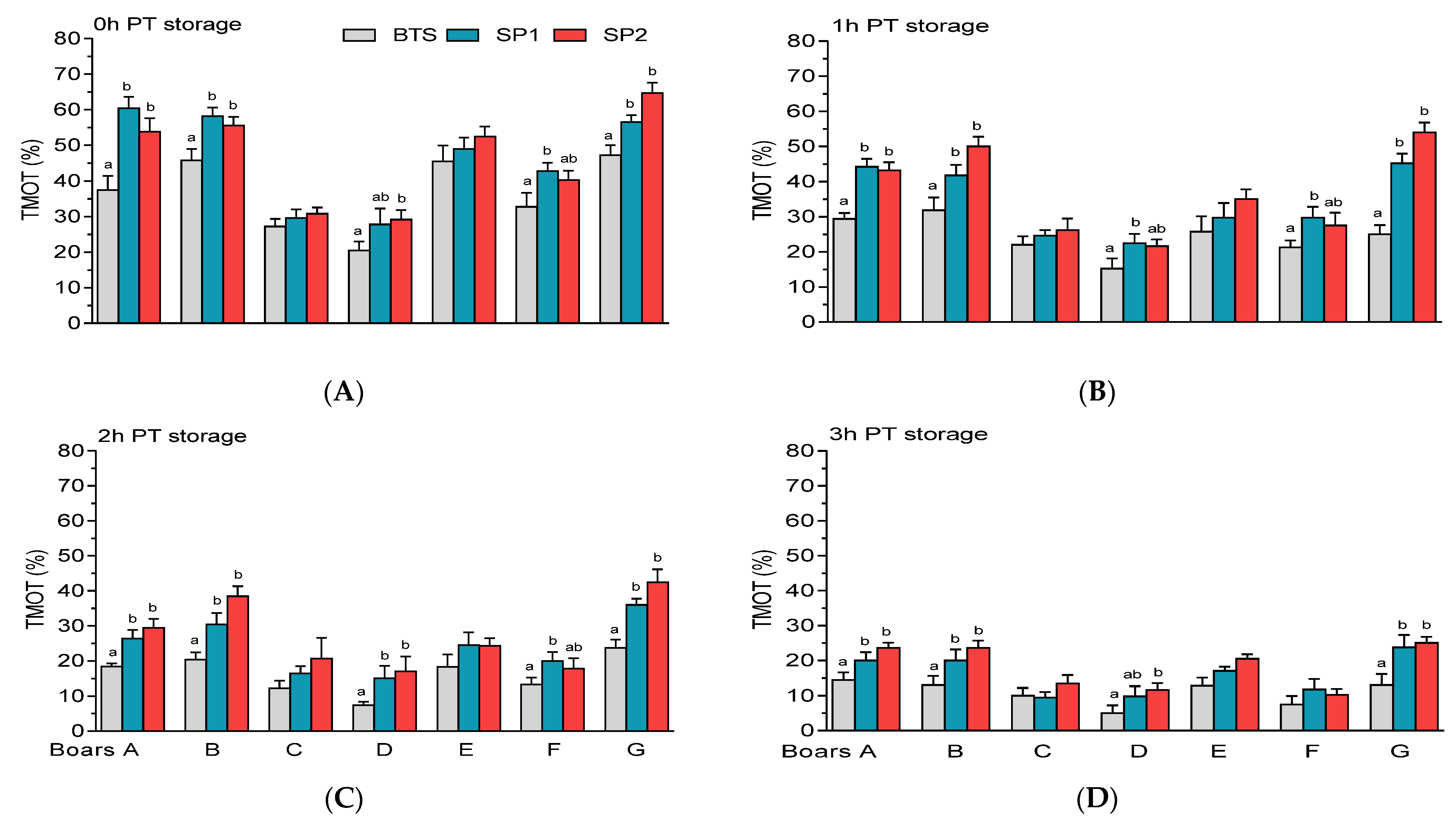
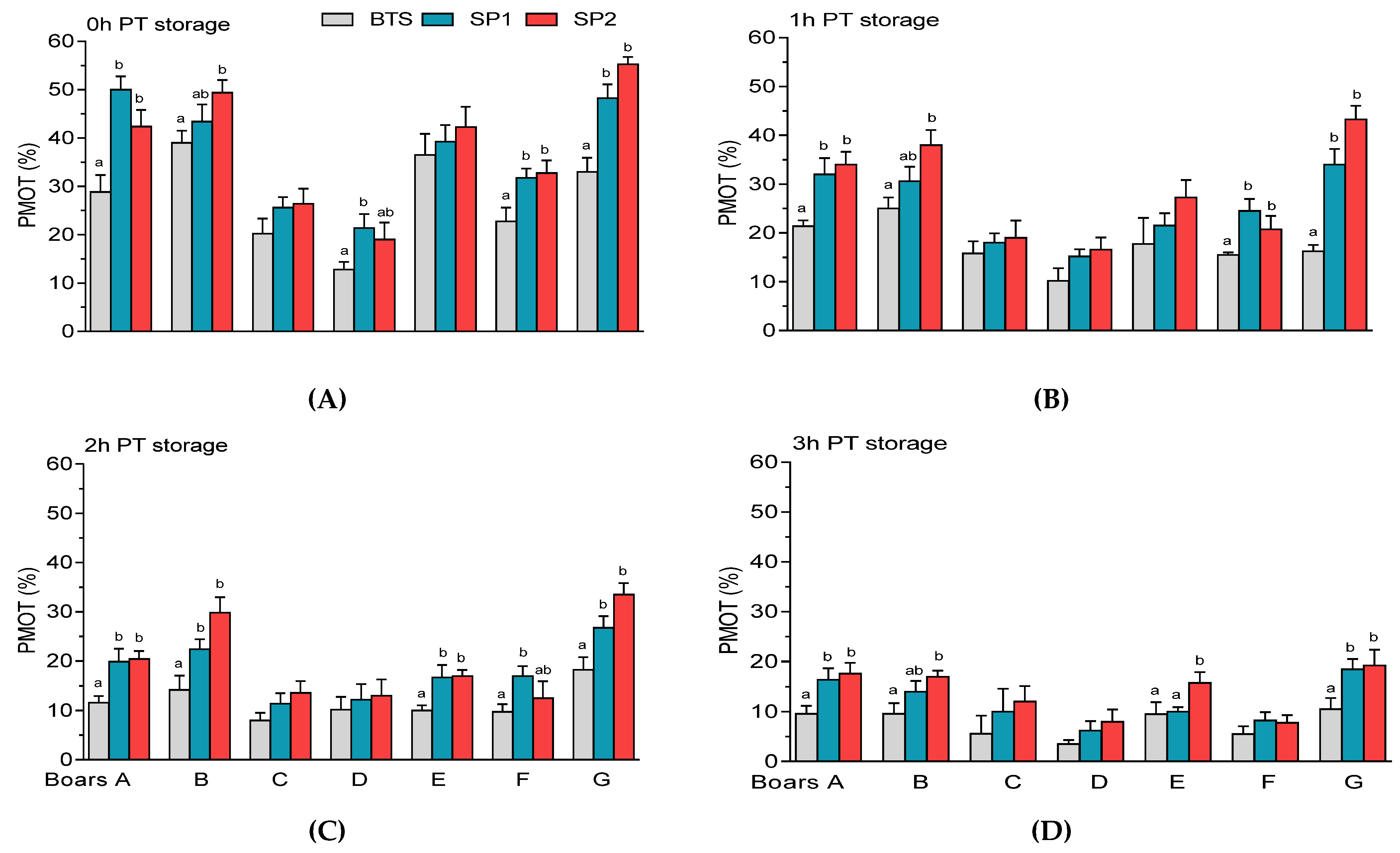
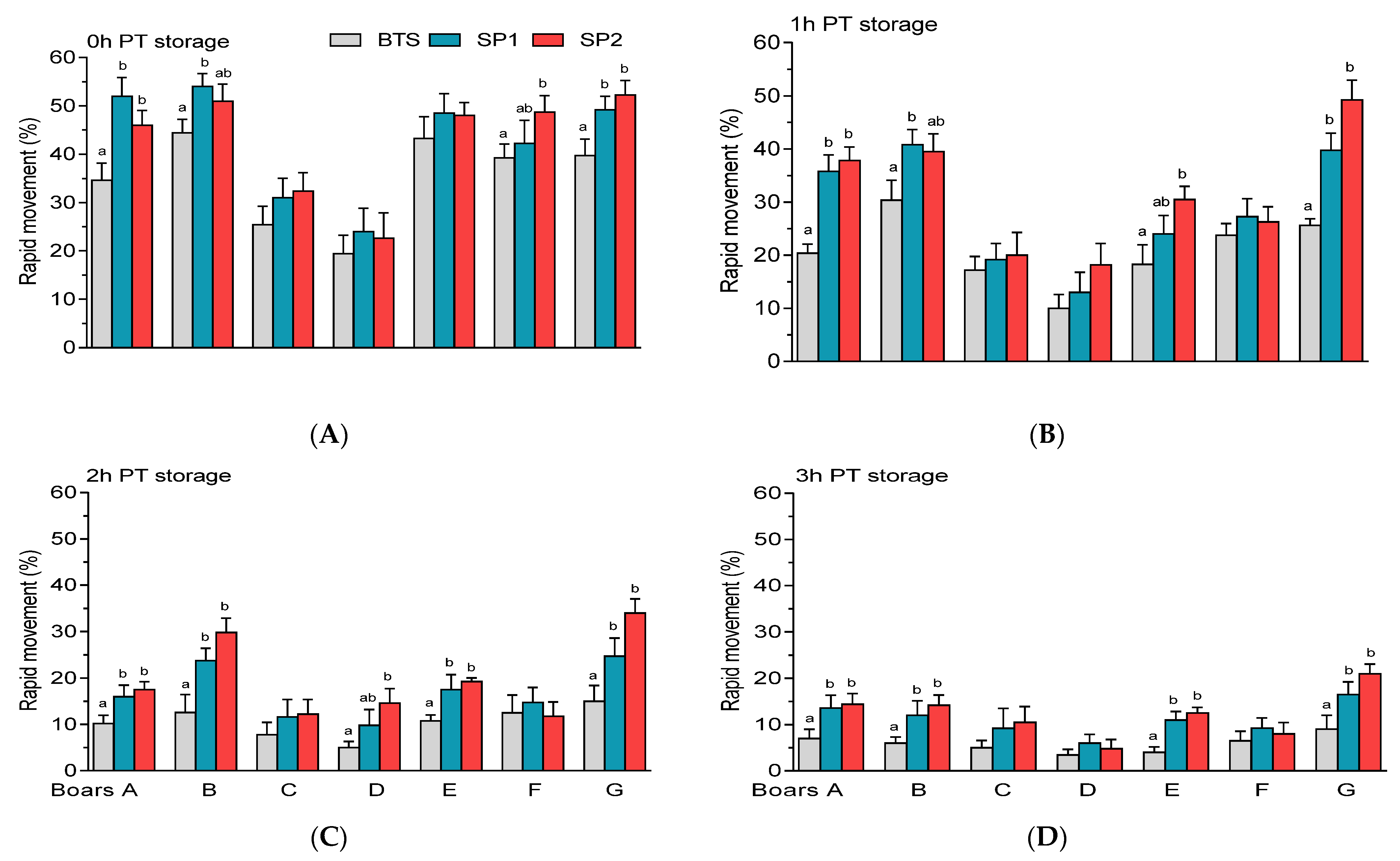
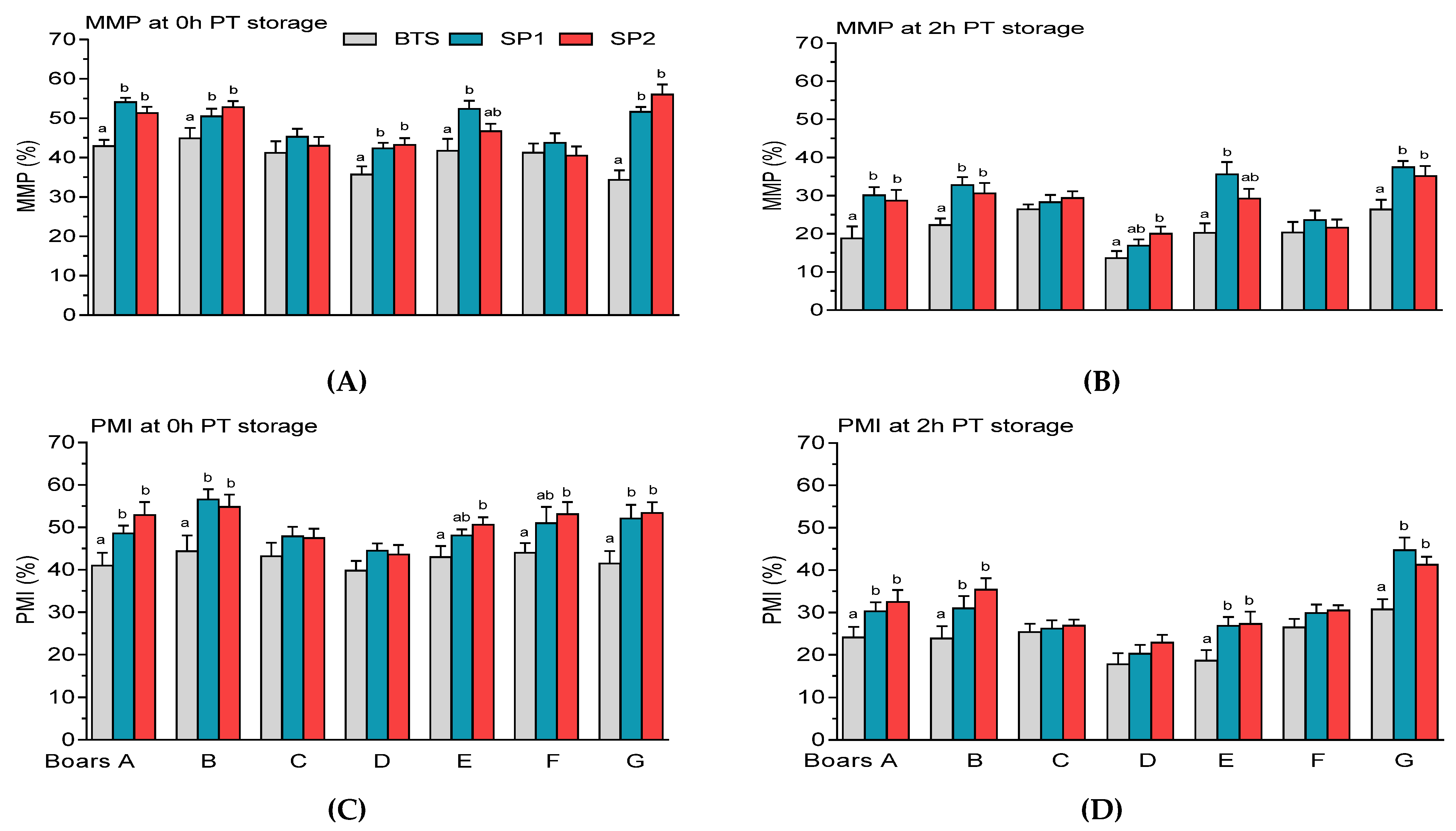
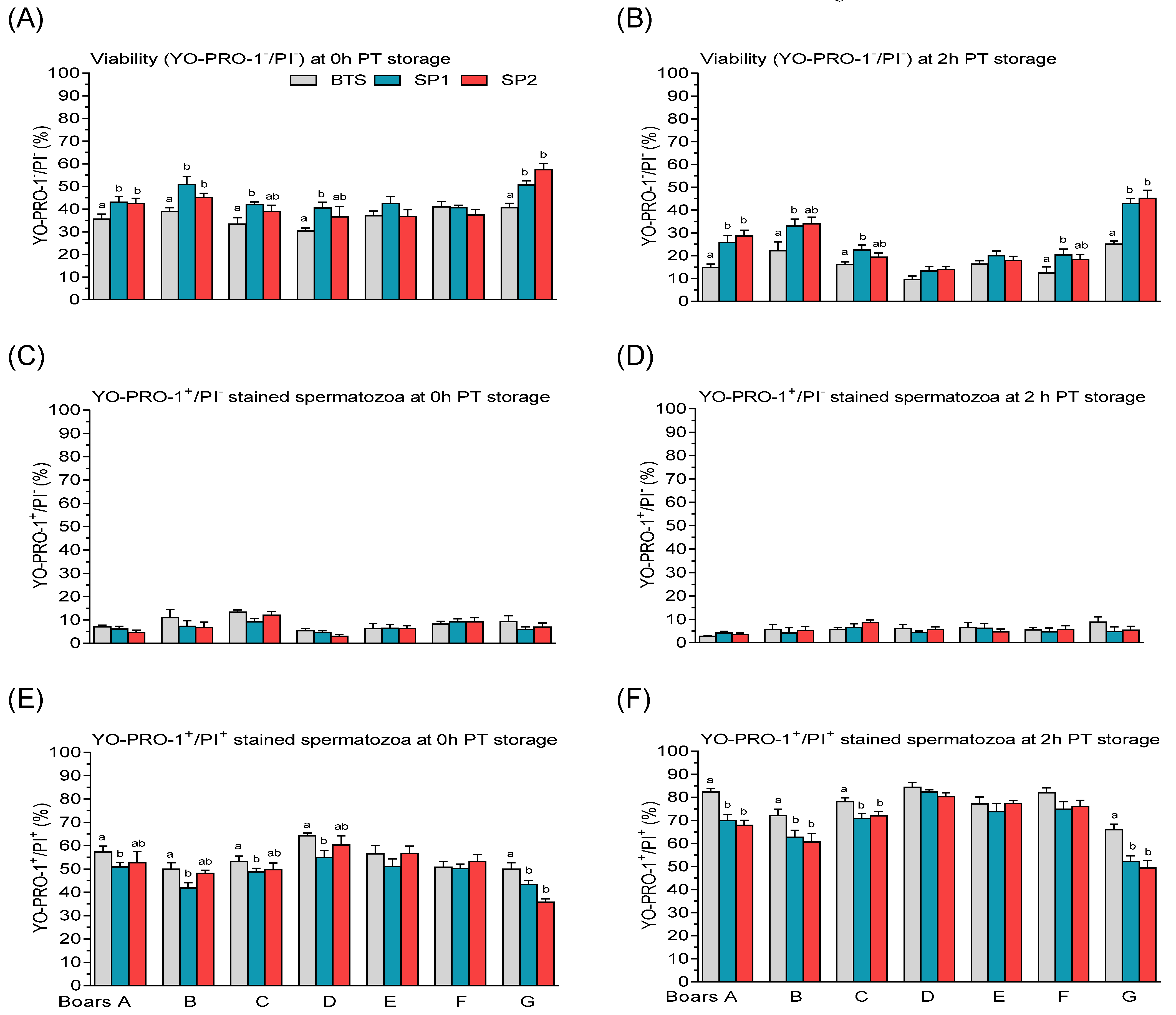
3.2. Post-Thaw Semen Assessment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Knox, R.V. The fertility of frozen boar sperm when used for artificial insemination. Reprod. Domest. Anim. 2015, 50, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Casas, I.; Sancho, S.; Briz, M.; Pinart, E.; Bussalleu, E.; Yeste, M.; Bonet, S. Fertility after post-cervical artificial insemination with cryopreserved sperm from boar ejaculates of good and poor freezability. Anim. Reprod. Sci. 2010, 118, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Uribe, H.F.J.; Giraldo, V.J.; Mesa, H.; Londoño, G.G. Effect of seminal plasma and sperm of boars valued by freezability on seminal cryopreservation. Rev. Fac. Nac. De Agron. 2016, 69, 7903–7910. [Google Scholar] [CrossRef]
- Valencia, J.; Gómez, G.; López, W.; Mesa, H.; Henao, F.J. Relationship between HSP90a, NPC2 and L-PGDS proteins to boar semen freezability. J. Anim. Sci. Biotechnol. 2017, 8, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valencia, J.; Yeste, M.; Quintero-Moreno, A.; Niño-Cardenas, C.D.P.; Henao, F.J. Relative content of Niemann-Pick C2 protein (NPC2) in seminal plasma, but not that of spermadhesin AQN-1, is related to boar sperm cryotolerance. Theriogenology 2020, 14, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Yeste, M. Sperm cryopreservation update: Cryodamage, markers, and factors affecting the sperm freezability in pigs. Theriogenology 2016, 85, 47–64. [Google Scholar] [CrossRef] [PubMed]
- Flowers, W.L.; Deller, F.; Stewart, K.R. Use of heterospermic inseminations and paternity testing to evaluate the relative contributions of common sperm traits and seminal plasma proteins in boar fertility. Anim Reprod. Sci. 2016, 17, 123–131. [Google Scholar] [CrossRef]
- Perez-Patiño, C.; Parrilla, I.; Barranco, I.; Vergara-Barberán, M.; Simo-Alfonso, E.F.; Herrero-Martínez, J.M.; Rodríguez-Martínez, H.; Martínez, E.A.; Roca, J. New in-depth analytical approach of the porcine seminal plasma proteome reveals potential fertility biomarkers. J. Proteome Res. 2018, 17, 1065–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Martinez, H.; Martinez, E.A.; Calvete, J.J.; Peña Vega, F.J.; Roca, J. Seminal plasma: Relevant for fertility? Int. J. Mol. Sci. 2021, 22, 4368. [Google Scholar] [CrossRef]
- De Lazari, L.F.; Sontag, E.S.; Schneider, A.; Moura, A.A.; Vasconcelos, F.R.; Nagano, C.S.; Dalberto, P.F.; Bizarro, C.V.; Mattos, R.C.; Jobim, M.I.M.; et al. Proteomic identification of boar seminal plasma proteins related to sperm resistance to cooling at 17 °C. Theriogenology 2020, 147, 135–145. [Google Scholar] [CrossRef]
- Gomes, F.P.; Park, R.; Viana, A.G.; Fernandez-Costa, C.; Topper, E.; Kaya, A.; Memili, E.; Yates III, J.R.; Moura, A.A. Protein signatures of seminal plasma from bulls with contrasting frozen-thawed sperm viability. Sci. Rep. 2020, 10, 14661. [Google Scholar] [CrossRef]
- Jia, B.; Liang, J.; Lv, C.; Memon, S.; Fang, Y.; Wu, G.; Quan, G. The characteristics of proteome and metabolome associated with contrasting sperm motility in goat seminal plasma. Sci. Rep. 2021, 11, 15562. [Google Scholar] [CrossRef] [PubMed]
- Perez-Patiño, C.; Barranco, I.; Parrilla, I.; Valero, M.L.; Martínez, E.A.; Rodríguez-Martínez, H.; Roca, J. Characterization of the porcine seminal plasma proteome comparing ejaculate portions. J. Proteom. 2016, 142, 15–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, M.; Roca, J.; Calvete, J.J.; Sanz, L.; Muino-Blanco, T.; Cebrian-Perez, J.A.; Vázquez, J.M.; Martínez, E.A. Cryosurvival and in vitro fertilizing capacity post-thaw is improved when boar spermatozoa are frozen in the presence of seminal plasma from good freezer boars. J. Androl. 2007, 28, 689–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravia, F.; Wallgren, M.; Johannisson, A.; Calvete, J.J.; Sanz, L.; Pena, F.J.; Roca, J.; Rodriguez-Martinez, H. Exposure to the seminal plasma of different portions of the boar ejaculate modulates the survival of spermatozoa cryopreserved in MiniFlatPacks. Theriogenology 2008, 71, 662–675. [Google Scholar] [CrossRef]
- Okazaki, T.; Abe, S.; Yoshida, S.; Shimada, M. Seminal plasma damages sperm during cryopreservation, but its presence during thawing improves semen quality and conception rates in boars with poor post-thaw semen quality. Theriogenology 2009, 71, 491–498. [Google Scholar] [CrossRef]
- Alkmin, D.V.; Perez-Patińo, C.; Barranco, I.; Parrilla, I.; Vázquez, J.; Martínez, E.A.; Rodríguez-Martínez, H.; Roca, J. Boar sperm cryosurvival is better after exposure to seminal plasma from selected fractions than to those from entire ejaculate. Cryobiology 2014, 69, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Barranco, I.; Tvarijonaviciute, A.; Molina, M.F.; Martínez, E.A.; Rodríguez-Martínez, H.; Parrilla, I.; Roca, J. Seminal plasma antioxidants are directly involved in boar sperm cryotolerance. Theriogenology 2018, 107, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Torres, M.A.; Monterio, M.S.; Passarelli, M.S.; Papa, F.O.; Dell’Agua, J.A., Jr.; Alvarenga, M.A.; Martins, S.M.M.K.; de Andrade, A.F.C. The ideal holding time for boar semen is 24 h at 17 °C prior to short-cryopreservation protocols. Cryobiology 2019, 86, 58–64. [Google Scholar] [CrossRef]
- Garcia, J.C.; Dominguez, J.C.; Pena, F.J.; Alegre, B.; Gonzalez, R.; Castro, M.J.; Habing, G.G.; Kirkwood, R.N. Thawing boar semen in the presence of seminal plasma: Effects on sperm quality and fertility. Anim. Reprod. Sci. 2010, 119, 160–165. [Google Scholar] [CrossRef]
- Vadnais, M.L.; Roberts, K.P. Seminal plasma proteins inhibit in vitro- and cooling-induced capacitation in boar spermatozoa. Reprod. Fertil. Dev. 2010, 22, 893–900. [Google Scholar] [CrossRef]
- Torres, M.A.; Ravagnani, G.M.; Leal, D.F.; Martins, M.M.K.; Muro, B.B.D.; Meirelles, F.V.; Papa, F.O.; Junior, J.A.D.; Alvarenga, M.A.; Moretti, A.S.; et al. Seminal plasma arising from the whole boar sperm-rich fraction increases the stability of sperm membrane after thawing. J. Anim. Sci. 2016, 94, 1906–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, F.; Chen, Y.; Guo, C.; Li, C.; Wei, H.; Li, L.; Meng, L.; Zhang, S. Analysis of differentially abundant proteins related to boar fertility in seminal plasma using iTRAQ-based quantitative proteomics. J. Proteom. 2021, 236, 104120. [Google Scholar] [CrossRef]
- Fraser, L.; Strzeżek, J. Effect of different procedures of ejaculate collection, extenders and packages on DNA integrity of boar spermatozoa following freezing-thawing. Anim. Reprod. Sci. 2007, 99, 317–329. [Google Scholar] [CrossRef]
- Jonáková, V.; Tichá, M. Boar seminal plasma proteins and their binding properties. A review. Collect. Czechoslov. Chem. Commun. 2004, 69, 461–475. [Google Scholar] [CrossRef]
- Perez-Patiño, C.; Parrilla, I.; Li, J.; Barranco, I.; Martínez, E.A.; Rodríguez-Martínez, H.; Roca, J. The proteome of pig spermatozoa is remodeled during ejaculation. Mol. Cell. Proteom. 2019, 18, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Wasilewska-Sakowska, K.; Zasiadczyk, Ł.; Fraser, L. Effect of fractionated seminal plasma on sperm characteristics following cryopreservation of boar semen. Ann. Anim. Sci. 2019, 19, 695–712. [Google Scholar] [CrossRef] [Green Version]
- Wasilewska-Sakowska, K.; Zasiadczyk, Ł.; Fraser, L.; Strzeżek, J.; Karpiesiuk, K. Effect of post-thaw supplementation of fractionated seminal plasma on survival of boar spermatozoa. Pol. J. Vet. Sci. 2019, 22, 617–625. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Lipecka, J.; Chhuon, C.; Bourderioux, M.; Bessard, M.A.; van Endert, P.; Edelman, A.; Guerrera, I.C. Sensitivity of mass spectrometry analysis depends on the shape of the filtration unit used for filter aided sample preparation (FASP). Proteomics 2016, 16, 1852–1857. [Google Scholar] [CrossRef]
- Cox, J.; Hein, M.Y.; Luber, C.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A peptide search engine integrated into the MaxQuant environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [Green Version]
- Mi, H.; Ebert, D.; Muruganujan, A.; Millis, C.; Albou, L.P.; Mushayamaha, T.; Thomas, P.D. PANTHER version 16: A revised family classification, tree-based classification tool, enhancer regions and extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef] [PubMed]
- Fonseka, P.; Pathan, M.; Chitti, S.V.; Kang, T.; Mathivanan, S. FunRich enables enrichment analysis of OMICS datasets. J. Mol. Biol. 2021, 433, 166747. [Google Scholar] [CrossRef]
- Fraser, L.; Parda, A.; Filipowicz, K.; Strzeżek, J. Comparison of post-thaw DNA integrity of boar spermatozoa assessed with the neutral Comet assay and sperm-Sus Halomax test kit. Reprod. Domest. Anim. 2010, 45, e155–e160. [Google Scholar] [CrossRef] [PubMed]
- Wasilewska, K.; Fraser, L. Boar variability in sperm cryo-tolerance after cooling of semen in different long term extenders at various temperatures. Anim. Reprod. Sci. 2017, 185, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Zasiadczyk, Ł.; Fraser, L.; Kordan, W.; Wasilewska, K. Individual and seasonal variations in the quality of fractionated boar ejaculates. Theriogenology 2015, 83, 1287–1303. [Google Scholar] [CrossRef] [PubMed]
- Dziekońska, A.; Fraser, L.; Strzeżek, J. Effect of different storage temperatures on the metabolic activity of spermatozoa following liquid storage of boar semen. J. Anim. Feed. Sci. 2009, 18, 638649. [Google Scholar] [CrossRef]
- Fraser, L.; Strzeżek, J.; Kordan, W. Post-thaw sperm characteristics following long-term storage of boar semen in liquid nitrogen. Anim. Reprod. Sci. 2014, 147, 119–127. [Google Scholar] [CrossRef]
- Garner, D.L.; Johnson, L.A. Viability assessment of mammalian sperm using SYBR-14 and propidium iodide. Biol. Reprod. 1995, 53, 276–284. [Google Scholar] [CrossRef]
- Soler, A.J.; Esteso, M.C.; Fernandez-Santos, M.R.; Garde, J.J. Characteristics of Iberian red deer (Cervus elaphus hispanicus) spermatozoa cryopreserved after storage at 5 degrees C in the epididymis for several days. Theriogenology 2005, 64, 1503–1517. [Google Scholar] [CrossRef]
- Aquino-Cortez, A.; Pinheiro, B.Q.; Lima, D.B.C.; Silva, H.V.R.; Mota-Filho, A.C.; Martins, J.A.M.; Rodriguez-Villamil, P.; Moura, A.A.; Silva, L.D.M. Proteomic characterization of canine seminal plasma. Theriogenology 2017, 95, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Jodar, M.; Sendler, E.; Krawetz, S.A. The protein and transcript profiles of human semen. Cell Tissue Res. 2016, 363, 85–96. [Google Scholar] [CrossRef]
- Waberski, D.; Schäfer, J.; Bölling, A.; Scheld, M.; Henning, H.; Hambruch, N.; Schuberth, H.J.; Pfarrer, C.; Wrenzycki, C.; Hunter, R.H.F. Seminal plasma modulates the immune-cytokine network in the porcine uterine tissue and pre-ovulatory follicles. PLoS ONE 2018, 13, e0202654. [Google Scholar] [CrossRef] [Green Version]
- Martinez, C.A.; Cambra, J.M.; Gil, M.A.; Parrilla, I.; Alvarez-Rodriguez, M.; Rodriguez-Martinez, H.; Cuello, C.; Martinez, E.A. Seminal plasma induces overexpression of genes associated with embryo development and implantation in Day-6 porcine blastocysts. Int. J. Mol. Sci. 2020, 21, 3662. [Google Scholar] [CrossRef] [PubMed]
- Kurowska, P.; Barbe, A.; Różycka, M.; Chmielińska, J.; Dupont, J.; Rak, A. Apelin in reproductive physiology and pathology of different species: A Critical Review. Int. J. Endocrinol. 2018, 2018, 9170480. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lu, J.; Sun, X.; Pang, Q.; Zhao, Y. Molecular cloning, mRNA expression, and localization of the G-protein subunit Galphaq in sheep testis and epididymis. Asian-Australas. J. Anim. Sci. 2016, 29, 1702–1709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merlet, F.; Weinstein, L.S.; Goldsmith, P.K.; Rarick, T.; Hall, J.L. Identification and localization of G protein subunits in human spermatozoa. Mol. Human Reprod. 1999, 5, 38–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revelli, A.; Ghigo, D.; Moffa, F.; Massobrio, M.; Tur-Kaspa, I. Guanylate cyclase activity and sperm function. Endrocri. Rev. 2002, 23, 484–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wysocki, P.; Orzo1ek, A.; Strzeżek, J.; Koziorowska-Gilun, M.; Zasiadczyk, Ł.; Kordan, W. The activity of N-acetyl-b-hexosaminidase in boar seminal plasma is linked with semen quality and its suitability for cryopreservation. Theriogenology 2015, 83, 1194–2202. [Google Scholar] [CrossRef]
- Álvarez-Rodríguez, M.; Martinez, C.A.; Wright, D.; Rodríguez-Martinez, H. The role of semen and seminal plasma in inducing large-scale genomic changes in the female porcine peri-ovulatory tract. Sci. Rep. 2020, 10, 5061. [Google Scholar] [CrossRef] [Green Version]
- De Lazari, F.L.; Sontag, E.R.; Schneider, A.; Moura, A.A.A.; Vasconcelos, F.R.; Nagano, C.S.; Mattos, R.C.; Jobim, M.I.M.; Bustamante-Filho, I.C. Seminal plasma proteins and their relationship with sperm motility and morphology in boars. Andrologia 2018, 51, e13222. [Google Scholar] [CrossRef]
- Richard, J.P.; Leahy, T.; Soleihavoup, C.; Tsikis, G.; Labas, V.; Harichaux, G.; Lynch, G.W.; Druart, X.; de Graaf, S.P. The identification of proteomics markers of sperm freezing resilience in ram seminal plasma. J. Proteom. 2015, 126, 303–311. [Google Scholar] [CrossRef]
- Jain, A.K.; Allton, K.; Aundrietta, D.; Duncan, A.D.; Michelle, C.; Barton, M.C. TRIM24 is a p53-induced E3-ubiquitin ligase that undergoes ATM-mediated phosphorylation and autodegradation during DNA damage. Mol. Cell. Biol. 2014, 34, 2695–2709. [Google Scholar] [CrossRef] [Green Version]
- Légaré, C.; Thabet, M.; Gatti, J.L.; Sullivan, R. HE1/NPC2 status in human reproductive tract and ejaculated spermatozoa: Consequence of vasectomy. Mol. Hum. Reprod. 2006, 12, 461–468. [Google Scholar] [CrossRef]
- Fouchecourt, S.; Metayer, S.; Locatelli, A.; Dacheux, F.; Dacheux, J.L. Stallion epididymal fluid proteome: Qualitative and quantitative characterization; secretion and dynamic changes of major proteins. Biol. Reprod. 2000, 62, 1790–1803. [Google Scholar] [CrossRef] [PubMed]
- Belleannée, C.; Labas, V.; Teixeira-Gomes, A.P.; Gatti, J.L.; Dacheux, J.L.; Dacheux, F. Identification of luminal and secreted proteins in bull epididymis. J. Proteom. 2011, 74, 59–78. [Google Scholar] [CrossRef] [PubMed]
- Girourard, J.; Frenette, G.; Sullivan, R. Seminal plasma proteins regulate the association of lipids and proteins within detergent-resistant membrane domains of bovine spermatozoa. Biol. Reprod. 2008, 78, 921–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero, I.; Vazquez, J.M.; Mayor, G.M.; Alminana, C.; Calvete, J.J.; Sanz, L.; Roca, J.; Martinez, E.A. PSP-I/PSP-II spermadhesin exert a decapacitation effect on highly extended boar spermatozoa. Int. J. Androl. 2009, 32, 505–513. [Google Scholar] [CrossRef]
- González-Cadavid, V.; Martins, J.A.M.; Moreno, F.B.; Andrade, T.S.; Santos, A.C.L.; Monteiro Moreira, A.C.O.; Moreira, R.A.; Moura, A.A. Seminal plasma proteins of adult boars and correlations with sperm parameters. Theriogenology 2014, 82, 697–707. [Google Scholar] [CrossRef] [Green Version]
- Vilagran, I.; Yeste, M.; Sancho, S.; Castillo, J.; Oliva, R.; Bonet, S. Comparative analysis of boar seminal plasma proteome from different freezability ejaculates and identification of Fibronectin 1 as sperm freezability marker. Andrology 2015, 3, 345–356. [Google Scholar] [CrossRef]
- Ma, D.D.; Wang, D.H.; Yang, W.X. Kinesins in spermatogenesis. Biol. Reprod. 2017, 96, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Tomar, A.K.; Sooch, B.S.; Raj, I.; Singh, S.; Singh, T.P.; Yadav, S. Isolation and identification of Concanavalin A binding glycoproteins from human seminal plasma: A step towards identification of male infertility marker proteins. Dis. Markers 2011, 31, 379–386. [Google Scholar] [CrossRef]
- Fitzgerald, J.S.; Toth, B.; Jeschke, U.; Schleussner, E.; Markert, U.R. Knocking off the suppressors of cytokine signaling (SOCS): Their roles in mammalian pregnancy. J. Reprod. Immunol. 2009, 83, 117–123. [Google Scholar] [CrossRef]
- Pizzagalli, M.D.; Bensimon, A.; Superti-Furga, G. A guide to plasma membrane solute carrier proteins. FEBS J. 2021, 288, 2784–2835. [Google Scholar] [CrossRef]
- Atikuzzaman, M.; Alvarez-Rodriguez, M.; Vicente-Carrillo, A.; Johnsson, M.; Wright, D.; Rodriguez-Martinez, H. Conserved gene expression in sperm reservoirs between birds and mammals in response to mating. BMC Genom. 2017, 18, 563. [Google Scholar] [CrossRef] [Green Version]
- Yuan, S.; Swiggin, H.M.; Zheng, H.; Yan, W. A testis-specific gene, Ubqlnl, is dispensable for mouse embryonic development and spermatogenesis. Mol. Reprod. Dev. 2015, 82, 408–4049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimarães, D.B.; Barros, T.B.; van Tilburg, M.F.; Martins, J.A.M.; Moura, A.A.; Moreno, F.B.; Monteiro-Moreira, A.C.; Moreira, R.A.; Toniolli, R. Sperm membrane proteins associated with the boar semen cryopreservation. Anim. Reprod. Sci. 2017, 183, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Gago; Domínguez, J.C.; Martínez-Pastor, F. Seminal plasma applied post-thawing affects boar sperm physiology: A flow cytometry study. Theriogenology 2013, 80, 400–410. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sperm Parameters (%) | Means ± SEM (n = 32) |
|---|---|
| Total motility (TMOT) | 86.97 ± 1.06 |
| Progressive motility (PMOT) | 59.52 ± 1.59 |
| Rapid movement (%) | 60.06 ± 2.70 |
| Mitochondrial membrane potential (MMP) | 85.80 ± 0.35 |
| Plasma membrane integrity (PMI) | 86.98 ± 0.31 |
| Acrosome integrity (%) | 88.94 ± 0.37 |
| Viability (YO-PRO-1−/PI−) | 86.16 ± 0.66 |
| Plasma membrane apoptotic-like changes (YO-PRO-1+/PI−) | 5.81 ± 0.35 |
| Dead spermatozoa (YO-PRO-1+/PI+) | 8.03 ± 0.41 |
| Sperm | Boar | Treatment | PT Storage | Boar × Treatment | Boar × PT Storage |
|---|---|---|---|---|---|
| Parameters | p-Value | p-Value | p-Value | p-Value | p-Value |
| (A) | |||||
| Total motility (TMOT) | 0.001 | 0.001 | 0.001 | 0.009 | 0.001 |
| Progressive motility (PMOT) | 0.001 | 0.001 | 0.001 | 0.008 | 0.001 |
| Rapid movement | 0.001 | 0.001 | 0.001 | n.s | 0.003 |
| (B) | |||||
| Mitochondrial membrane potential (MMP) | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 |
| Plasma membrane integrity (PMI) | 0.001 | 0.001 | 0.001 | n.s | 0.001 |
| Acrosome integrity | 0.001 | 0.001 | 0.001 | 0.019 | 0.001 |
| Viability (YO-PRO-1−/PI−) | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 |
| Plasma membrane apoptotic-like changes (YO-PRO-1+/PI−) | 0.001 | n.s | n.s | n.s | 0.043 |
| Dead spermatozoa (YO-PRO-1+/PI+) | 0.00.1 | 0.001 | 0.001 | 0.017 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraser, L.; Wasilewska-Sakowska, K.; Zasiadczyk, Ł.; Piątkowska, E.; Karpiesiuk, K. Fractionated Seminal Plasma of Boar Ejaculates Analyzed by LC–MS/MS: Its Effects on Post-Thaw Semen Quality. Genes 2021, 12, 1574. https://doi.org/10.3390/genes12101574
Fraser L, Wasilewska-Sakowska K, Zasiadczyk Ł, Piątkowska E, Karpiesiuk K. Fractionated Seminal Plasma of Boar Ejaculates Analyzed by LC–MS/MS: Its Effects on Post-Thaw Semen Quality. Genes. 2021; 12(10):1574. https://doi.org/10.3390/genes12101574
Chicago/Turabian StyleFraser, Leyland, Karolina Wasilewska-Sakowska, Łukasz Zasiadczyk, Elżbieta Piątkowska, and Krzysztof Karpiesiuk. 2021. "Fractionated Seminal Plasma of Boar Ejaculates Analyzed by LC–MS/MS: Its Effects on Post-Thaw Semen Quality" Genes 12, no. 10: 1574. https://doi.org/10.3390/genes12101574