
Association and Genetic Expression between Genes Involved in HPA Axis and Suicide Behavior: A Systematic Review

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Methods
2.1. Search Strategy
2.2. Inclusion Criteria and Data Extraction
2.3. Quality Assessment of Primary Studies
2.4. Data Synthesis
3. Results
3.1. Study Selection
3.2. Studies Caracteristics
3.3. Phenotypes and Genes
3.4. Synthesis of Results
3.4.1. FKBP5 Gene
3.4.2. CRH Gene
3.4.3. CRHBP Gene
3.4.4. CRHR1 Gene
3.4.5. CRHR2 Gene
3.4.6. NR3C1 Gene
3.4.7. NR3C2 Gene
3.4.8. SKA2 Gene
3.4.9. MC2R Gene
3.4.10. POMC Gene
4. Discussion
4.1. Main Findings
4.2. HPA Axis and Suicide Behavior
4.3. Strengths and Limitations
4.4. Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nock, M.K.; Borges, G.; Bromet, E.J.; Cha, C.B.; Kessler, R.C.; Lee, S. Suicide and Suicidal Behavior. Epidemiol. Rev. 2008, 30, 133–154. [Google Scholar] [CrossRef] [PubMed]
- Silverman, M.M.; Berman, A.L.; Sanddal, N.D.; O’Carroll, P.W.; Joiner, T.E. Rebuilding the Tower of Babel: A Revised Nomenclature for the Study of Suicide and Suicidal Behaviors Part 2: Suicide-Related Ideations, Communications, and Behaviors. Suicide Life-Threat. Behav. 2007, 37, 264–277. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, S. Epidemiology of Suicide and the Psychiatric Perspective. Int. J. Environ. Res. Public Health 2018, 15, 1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turecki, G.; Brent, D.A. Suicide and suicidal behaviour. Lancet 2016, 387, 1227–1239. [Google Scholar] [CrossRef]
- Van Heeringen, K. Frontiers in Neuroscience. Stress–Diathesis Model of Suicidal Behavior. In The Neurobiological Basis of Suicide; Dwivedi, Y., Ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Boca Raton, FL, USA, 2012. Available online: https://pubmed.ncbi.nlm.nih.gov/23035289/ (accessed on 15 July 2021).
- Mann, J.J.; Rizk, M.M. A Brain-Centric Model of Suicidal Behavior. Am. J. Psychiatry 2020, 177, 902–916. [Google Scholar] [CrossRef] [PubMed]
- Baud, P. Personality traits as intermediary phenotypes in suicidal behavior: Genetic issues. Am. J. Med. Genet. Part C Semin. Med. Genet. 2005, 133C, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the Hypothalamic-Pituitary-Adrenocortical Stress Response. Compr. Physiol. 2016, 6, 603–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kloet, E.R.; Joels, M.; Holsboer, F. Stress and the brain: From adaptation to disease. Nat. Rev. Neurosci. 2005, 6, 463–475. [Google Scholar] [CrossRef]
- Kloet, E.R. Hormones and the Stressed Brain. Ann. N.Y. Acad. Sci. 2004, 1018, 1–15. [Google Scholar] [CrossRef]
- Lim, C.T.; Khoo, B.; Feingold, K.R.; Anawalt, B.; Boyce, A.; Chrousos, G.; de Herder, W.W.; Dhatariya, K.; Dungan, K.; Grossman, A.; et al. Normal Physiology of ACTH and GH Release in the Hypothalamus and Anterior Pituitary in Man. In Endotext; MDText.com, Inc.: South Dartmouth, MA, USA, 2020. Available online: https://pubmed.ncbi.nlm.nih.gov/25905340/ (accessed on 21 May 2021).
- Reul, J.M.M.; Holsboer, F. On the role of corticotropin-releasing hormone receptors in anxiety and depression. Dialog Clin. Neurosci. 2002, 4, 31–46. [Google Scholar] [CrossRef]
- Ketchesin, K.; Stinnett, G.S.; Seasholtz, A.F. Corticotropin-releasing hormone-binding protein and stress: From invertebrates to humans. Stress 2017, 20, 449–464. [Google Scholar] [CrossRef]
- Pérez-Ortiz, J.M.; García-Gutiérrez, M.S.; Navarrete, F.; Giner, S.; Manzanares, J. Gene and protein alterations of FKBP5 and glucocorticoid receptor in the amygdala of suicide victims. Psychoneuroendocrinology 2013, 38, 1251–1258. [Google Scholar] [CrossRef]
- Fries, G.R.; Gassen, N.; Schmidt, U.; Rein, T. The FKBP51-Glucocorticoid Receptor Balance in Stress-Related Mental Disorders. Curr. Mol. Pharmacol. 2015, 9, 126–140. [Google Scholar] [CrossRef]
- Rice, L.; Waters, E.C.; Eccles, J.; Garside, H.; Sommer, P.; Kay, P.; Blackhall, F.H.; Zeef, L.; Telfer, B.; Stratford, I.; et al. Identification and functional analysis of SKA2 interaction with the glucocorticoid receptor. J. Endocrinol. 2008, 198, 499–509. [Google Scholar] [CrossRef]
- O’Connor, D.B.; Gartland, N.; O’Connor, R.C. Stress, cortisol and suicide risk. Dev. Psychopathol. 2020, 152, 101–130. [Google Scholar] [CrossRef]
- Berardelli, I.; Serafini, G.; Cortese, N.; Fiaschè, F.; O’Connor, R.C.; Pompili, M. The Involvement of Hypothalamus–Pituitary–Adrenal (HPA) Axis in Suicide Risk. Brain Sci. 2020, 10, 653. [Google Scholar] [CrossRef]
- Segura, A.G.; Mitjans, M.; Jimenez, E.; Fatjó-Vilas, M.; Ruiz, V.; Saiz, P.; García-Portilla, M.; Gonzalez-Blanco, L.; Bobes, J.; Vieta, E.; et al. Association of childhood trauma and genetic variability of CRH-BP and FKBP5 genes with suicidal behavior in bipolar patients. J. Affect. Disord. 2019, 255, 15–22. [Google Scholar] [CrossRef]
- Park, S.; Hong, J.P.; Lee, J.-K.; Park, Y.-M.; Park, Y.; Jeon, J.; Ahn, M.H.; Yoon, S.C. Associations between the neuron-specific glucocorticoid receptor (NR3C1) Bcl-1 polymorphisms and suicide in cancer patients within the first year of diagnosis. Behav. Brain Funct. 2016, 12, 22. [Google Scholar] [CrossRef] [Green Version]
- Berent, D.; Szymańska, B.; Kulczycka-Wojdala, D.; Macander, M.; Pawłowska, Z.; Wojnar, M. The role of childhood adversities, FKBP5, BDNF, NRN1, and generalized self-efficacy in suicide attempts in alcohol-dependent patients. Pharmacol. Rep. Pr. 2020, 72, 730–743. [Google Scholar] [CrossRef]
- Hernández-Díaz, Y.; Genis-Mendoza, A.; Fresán, A.; González-Castro, T.; Tovilla-Zárate, C.; Juárez-Rojop, I.; López-Narváez, M.; Martínez-Magaña, J.; Nicolini, H. Knowledge, Emotions and Stressors in Front-Line Healthcare Workers during the COVID-19 Outbreak in Mexico. Int. J. Environ. Res. Public Heal. 2021, 18, 5622. [Google Scholar] [CrossRef]
- Sanabrais-Jiménez, M.A.; Sotelo-Ramirez, C.E.; Ordoñez-Martinez, B.; Jiménez-Pavón, J.; Ahumada-Curiel, G.; Piana-Diaz, S.; Flores-Flores, G.; Flores-Ramos, M.; Jiménez-Anguiano, A.; Camarena, B. Effect of CRHR1 and CRHR2 gene polymorphisms and childhood trauma in suicide attempt. J. Neural Transm. 2019, 126, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Wasserman, D.; Wasserman, J.; Rozanov, V.; Sokolowski, M. Depression in suicidal males: Genetic risk variants in theCRHR1gene. Genes Brain Behav. 2009, 8, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Kaminsky, A.Z.; Wilcox, H.; Eaton, W.W.; Van Eck, K.; Kilaru, V.; Jovanovic, T.; Klengel, T.; Bradley, B.P.; Binder, E.B.; Ressler, K.J.; et al. Epigenetic and genetic variation at SKA2 predict suicidal behavior and post-traumatic stress disorder. Transl. Psychiatry 2015, 5, e627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, G.N.; Rizavi, H.S.; Zhang, H.; Bhaumik, R.; Ren, X. The Expression of the Suicide-Associated Gene SKA2 Is Decreased in the Prefrontal Cortex of Suicide Victims but Not of Nonsuicidal Patients. Int. J. Neuropsychopharmacol. 2016, 19. [Google Scholar] [CrossRef] [Green Version]
- De Luca, V.; Tharmalingam, S.; Zai, C.; Potapova, N.; Strauss, J.; Vincent, J.; Kennedy, J. Association of HPA axis genes with suicidal behaviour in schizophrenia. J. Psychopharmacol. 2008, 24, 677–682. [Google Scholar] [CrossRef]
- Roy, A.; Hodgkinson, C.A.; DeLuca, V.; Goldman, D.; Enoch, M.-A. Two HPA axis genes, CRHBP and FKBP5, interact with childhood trauma to increase the risk for suicidal behavior. J. Psychiatr. Res. 2012, 46, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Pjevac, M.; Pregel, J.P. Neurobiology of suicidal behaviour. Psychiatr. Danub. 2012, 24, 336–341. [Google Scholar]
- Duval, F.; Mokrani, M.-C.; Monreal, J.; Weiss, T.; Fattah, S.; Hamel, B.; Macher, J.-P. Interaction between the serotonergic system and HPA and HPT axes in patients with major depression: Implications for pathogenesis of suicidal behavior. Dialog Clin. Neurosci. 2002, 4, 417. [Google Scholar] [CrossRef]
- Papiol, S.; Arias, B.; Gastó, C.; Gutiérrez, B.; Catalán, R.; Fañanás, L. Genetic variability at HPA axis in major depression and clinical response to antidepressant treatment. J. Affect. Disord. 2007, 104, 83–90. [Google Scholar] [CrossRef]
- Willour, V.L.; Chen, H.; Toolan, J.; Belmonte, P.; Cutler, D.J.; Goes, F.S.; Zandi, P.P.; Lee, R.; MacKinnon, D.F.; Mondimore, F.M.; et al. Family-based association of FKBP5 in bipolar disorder. Mol. Psychiatry 2008, 14, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Brent, D.; Melhem, N.; Ferrell, R.; Emslie, G.; Wagner, K.D.; Ryan, N.; Vitiello, B.; Birmaher, B.; Mayes, T.; Zelazny, J.; et al. Association of FKBP5 Polymorphisms with Suicidal Events in the Treatment of Resistant Depression in Adolescents (TORDIA) Study. Am. J. Psychiatry 2010, 167, 190–197. [Google Scholar] [CrossRef]
- Roy, A.; Gorodetsky, E.; Yuan, Q.; Goldman, D.; Enoch, M.A. Interaction of FKBP5, a stress-related gene, with childhood trauma increases the risk for attempting suicide. Neuropsychopharmacoly 2010, 35, 1674–1683. [Google Scholar] [CrossRef] [Green Version]
- Perroud, N.; Bondolfi, G.; Uher, R.; Gex-Fabry, M.; Aubry, J.-M.; Bertschy, G.; Malafosse, A.; Kosel, M. Clinical and genetic correlates of suicidal ideation during antidepressant treatment in a depressed outpatient sample. Pharmacogenomics 2011, 12, 365–377. [Google Scholar] [CrossRef]
- Supriyanto, I.; Sasada, T.; Fukutake, M.; Asano, M.; Ueno, Y.; Nagasaki, Y.; Shirakawa, O.; Hishimoto, A. Association of FKBP5 gene haplotypes with completed suicide in the Japanese population. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 252–256. [Google Scholar] [CrossRef]
- Leszczyńska-Rodziewicz, A.; Szczepankiewicz, A.; Skibińska, M.; Pawlak, J.; Dmitrzak-Węglarz, M.; Hauser, J.; Narozna, B. Possible association between haplotypes of the FKBP5 gene and suicidal bipolar disorder, but not with melancholic depression and psychotic features, in the course of bipolar disorder. Neuropsychiatr. Dis. Treat. 2014, 10, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Breen, M.E.; Seifuddin, F.; Zandi, P.P.; Potash, J.B.; Willour, V.L. Investigating the role of early childhood abuse and HPA axis genes in suicide attempters with bipolar disorder. Psychiatr. Genet. 2015, 25, 106–111. [Google Scholar] [CrossRef] [Green Version]
- Fudalej, S.; Kopera, M.; Wołyńczyk-Gmaj, D.; Fudalej, M.; Krajewski, P.; Wasilewska, K.; Szymański, K.; Chojnicka, I.; Podgórska, A.; Wojnar, M.; et al. Association between FKBP5 Functional Polymorphisms and Completed Suicide. Neuropsychobiology 2015, 72, 126–131. [Google Scholar] [CrossRef]
- Breen, M.E.; Gaynor, S.C.; Monson, E.T.; De Klerk, K.; Parsons, M.G.; Braun, T.A.; DeLuca, A.P.; Zandi, P.P.; Potash, J.B.; Willour, V.L. Targeted Sequencing of FKBP5 in Suicide Attempters with Bipolar Disorder. PLoS ONE 2016, 11, e0169158. [Google Scholar] [CrossRef]
- Yin, H.; Galfalvy, H.; Pantazatos, S.P.; Huang, Y.-Y.; Rosoklija, G.B.; Dwork, A.J.; Burke, A.; Arango, V.; Oquendo, M.A.; Mann, J.J. Glucocorticoid Receptor-Related Genes: Genotype and Brain Gene Expression Relationships to Suicide and Major Depressive Disorder. Depress. Anxiety 2016, 33, 531–540. [Google Scholar] [CrossRef] [Green Version]
- Mirkovic, B.; Cohen, D.; Laurent, C.; Lasfar, M.; Marguet, C.; Gerardin, P. A case-control association study of 12 candidate genes and attempted suicide in French adolescents. Int. J. Adolesc. Med. Health 2017, 32. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, X.-Z.; Benedek, D.M.; Fullerton, C.S.; Forsten, R.D.; Naifeh, J.; Li, X.; Wynn, G.; Russell, D.W.; Ursano, R.J. Genetic predictor of current suicidal ideation in US service members deployed to Iraq and Afghanistan. J. Psychiatr. Res. 2019, 113, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Nobile, B.; Ramoz, N.; Jaussent, I.; Dubois, J.; Guillaume, S.; Gorwood, P.; Courtet, P. Polymorphisms of stress pathway genes and emergence of suicidal ideation at antidepressant treatment onset. Transl. Psychiatry 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wasserman, D.; Sokolowski, M.; Rozanov, V.; Wasserman, J. The CRHR1 gene: A marker for suicidality in depressed males exposed to low stress. Genes Brain Behav. 2007, 7, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Ben-Efraim, Y.J.; Wasserman, D.; Sokolowski, M. Gene-environment interactions between CRHR1 variants and physical assault in suicide attempts. Genes Brain Behav. 2011, 10, 663–672. [Google Scholar] [CrossRef]
- Guillaume, S.; Perroud, N.; Jollant, F.; Jaussent, I.; Olié, E.; Malafosse, A.; Courtet, P. HPA axis genes may modulate the effect of childhood adversities on decision-making in suicide attempters. J. Psychiatr. Res. 2013, 47, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Leszczynska-Rodziewicz, A.; Maciukiewicz, M.; Szczepankiewicz, A.; Pogłodziński, A.; Hauser, J. Association between OPCRIT dimensions and polymorphisms of HPA axis genes in bipolar disorder. J. Affect. Disord. 2013, 151, 744–747. [Google Scholar] [CrossRef]
- Pawlak, J.; Dmitrzak-Weglarz, M.; Wiłkość-Dębczyńska, M.; Szczepankiewicz, A.; Leszczynska-Rodziewicz, A.; Zaremba, D.; Kapelski, P.; Rajewska-Rager, A.; Hauser, J. Suicide behavior as a quantitative trait and its genetic background. J. Affect. Disord. 2016, 206, 241–250. [Google Scholar] [CrossRef]
- Bastos, C.R.; Gazal, M.; Quevedo, L.D.A.; Costa, J.L.; Wiener, C.D.; Jansen, K.; de Mola, C.L.; Oses, J.P.; Souza, L.D.; Portela, L.V.; et al. Polymorphism in CRHR1 gene affects the IL-1β levels in suicidal attempters. J. Psychiatr. Res. 2017, 86, 34–38. [Google Scholar] [CrossRef]
- Ludwig, B.; Kienesberger, K.; Carlberg, L.; Swoboda, P.; Bernegger, A.; Koller, R.; Wang, Q.; Inaner, M.; Zotter, M.; Kapusta, N.D.; et al. Influence of CRHR1 Polymorphisms and Childhood Abuse on Suicide Attempts in Affective Disorders: A GxE Approach. Front. Psychiatry 2018, 9, 165. [Google Scholar] [CrossRef] [Green Version]
- Sadeh, N.; Wolf, E.; Logue, M.W.; Hayes, J.P.; Stone, A.; Griffin, L.M.; Schichman, S.A.; Miller, M. Epigenetic Variation at Ska2 Predicts Suicide Phenotypes and Internalizing Psychopathology. Depress. Anxiety 2016, 33, 308–315. [Google Scholar] [CrossRef] [Green Version]
- McGowan, O.P.; Sasaki, A.; D’Alessio, A.C.; Dymov, S.; Labonté, B.; Szyf, M.; Turecki, G.; Meaney, M.J. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat. Neurosci. 2009, 12, 342–348. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, D.; Fullerton, J.; Webster, M.J.; Weickert, C.S. Glucocorticoid Receptor 1B and 1C mRNA Transcript Alterations in Schizophrenia and Bipolar Disorder, and Their Possible Regulation by GR Gene Variants. PLoS ONE 2012, 7, e31720. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Qi, X.-R.; Gao, S.-F.; Lu, J.; van Wamelen, D.; Kamphuis, W.; Bao, A.-M.; Swaab, D. Different stress-related gene expression in depression and suicide. J. Psychiatr. Res. 2015, 68, 176–185. [Google Scholar] [CrossRef]
- Roy, B.; Shelton, R.C.; Dwivedi, Y. DNA methylation and expression of stress related genes in PBMC of MDD patients with and without serious suicidal ideation. J. Psychiatr. Res. 2017, 89, 115–124. [Google Scholar] [CrossRef]
- Melhem, N.M.; Munroe, S.; Marsland, A.; Gray, K.; Brent, D.; Porta, G.; Douaihy, A.; Laudenslager, M.; DePietro, F.; Diler, R.S.; et al. Blunted HPA axis activity prior to suicide attempt and increased inflammation in attempters. Psychoneuroendocrinology 2017, 77, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.B.; Munroe, S.; Gray, K.; Porta, G.; Douaihy, A.; Marsland, A.; Brent, D.; Melhem, N.M. The role of substance use, smoking, and inflammation in risk for suicidal behavior. J. Affect. Disord. 2018, 243, 33–41. [Google Scholar] [CrossRef]
- Hiroi, N.; Wong, M.-L.; Licinio, J.; Park, C.; Young, M.; Gold, P.W.; Chrousos, G.P.; Bornstein, S.R. Expression of corticotropin releasing hormone receptors type I and type II mRNA in suicide victims and controls. Mol. Psychiatry 2001, 6, 540–546. [Google Scholar] [CrossRef] [Green Version]
- Merali, Z.; Du, L.; Hrdina, P.; Palkovits, M.; Faludi, G.; Poulter, M.O.; Anisman, H. Dysregulation in the Suicide Brain: mRNA Expression of Corticotropin-Releasing Hormone Receptors and GABAA Receptor Subunits in Frontal Cortical Brain Region. J. Neurosci. 2004, 24, 1478–1485. [Google Scholar] [CrossRef] [Green Version]
- Thomas, N.; Armstrong, C.W.; Hudaib, A.R.; Kulkarni, J.; Gurvich, C. A network meta-analysis of stress mediators in suicide behaviour. Front. Neuroendocrinol. 2021, 63, 100946. [Google Scholar] [CrossRef]
- Zheng, D.; Bi, X.; Zhang, T.; Han, C.; Ma, T.; Wang, L.; Sun, M.; Cui, K.; Yang, L.; Liu, L. Epigenetic Alterations of the Promoter Region of the POMC Gene in Adolescent Depressive Disorder Patients with Nonsuicidal Self-Injury Behaviors. Psychol. Res. Behav. Manag. 2020, 13, 997–1008. [Google Scholar] [CrossRef]
- Groh, A.; Rhein, M.; Buchholz, V.; Burkert, A.; Huber, C.G.; Lang, U.E.; Borgwardt, S.; Heberlein, A.; Muschler, M.A.; Hillemacher, T.; et al. Epigenetic Effects of Intravenous Diacetylmorphine on the Methylation of POMC and NR3C1. Neuropsychobiology 2017, 75, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.N.; Rizavi, H.S.; Ren, X.; Dwivedi, Y.; Palkovits, M. Region-specific alterations in glucocorticoid receptor expression in the postmortem brain of teenage suicide victims. Psychoneuroendocrinology 2013, 38, 2628–2639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braquehais, M.D.; Picouto, M.D.; Casas, M.; Sher, L. Hypothalamic-pituitary-adrenal axis dysfunction as a neurobiological correlate of emotion dysregulation in adolescent suicide. World J. Pediatr. 2012, 8, 197–206. [Google Scholar] [CrossRef] [PubMed]
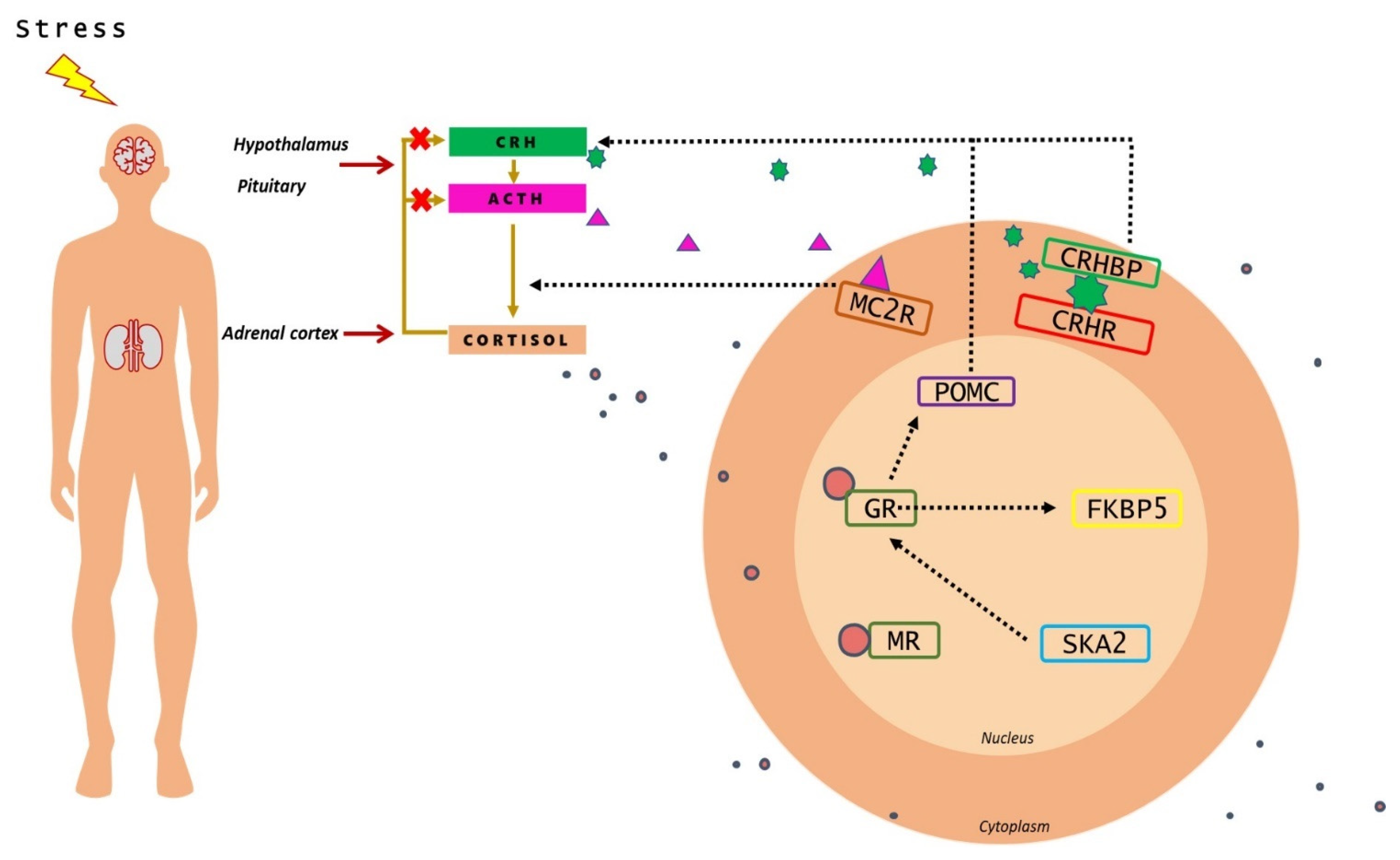


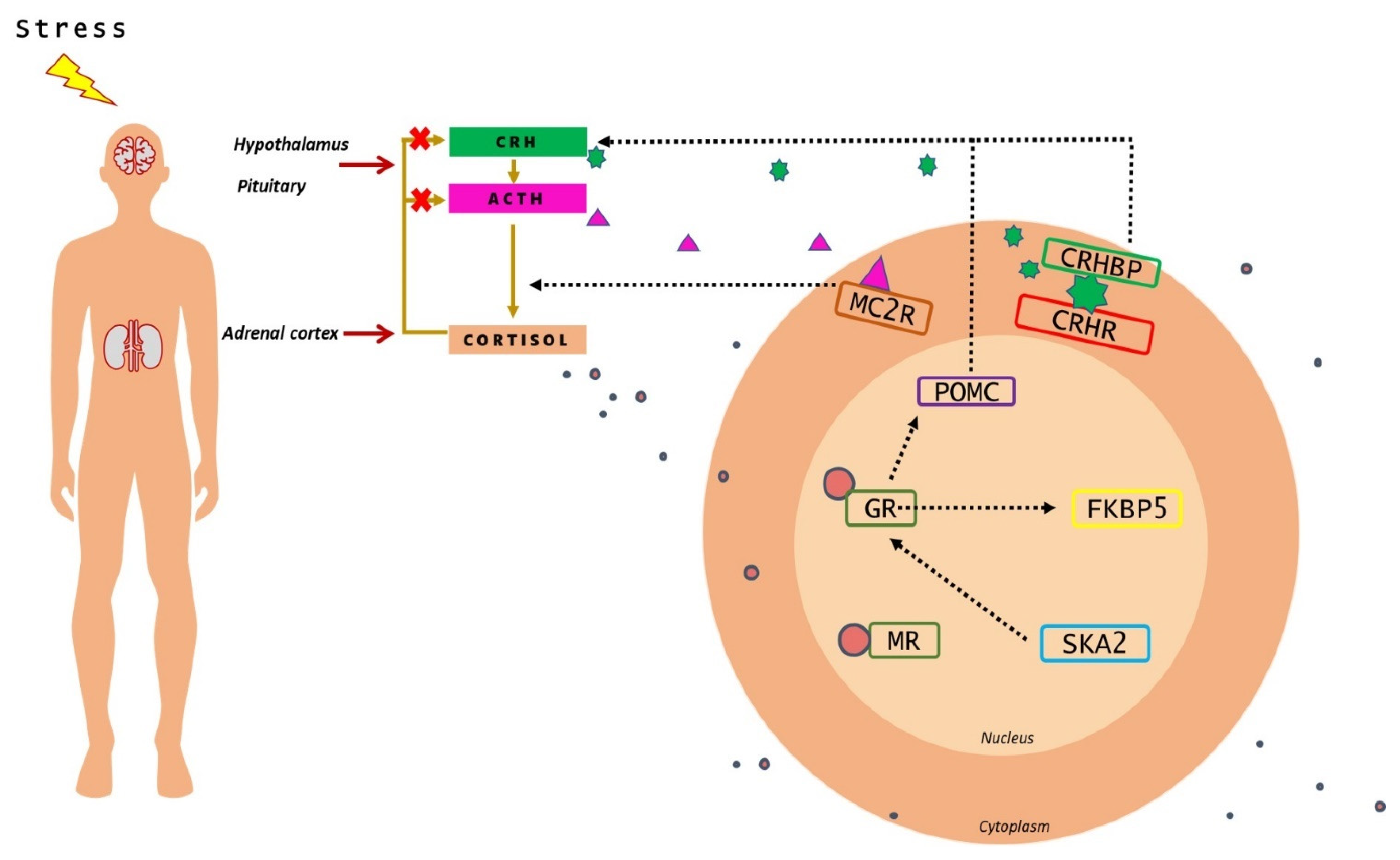{kind=link}
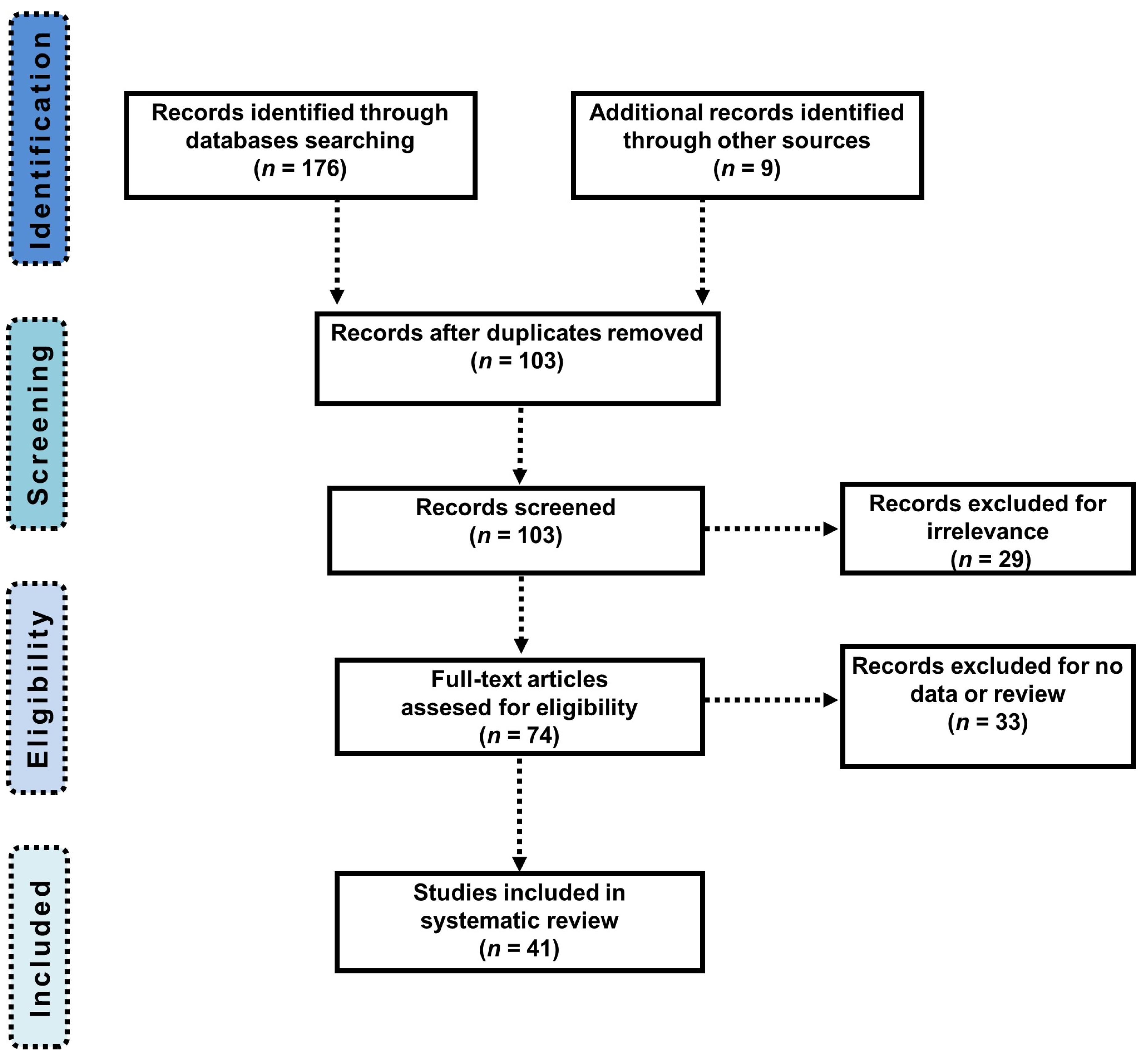{kind=link}
| Author, Year | Chromosome | SNP | Location | Substitution | Diagnostic | Suicide Behavior | Population Data (N) | Measurement on Trauma Exposure | Tissue Source | Main Outcomes | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cases | Control | ||||||||||
| FKBP5 | |||||||||||
| Papiol, S. 2007 [31] | 6 | rs1360780 | Intron | T > C | MDD | SA | 24 | 96 | - | Blood | T allele carriers showed 2.10 increased for non-responding to citalopram treatment at week 4 |
| Willour, V. L. 2009 [32] | 6 | rs1043805 | 3’UTR | T > A | BD | SA | 544 | - | - | - | Four SNPs showed significant associations with SA: rs1043805, rs3800373, rs9296158 and rs1360780. |
| rs3800373 | 3′UTR | C > A | |||||||||
| rs7757037 | Intron | G > A | |||||||||
| rs3798346 | Intron | A > G | |||||||||
| rs9296158 | Intron | G > A | |||||||||
| rs1360780 | Intron | T > A | |||||||||
| rs4713902 | Intron | T > C | |||||||||
| rs6912833 | Intron | A > T | |||||||||
| rs9380525 | Intron | G > A | |||||||||
| Brent, D. 2010 [33] | 6 | rs3800373 | 3′UTR | C > A | MDD | SB | 18 | - | - | Blood and Buccal | Genotypes of rs1360780TT and rs3800373GG were associated with SB, even after controlling for treatment effects and relevant covariates. |
| rs1360780 | Intron | T > A | |||||||||
| Roy, A. 2010 [34] | 6 | rs3800373 | 3′UTR | C > A | SUD | SA | 248 | 1465 | CTQ | Blood | 1. Three SNPs showed significant associations with SA: rs3777747, rs4713902, and rs9470080. 2. Three SNPs showed a G X E interaction: rs3800373, rs9296158, and 1360780. 3. There were no interactive effects between substance dependence, CTQ scores, and FKBP5 SNPs in SA. |
| rs9470065 | Intron | G > A | |||||||||
| rs9296158 | Intron | G > A | |||||||||
| rs4713899 | Intron | G > A | |||||||||
| rs9470067 | Intron | G > A | |||||||||
| rs3777747 | Intron | G > A | |||||||||
| rs7762668 | Intron | G > A | |||||||||
| rs9462099 | Intron | T > C | |||||||||
| rs9380524 | Intron | C > A | |||||||||
| rs1360780 | Intron | T > C | |||||||||
| rs7771722 | Intron | G > A | |||||||||
| rs4713902 | Intron | T > C | |||||||||
| rs9462100 | Intron | T > C | |||||||||
| rs2092427 | Intron | G > A | |||||||||
| rs7762760 | Intron | G > A | |||||||||
| rs9470080 | Intron | T > C | |||||||||
| Perroud, N. 2011 [35] | 6 | rs1360780 | Intron | T > C | DE | SI | 131 | - | - | - | The T allele was a risk factor of SI and it was associated with the response to antidepressant treatment. |
| Supriyanto, I. 2011 [36] | 6 | rs3800373 | 3′UTR | C > A | SV | CS | 219 | 228 | - | Blood | No association. |
| rs1360780 | Intron | T > C | |||||||||
| rs2395635 | Intron | A > C | |||||||||
| Roy, A. 2012 [28] | 6 | rs3800373 | 3′UTR | C > A | SUD | SA | 141 | 689 | CTQ | Blood | 1. In the group exposed to severe trauma, the prevalence of SA was 0.49 in carriers of the major homozygote. 2. An analysis of the interaction of total CTQ score with combined FKBP5 rs3800373 and CRHBP rs7728378 genotypes was significant. |
| Leszczynska-Rodziewicz, A. 2014 [37] | 6 | rs1360780 | Intron | T > C | BD | SA | 156 | 724 | - | Blood | No association. |
| rs755658 | Intron | C > T | |||||||||
| rs4713916 | Intron | A > G | |||||||||
| rs7748266 | Intron | T > C | |||||||||
| rs9296158 | Intron | G >A | |||||||||
| rs9394309 | Intron | G >A | |||||||||
| rs9470080 | Intron | T > A | |||||||||
| rs3800373 | 3′UTR | C > A | |||||||||
| Breen, M. E. 2015 [38] | 6 | rs6926133 | Intron | A > C | BD | SA | 631 | 657 | ELES | - | No association. |
| rs12200498 | Intron | G > A | |||||||||
| rs9380526 | Intron | C > T | |||||||||
| rs16879378 | Intron | A > C | |||||||||
| rs4713899 | Intron | G > A | |||||||||
| Fudalej, S. 2015 [39] | 6 | rs3800373 | 3′UTR | C > A | SV | CS | 520 | 475 | - | Blood | A significant association between the high-induction rs3800373 C allele and SV was detected. |
| rs1360780 | Intron | T > C | |||||||||
| Breen, M. E. 2016 [40] | 6 | rs141713011 | Intron/3′UTR | G > T | BD or SD | SA | 476 | 473 | - | - | rs141713011 showed an excess of minor alleles in SA that was statistically significant following correction for multiple testing, but it could not be replicated. |
| rs140664762 | Intron/3′UTR | G > A | |||||||||
| rs575259136 | Intron | A > AAAG | |||||||||
| rs13192954 | Intron | A > G | |||||||||
| rs553156199 | 3′UTR | C > T | |||||||||
| Yin, H. 2016 [41] | 6 | rs3800373 | 3′UTR | C > A | MDD | SA and CS | SA: 87 CS: 121 | SA: 190 CS: 88 | - | SA: - SV: Brain | 1. rs9296158, rs3777747, rs4713902, rs7757037, rs737054, and rs9380529 showed evidence of association at uncorrected p < 0.05 level with SA 2. There was no evidence of an association between these SNPs and death by suicide in the postmortem sample. |
| rs9296158 | Intron | A > C | |||||||||
| rs3777747 | Intron | A >G | |||||||||
| rs4713902 | Intron | T > C | |||||||||
| rs9470080 | Intron | T > A | |||||||||
| rs7757037 | Intron | G > A | |||||||||
| rs737054 | Intron | G > A | |||||||||
| rs9380529 | Intron | A > C | |||||||||
| Mirkovic, B. 2017 [42] | 6 | rs1360780 | Intron | T > C | Mixed | SA | 98 | 150 | - | Saliva | No association. |
| rs3800373 | 3′UTR | C > A | |||||||||
| Segura, A. G. 2019 [19] | 6 | rs3777747 | Intron | A >G | BD | SB | 129 | - | CTQ | Blood | 1. rs3777747AA and rs2766533GG genotypes were associated with SB. 2. Did not find an interaction between any CTQ scores and SNPs. |
| rs1360780 | Intron | T >A | |||||||||
| rs17542466 | Intron | A > C | |||||||||
| rs2766533 | Intron | G > A | |||||||||
| Zhang, L. 2019 [43] | 6 | rs1360780 | Intron | T > C | PTSD and DE | SI | 266 | 3623 | LEC | Saliva | No association. |
| rs9470080 | Intron | T > A | |||||||||
| rs3800373 | 3′UTR | C > A | |||||||||
| rs9296158 | Intron | A > C | |||||||||
| Nobile, B. 2020 [44] | 6 | rs737054 | Intron | G > A | MDD | SI and SA | SI: 99SA:9 | 384 | - | Buccal | The TT genotype of rs737054 and TT genotype of rs6902321 were significantly associated with SI. These associations were not significant after multiple test corrections. |
| rs6902321 | Intron | C > T | |||||||||
| rs3800373 | 3′UTR | C > A | |||||||||
| rs7757037 | Intron | G > A | |||||||||
| rs1360780 | Intron | T > C | |||||||||
| rs9470080 | Intron | T > A | |||||||||
| Berent, D. 2020 [21] | 6 | rs1360780 | Intron | T > C | AD | SA | 176 | 127 | ACE | Buccal | No association. |
| Hernández-Díaz, Y. 2021 [22] | 6 | rs4713916 | Intron | A > G | SA | SA | 146 | 277 | - | Blood | 1. rs1360780 T minor allele was found to be a risk factor for SA. 2. rs3800373 C minor allele was found to be a protective factor for SA. |
| rs1360780 | Intron | T > C | |||||||||
| rs4713902 | Intron | T > C | |||||||||
| rs3800373 | 3′UTR | C > A | |||||||||
| rs9296158 | Intron | A > C | |||||||||
| CRH | |||||||||||
| Wasserman, D. 2008 [45] | 8 | rs1870393 | Intron | A > C | Mixed | SA | 542 | - | SLEI | Blood | No association. |
| rs3176921 | 5′ region | C > T | |||||||||
| De Luca, V. 2010 [27] | 8 | rs3176921 | 5′ region | C > T | SZ | SA | 81 | 150 | - | Blood | No association. |
| Roy, A. 2012 [28] | 8 | rs6996265 | Intron | A > G | SUD | SA | 141 | 689 | CTQ | Blood | No association. |
| rs3176921 | 5′ region | C > T | |||||||||
| rs6472257 | 5′ region | C > T | |||||||||
| rs5030875 | Intergenic | T > G | |||||||||
| Breen, M. E. 2015 [38] | 8 | rs6990486 | Downstream | G > A | BD | SA | 631 | 657 | ELES | - | No association. |
| rs6472257 | Upstream | C > T | |||||||||
| rs7835214 | Downstream | T > C | |||||||||
| rs10957368 | Downstream | T > C | |||||||||
| rs10105164 | Downstream | C > T | |||||||||
| CRHBP | |||||||||||
| Papiol, S. 2007 [31] | 5 | rs7728378 | Intron | C > T | MDD | SA | 24 | 96 | - | Blood | No association. |
| rs1875999 | 3′UTR | A > G | |||||||||
| De Luca, V. 2010 [27] | 5 | rs1875999 | 3′UTR | A > G | SZ | SA | 81 | 150 | - | Blood | The heterozygous genotype was significantly associated with SA as a risk of attempt. |
| Roy, A. 2012 [28] | 5 | rs3792738 | 5′UTR | C > A | SUD | SA | 141 | 689 | CTQ | Blood | rs6453267, rs7728378, and rs10474485 showed a nominally significant interaction with the continuous CTQ score to predict SA. 2. There was an additive effect of FKBP5 rs3800373 and CRHBP rs7728378 in the group exposed to severe trauma. |
| rs328967 | Intron | A > G | |||||||||
| rs6453267 | Intron | G > A | |||||||||
| rs7728378 | Intron | C > T | |||||||||
| rs1875999 | 3′UTR | A > G | |||||||||
| rs10474485 | Intron | C > A | |||||||||
| rs1715747 | Intron | C > T | |||||||||
| rs1500 | Alt isoform | C > G | |||||||||
| Breen, M. E. 2015 [38] | 5 | rs7721799 | Intron | G > A | BD | SA | 631 | 657 | ELES | - | No association. |
| rs2174444 | Downstream | C > T | |||||||||
| rs10473984 | Downstream | G > T | |||||||||
| Segura, A. G. 2019 [19] | 5 | rs7728378 | Intron | C > T | BD | SB | 139 | - | CTQ | Blood | 1. rs7728378-C carriers were associated with SB. This association did not remain significant after correcting for multiple comparisons. 2. Did not find an interaction between any CTQ scores and SNPs. |
| rs10474485 | Intron | C > A | |||||||||
| CRHR1 | |||||||||||
| Papiol, S. 2007 [31] | 17 | rs110402 | Intron | C > T | MDD | SA | 24 | 96 | - | Blood | TT homozygous had nearly 3 times more risk to develop seasonal pattern episodes. |
| rs242937 | Intron | A > C | |||||||||
| Wasserman, D. 2008 [45] | 17 | rs1396862 | Intron | G > A | Mixed | SA | 542 | - | SLEI | Blood | Stratification based on the levels of lifetime stress showed reproducible association and linkage of rs4792887 to SA exposed to low levels of stress mainly in males who were depressed. |
| rs4792887 | Intron | C > T | |||||||||
| Wasserman, D. 2009 [24] | 17 | rs4792887 | Intron | C > T | Mixed | SA | 672 | - | SLEI | Blood | 1. The minor T-allele of rs12936511 was significantly transmitted in males with SB and with increased BDI scores. 2. Association and linkage with increased BDI scores among suicidal males with an additional SNP, located proximally to the index SNP rs4792887, as well as with two distal SNPs, which were correlated with index SNP rs4792887. |
| rs110402 | Intron | C > T | |||||||||
| rs12936511 | Exon | C > T | |||||||||
| rs242939 | Intron | A > G | |||||||||
| rs242938 | Intron | A > C | |||||||||
| rs1876831 | Intron | C > T | |||||||||
| rs16940665 | Exon | T > C | |||||||||
| rs4792887 | Intron | C > T | |||||||||
| rs110402 | Intron | C > T | |||||||||
| De Luca, V. 2010 [27] | 17 | rs16940665 | Exon | T > C | SZ | SA | 81 | 150 | - | Blood | No association. |
| Ben-Efraim, Y. J. 2011 [46] | 17 | rs4792887 | Intron | C > A | DE | SA | 284 | 354 | SLEI | - | 1. G×E predominantly in females with SA between rs7209436 and childhood/adolescence physical assault or attack. 2. Male-specific G×E between rs16940665 and physical assault or attack exposure in adulthood. 3. Male-specific G×E in depressed SA, rs4792887, and cumulative stressful life events. |
| rs110402 | Intron | C > T | |||||||||
| rs16940665 | Exon | T > C | |||||||||
| rs4792887 | Intron | C > T | |||||||||
| Roy, A. 2012 [28] | 17 | rs9900679 | Intron | A > C | SUD | SA | 141 | 689 | CTQ | Blood | No association. |
| rs4792887 | Intron | C > T | |||||||||
| rs110402 | Intron | C > T | |||||||||
| rs249224 | Intron | C > A | |||||||||
| rs8072451 | Intron | C > T | |||||||||
| rs81189 | Intron | G > C | |||||||||
| rs24939 | Intron | A > G | |||||||||
| rs173365 | Intron | T > C | |||||||||
| rs17689918 | Intron | G > A | |||||||||
| Guillaume, S. 2013 [47] | 17 | rs242948 | Downstream | C > T | Mixed | SA | 218 | - | CTQ | Blood | Sexual abuse and emotional neglect in childhood interacted with rs1396862, rs878886, and rs242948 to modulate adult decision making in SA. |
| rs1396862 | Intron | G > A | |||||||||
| rs878886 | 3′UTR | G > T | |||||||||
| rs4076452 | Intron | G > C | |||||||||
| Leszczynska-Rodziewicz, A. 2013 [48] | 17 | rs4076452 | Intron | G > C | BD | SA | 225 | 712 | - | Blood | No association. |
| rs12936511 | Exon | C > T | |||||||||
| rs4792887 | Intron | C > T | |||||||||
| rs24290 | Intron | T > C | |||||||||
| rs878886 | 3′ UTR | G > T | |||||||||
| rs173365 | Intron | T > C | |||||||||
| rs110402 | Intron | C > T | |||||||||
| Breen, M. E. 2015 [38] | 17 | rs2664008 | Intron | G > A | BD | SA | 631 | 657 | ELES | - | Significant interaction between rs2664008 and a history of childhood physical and/or sexual abuse was reported; however, this interaction was not significant after correcting for multiple testing. |
| rs1724425 | Intron | C > T | |||||||||
| rs1526123 | Intron | T > A | |||||||||
| rs6593447 | Intron | A > G | |||||||||
| rs11655764 | Intron | G > A | |||||||||
| Pawlak, J. 2016 [49] | 17 | rs4792877 | Intron | A > G | AD | SA | 277 | 847 | - | Blood | rs16940665 polymorphism was associated with SA in MDD males. |
| rs12936511 | Exon | C > T | |||||||||
| rs110402 | Intron | C > T | |||||||||
| rs16940665 | Exon | T > C | |||||||||
| Mirkovic, B. 2017 [42] | 17 | rs4792887 | Intron | C > T | Mixed | SA | 98 | 150 | - | Buccal | No association. |
| Bastos, C. R. 2017 [50] | 17 | rs110402 | Intron | C > T | Mixed | SI and SA | SI: 15SA: 20 | 136 | - | Blood | Individuals who carried the A allele increased in 15% additional risk for SA via the increase in IL-1b levels. |
| Ludwig, B. 2018 [51] | 17 | rs7209436 | Intron | C > T | AD | SA | 70 | 181 | CTQ and BLEQ | Blood | Significant gene-environment-interactions were found for the SNPs rs7209436 and rs110402, reflecting the impact of childhood trauma and CRHR1 gene polymorphisms in previous SA. |
| rs4792887 | Intron | C > T | |||||||||
| rs110402 | Intron | C > T | |||||||||
| rs242924 | Intron | C > A | |||||||||
| rs242939 | Intron | A > G | |||||||||
| Segura, A. G. 2019 [19] | 17 | rs110402 | Intron | C > T | BD | SB | 129 | - | CTQ | Blood | No association. |
| rs242940 | Intron | A > G | |||||||||
| Sanabrais-Jiménez, M.A. 2019 [23] | 17 | rs110402 | Intron | C > T | BD and MDD | SA | 183 | 183 | CTQ | Blood | The analysis showed an interaction of CRHR1 and CRHR2 genes with childhood trauma, thus conferring increased risk of having presented at least one SA. |
| rs242924 | Intron | C > A | |||||||||
| rs16940665 | Exon | T > C | |||||||||
| Nobile, B. 2020 [44] | 17 | rs878886 | 3′UTR | G > T | MDD | SI and SA | SI: 99SA: 9 | 384 | - | Buccal | No association. |
| CRHR2 | |||||||||||
| Papiol, S. 2007 [31] | 7 | rs2240403 | Exon | C > T | MDD | SA | 24 | 96 | - | Blood | Allele G carriers of rs2270007 showed a worse overall response to citalopram though time of follow-up and showed 2.93 increased risk for nonresponding to citalopram treatment at week 4. |
| rs2270007 | Intron | G > C | |||||||||
| De Luca, V. 2010 [27] | 7 | rs1076292 | Intron | C > T | SZ | SA | 81 | 150 | Blood | No association | No association. |
| Roy, A. 2012 [28] | 7 | rs3779250 | Intron | G > A | SUD | SA | 141 | 689 | CTQ | Blood | No association. |
| rs973002 | Intron | A > G | |||||||||
| rs8192498 | - | G > A | |||||||||
| rs2190242 | Intron | A > C | |||||||||
| rs2284217 | Intron | G > A | |||||||||
| rs2014663 | Intron | A > G | |||||||||
| rs6967702 | 5′ region | G > C | |||||||||
| rs4723002 | Intergenic | A > G | |||||||||
| rs255102 | Intergenic | T > A | |||||||||
| rs255105 | Intergenic | T > C | |||||||||
| rs255125 | Intergenic | G > A | |||||||||
| Guillaume, S. 2013 [47] | 7 | rs255098 | Intron | G > A | Mixed | SA | 218 | - | CTQ | Blood | Sexual abuse and emotional neglect in childhood interacted with rs255098 to modulate adult decision making in SA. |
| rs2270007 | Intron | G > C | |||||||||
| Breen, M. E. 2015 [38] | 7 | rs2267716 | Intron | T > C | BD | SA | 631 | 657 | ELES | - | No association. |
| rs11980048 | Intron | G > T | |||||||||
| rs4723002 | Intron | A > G | |||||||||
| rs2190242 | Intron | C > A | |||||||||
| rs4723003 | Intron | C > T | |||||||||
| Segura, A. G. 2019 [19] | 7 | rs4722999 | Intron | C > T | BD | SB | 129 | - | CTQ | Blood | No association. |
| rs2284219 | Intron | A > G | |||||||||
| rs255115 | Intron | G > A | |||||||||
| rs255102 | Intergenic | T > A | |||||||||
| Sanabrais-Jiménez, M.A. 2019 [23] | 7 | rs2190242 | Intron | C > A | BD and MDD | SA | 183 | 183 | CTQ | Blood | An interaction of CRHR1 and CRHR2 genes with childhood trauma, thus conferring an increased risk of having presented at least one SA. |
| rs2284217 | Intron | G > A | |||||||||
| rs2014663 | Intron | A > G | |||||||||
| NR3C1 | |||||||||||
| De Luca, V. 2010 [27] | 5 | rs6196 | Exon | A > G | SZ | SA | 81 | 150 | - | Blood | This SNP was significantly associated with SA, positively protecting against suicide attempt. |
| Supriyanto, I. 2011 [36] | 5 | rs6196 | Exon | A > G | - | CS | 219 | 228 | - | Blood | No association. |
| rs10052957 | Intron | G > A | |||||||||
| Leszczynska-Rodziewicz, A. 2013 [48] | 5 | rs41423247 | Intron | G > C | BD | SA | 225 | 712 | - | Blood | No association. |
| rs6195 | Intron | T > C | |||||||||
| rs6198 | 3′UTR | T > C | |||||||||
| rs6191 | 3′UTR | C > A | |||||||||
| rs6196 | Exon | A > G | |||||||||
| rs33388 | Intron | A > G | |||||||||
| Breen, M. E. 2015 [38] | 5 | rs4912905 | Intron | G > C | BD | SA | 631 | 657 | ELES | - | No association. |
| rs10042042 | Intron | G > A | |||||||||
| rs17209251 | Intron | A > G | |||||||||
| rs17100236 | Intron | T > C | |||||||||
| rs10477211 | Intron | A > G | |||||||||
| Yin, H. 2016 [41] | 5 | rs6196 | Exon | A > G | MDD | SA and CS | SA: 87 | 190 | - | SA: - | rs9324924 showed evidence of association at uncorrected p < 0.05 level with SA. |
| rs33388 | Intron | A > C | |||||||||
| rs33389 | Intron | C > T | SV: 121 | 88 | SV: Brain | ||||||
| rs10052957 | Intron | G > A | |||||||||
| rs9324924 | Intron | G > A | |||||||||
| Park, S. 2016 [20] | 5 | rs41423247 | Intron | G > C | Cancer | CS | 182 | 161 | - | SNP was associated with the susceptibility to suicide within the first year after cancer diagnosis. | |
| Segura, A. G. 2019 [19] | 5 | rs6198 | 3′UTR | T > C | BD | SB | 129 | - | CTQ | Blood | No association. |
| rs2963156 | Intron | T > C | |||||||||
| rs1837262 | Intron | C > A | |||||||||
| rs4912910 | Intron | A > G | |||||||||
| rs4634384 | Intron | C > T | |||||||||
| Nobile, B. 2020 [44] | 5 | rs2963155 | Intron | A > G | MDD | SI and SA | SI: 99 | 384 | - | Buccal | AG genotype of rs2963155 was associated with SI. This association was not significant after multiple test correction. |
| rs33388 | Intron | A > C | |||||||||
| rs4912905 | Intron | G > C | |||||||||
| rs41423247 | Intron | G > C | SA: 9 | ||||||||
| rs6189 | Exon | C > T | |||||||||
| rs12656106 | Intron | G > C | |||||||||
| rs4607376 | Intron | G > T | |||||||||
| NR3C2 | |||||||||||
| Supriyanto, I. 2011 [36] | 4 | rs5525 | Exon | A > C | CS | CS | 219 | 228 | - | Blood | No association. |
| rs5522 | Exon | C > T | |||||||||
| rs2070951 | 5′UTR | G > A | |||||||||
| Segura, A. G. 2019 [37] | 4 | rs5534 | 3′UTR | T > C | BP | SB | 129 | - | CTQ | Blood | No association. |
| rs12499208 | Intron | T > C | |||||||||
| rs6846591 | Intron | T > C | |||||||||
| rs5522 | Exon | C > T | |||||||||
| SKA2 | |||||||||||
| Kaminsky, Z. 2015 [25] | 17 | rs7208505 | 3′UTR | G > T | PTSD | SI and SA | SI: 325 SA: 746 | 658 | CTQ | Blood and Saliva | Significant interactions of SKA2 3′-UTR DNA methylation and rs7208505 genotype for SI and SA. |
| Yin, H. 2016 [41] | 17 | rs8082544 | - | A > G | MDD | SA and CS | SA: 87SV: 121 | 190 88 | - | SA: - SV: Brain | 1. rs12945875, rs9911583, and rs8067682 showed evidence for association at uncorrected p < 0.05 level with SA. 2. rs8082544 and rs7502947 showed association with death by suicide. |
| rs12945875 | Intron | G > A | |||||||||
| rs9911583 | Intron | G > A | |||||||||
| rs8067682 | Intron | A > G | |||||||||
| rs7502947 | Intron | G > A | |||||||||
| Sadeh, N. 2016 [52] | 17 | rs7208505 | 3′UTR | G > T | PTSD | SI, SP, SA | SI: 146 SA: 50 SP:92 | - | - | Blood | No association. |
| Nobile, B. 2020 [44] | 17 | rs7208505 | 3′UTR | G > T | MDD | SI and SA | SI: 99 SA:9 | 384 | - | Buccal | GG/AG genotype was significantly associated with SI. This association was not significant after multiple test correction. |
| MC2R | |||||||||||
| De Luca, V. 2010 [27] | 18 | rs4797825 | 3′UTR | C > T | SZ | SA | 81 | 150 | - | Blood | No association. |
| Breen, M. E. 2015 [38] | 18 | rs3744819 | 3′UTR | C > A | BD | SA | 631 | 657 | ELES | - | No association. |
| rs12456733 | Intron | G > A | |||||||||
| rs1941088 | Intron | G > A | |||||||||
| rs3888305 | 3′UTR | A > C | |||||||||
| rs4308014 | 3′UTR | C > T | |||||||||
| Segura, A. G. 2019 [37] | 18 | rs4797825 | 3′UTR | C > T | BD | SB | 129 | - | CTQ | Blood | No association. |
| rs9961110 | Intron | T > C | |||||||||
| rs17624314 | Intron | A > G | |||||||||
| POMC | |||||||||||
| Breen, M. E. 2015 [38] | 2 | rs7565877 | intron | A > G | BD | SA | 631 | 657 | ELES | - | No association. |
| rs6545975 | intron | C > A | |||||||||
| rs7565427 | intron | A > C | |||||||||
| rs934778 | intron | A > G | |||||||||
| rs1866146 | Downstream | G > A | |||||||||
| Segura, A. G. 2019 [19] | 2 | rs713586 | Intron | T > C | BD | SB | 129 | - | CTQ | Blood | No association. |
| rs6713532 | Intron | T > C | |||||||||
| rs6545975 | Intron | C > A | |||||||||
| rs934778 | Intron | A > G | |||||||||
| Author, Year | Suicide Behavior | N | Ethinicity | Tissue Source | Expression | Variant | Genotype/Expression | Trauma Exposure | Cortisol Concentrations |
|---|---|---|---|---|---|---|---|---|---|
| NR3C1 | |||||||||
| McGowan, P. 2009 [53] | CS | CS: 24 Controls:12 | Caucasian | Brain | ↓ | - | - | mRNA was significantly reduced in SV with a history of childhood abuse relative to non-abused SV | - |
| Sinclair, D. 2012 [54] | CS | CS: 21, Controls: 34 | Caucasian | Brain | ↓ | rs10052957 rs72801094 rs5871845 rs10482614 rs10482616 rs4634384 rs6190 rs1800445 rs41423247 rs6196 rs6198 | rs10052957, rs6190, rs41423247 ↓ | - | - |
| Pérez-Ortiz, J. M. 2013 [14] | CS | CS: 13 Controls: 13 | Caucasian | Brain | ↓ | - | - | - | - |
| Zhao, J. 2015 [55] | CS | CS: 17 Controls: 7 | Caucasian | Brain | No changes | - | - | - | - |
| Yin, H. 2016 [41] | CS | CS: 21, Controls: 38 | European | Brain | ↓ | rs6196 rs33388 rs33389 rs10052957 rs9324924 | No association. | - | - |
| Roy, B. 2017 [56] | SI | SI: 14, Controls: 20 | Caucasian and African-American | Blood | ↓ | - | - | - | - |
| Melhem, N. M. 2017 [57] | SA and SI | SA:38, SI:40, Controls:37 | Caucasian | Blood | SA ↓ | - | - | mRNA was significantly and negatively associated with childhood abuse. | HCC was associated with mRNA. |
| Chang, H. B. 2019 [58] | SA and SI | SA:38, SI:40, Controls:37 | Caucasian | Blood | No changes | - | - | - | No association. |
| FKBP5 | |||||||||
| Pérez-Ortiz, J. M. 2013 [14] | CS | CS: 13 Controls: 13 | Caucasian | Brain | ↓ | - | - | - | - |
| Yin, H. 2016 [41] | CS | CS: 21 Controls: 38 | European | Brain | ↓ | rs3800373 rs9296158 rs3777747 rs4713902 rs9470080 rs7757037 rs737054 rs9380529 | No association | - | - |
| Roy, B. 2017 [56] | SI | SI: 14 Controls: 20 | Caucasian and African-American | Blood | ↓ | - | - | - | - |
| Melhem, N. M. 2017 [57] | SA and SI | SA:38; SI:40 Controls:37 | Caucasian | Blood | SI ↓ | - | - | No association. | No association. |
| Chang, H. B. 2019 [58] | SA and SI | SA:38; SI:40 Controls:37 | Caucasian | Blood | mRNA was consistently correlated with heroin, painkillers, and ecstasy use. | - | - | - | No association. |
| CRHR1 | |||||||||
| Hiroi, N. 2001 [59] | CS | CS: 9 Controls: 7 | Caucasian | Brain | No change | - | - | - | - |
| Merali, Z. 2004 [60] | CS | CS: 12 Controls: 12 | Caucasian | Brain | ↓ | - | - | - | - |
| Zhao, J. 2015 [55] | CS | CS: 17 Controls: 7 | Caucasian | Brain | No change | - | - | - | - |
| Roy, B. 2017 [56] | CS | CS: 14 Controls: 20 | Caucasian and African-American | Blood | No change | - | - | - | - |
| CRHR2 | |||||||||
| Hiroi, N. 2001 [59] | CS | CS: 9 controls: 7 | Caucasian | Pituitary | No change | - | - | - | - |
| Merali, Z. 2004 [60] | CS | CS: 12 Controls: 12 | Caucasian | Brain | No change | - | - | - | - |
| Zhao, J. 2015 [55] | CS | CS: 17 Controls: 7 | Caucasian | Brain | No change | - | - | - | - |
| CRHBP | |||||||||
| Merali, Z. 2004 [60] | CS | CS: 12 Controls: 12 | Caucasian | Brain | No change | - | - | - | - |
| Zhao, J. 2015 [55] | CS | CS: 17 Controls: 7 | Caucasian | Brain | No change | - | - | - | - |
| Roy, B. 2017 [56] | SI | SI: 14 Controls: 20 | Caucasian and African-American | Blood | No change | - | - | - | - |
| CRH | |||||||||
| Merali, Z. 2004 [60] | CS | CS: 12 Controls: 12 | Caucasian | Brain | ↑ | - | - | - | - |
| Zhao, J. 2015 [55] | CS | CS: 17 Controls: 7 | Caucasian | Brain | ↑ | - | - | - | - |
| SKA2 | |||||||||
| Yin, H. 2016 [41] | CS | CS: 21 Controls: 38 | European | Brain | ↓ | rs8082544 | AG ↓ | - | - |
| rs7502947 | AG ↓ | ||||||||
| Pandey, G.N 2016 [26] | CS | CS: 52 Control: 51 | Caucasian | Brain | ↓ | - | - | - | |
| Melhem, N. M. 2017 [57] | SA and SI | SA:38, SI:40, Controls:37 | Caucasian | Blood | SI ↑ | - | - | No association. | No association. |
| Chang, H. B. 2019 [58] | SA and SI | SA:38, SI:40, Controls:37 | Caucasian | Blood | Not changes | - | - | - | No association. |
| Study | Year | Selection | Comparability | Outcome/Exposure | Score |
|---|---|---|---|---|---|
| Hiroi, N. [59] | 2001 | ★★★ | ★ ★ | ★★ | 7 |
| Merali, Z. [60] | 2004 | ★★★ | ★ ★ | ★★ | 7 |
| Papiol, S. 2007 [31] | 2007 | ★★ | ★ ★ | ★★ | 6 |
| Wasserman, D. [45] | 2008 | ★★ | ★ ★ | ★ ★ ★ | 7 |
| Willour, V. L. [32] | 2009 | ★★★ | ★ ★ | ★★ | 7 |
| Wasserman, D. [24] | 2009 | ★★ | ★ ★ | ★ ★ ★ | 7 |
| McGowan, P. [53] | 2009 | ★★ | ★ ★ | ★ ★ ★ | 7 |
| De Luca, V. [27] | 2010 | ★★★★ | ★ ★ | ★★ | 8 |
| Brent, D. [33] | 2010 | ★★★ | ★ ★ | ★★ | 7 |
| Roy, A. [34] | 2010 | ★★★★ | ★ ★ | ★★ | 8 |
| Perroud, N. [35] | 2011 | ★ ★ | ★ ★ | ★ ★ | 6 |
| Supriyanto, I. [36] | 2011 | ★ ★ ★ ★ | ★ ★ | ★ ★ | 8 |
| Ben-Efraim, Y. J. [46] | 2011 | ★ ★ ★ | ★ ★ | ★★ | 7 |
| Sinclair, D. [54] | 2012 | ★ ★ | ★ ★ | ★★ | 6 |
| Roy, A. [28] | 2012 | ★★★ | ★ ★ | ★ ★ | 7 |
| Guillaume, S. [47] | 2013 | ★ ★ | ★ ★ | ★ ★ | 6 |
| Leszczynska-Rodziewicz, A. [48] | 2013 | ★ ★ | ★ ★ | ★ ★ | 6 |
| Pérez-Ortiz, J. M. [14] | 2013 | ★ ★ ★ ★ | ★ ★ | ★ ★ | 8 |
| Leszczynska-Rodziewicz, A. [37] | 2014 | ★ ★ | ★ ★ | ★ ★ | 6 |
| Zhao, J. [55] | 2015 | ★★ | ★ ★ | ★ ★ | 6 |
| Breen, M. E. [38] | 2015 | ★★★ | ★ ★ | ★★ | 7 |
| Fudalej, S. [39] | 2015 | ★★ | ★ ★ | ★★ | 6 |
| Kaminsky, Z. [25] | 2015 | ★★★ | ★ ★ | ★★ | 7 |
| Park, S. [20] | 2016 | ★★★ | ★ ★ | ★★ | 7 |
| Breen, M. E. [40] | 2016 | ★★★★ | ★ ★ | ★★ | 8 |
| Pawlak, J. [49] | 2016 | ★★★★ | ★ ★ | ★★ | 8 |
| Pandey, G.N [26] | 2016 | ★★★★ | ★ ★ | ★★ ★ | 9 |
| Sadeh, N. [52] | 2016 | ★ ★ | ★ ★ | ★ ★ | 6 |
| Yin, H. [41] | 2016 | ★★★★ | ★ ★ | ★★ | 8 |
| Mirkovic, B. [42] | 2017 | ★★★ | ★ ★ | ★★★ ★ | 9 |
| Roy, B. [56] | 2017 | ★★★ | ★ ★ | ★★ | 7 |
| Bastos, C. R. [50] | 2017 | ★★★★ | ★ ★ | ★★★ | 9 |
| Melhem, N. M. [57] | 2017 | ★★★ | ★ ★ | ★★★ | 8 |
| Ludwig, B. [51] | 2018 | ★ ★ | ★ ★ | ★★★ | 7 |
| Chang, H. B. [58] | 2019 | ★★ | ★ ★ | ★★★ | 7 |
| Sanabrais-Jiménez, M.A. [23] | 2019 | ★★ | ★ ★ | ★★ | 6 |
| Segura, A. G. [19] | 2019 | ★★ | ★ ★ | ★★ | 6 |
| Zhang, L. [43] | 2019 | ★★★ | ★★ | ★★ | 7 |
| Nobile, B. [44] | 2020 | ★★★ | ★ ★ | ★★ | 7 |
| Berent, D. [21] | 2020 | ★★★★ | ★ ★ | ★★ ★ | 9 |
| Hernández-Díaz, Y. [22] | 2021 | ★★★ | ★ ★ ★ | ★★ ★ | 9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Díaz, Y.; Genis-Mendoza, A.D.; González-Castro, T.B.; Tovilla-Zárate, C.A.; Juárez-Rojop, I.E.; López-Narváez, M.L.; Nicolini, H. Association and Genetic Expression between Genes Involved in HPA Axis and Suicide Behavior: A Systematic Review. Genes 2021, 12, 1608. https://doi.org/10.3390/genes12101608
Hernández-Díaz Y, Genis-Mendoza AD, González-Castro TB, Tovilla-Zárate CA, Juárez-Rojop IE, López-Narváez ML, Nicolini H. Association and Genetic Expression between Genes Involved in HPA Axis and Suicide Behavior: A Systematic Review. Genes. 2021; 12(10):1608. https://doi.org/10.3390/genes12101608
Chicago/Turabian StyleHernández-Díaz, Yazmín, Alma Delia Genis-Mendoza, Thelma Beatriz González-Castro, Carlos Alfonso Tovilla-Zárate, Isela Esther Juárez-Rojop, María Lilia López-Narváez, and Humberto Nicolini. 2021. "Association and Genetic Expression between Genes Involved in HPA Axis and Suicide Behavior: A Systematic Review" Genes 12, no. 10: 1608. https://doi.org/10.3390/genes12101608
APA StyleHernández-Díaz, Y., Genis-Mendoza, A. D., González-Castro, T. B., Tovilla-Zárate, C. A., Juárez-Rojop, I. E., López-Narváez, M. L., & Nicolini, H. (2021). Association and Genetic Expression between Genes Involved in HPA Axis and Suicide Behavior: A Systematic Review. Genes, 12(10), 1608. https://doi.org/10.3390/genes12101608