
Regulation of the Fanconi Anemia DNA Repair Pathway by Phosphorylation and Monoubiquitination


Abstract
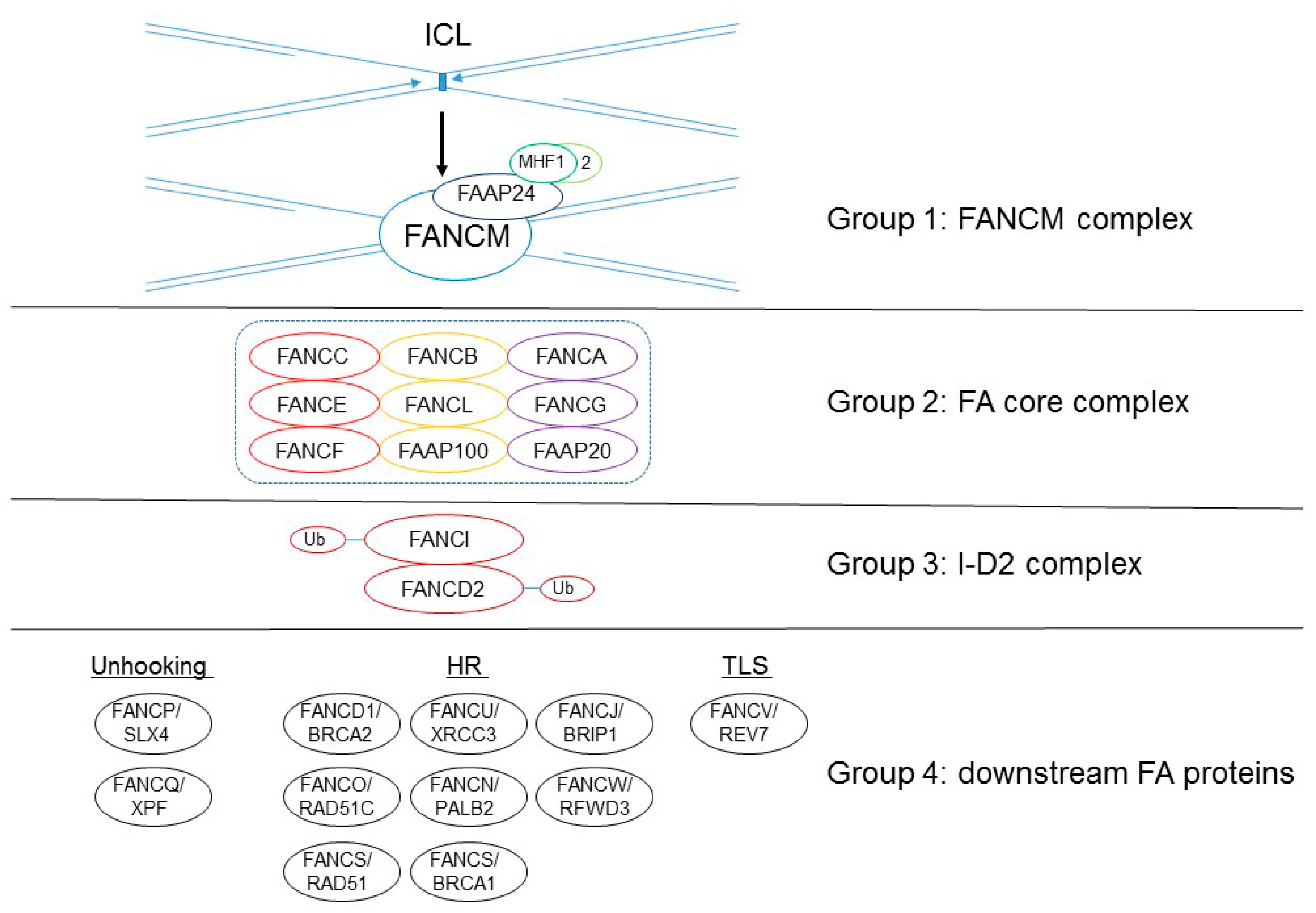
:1. Introduction

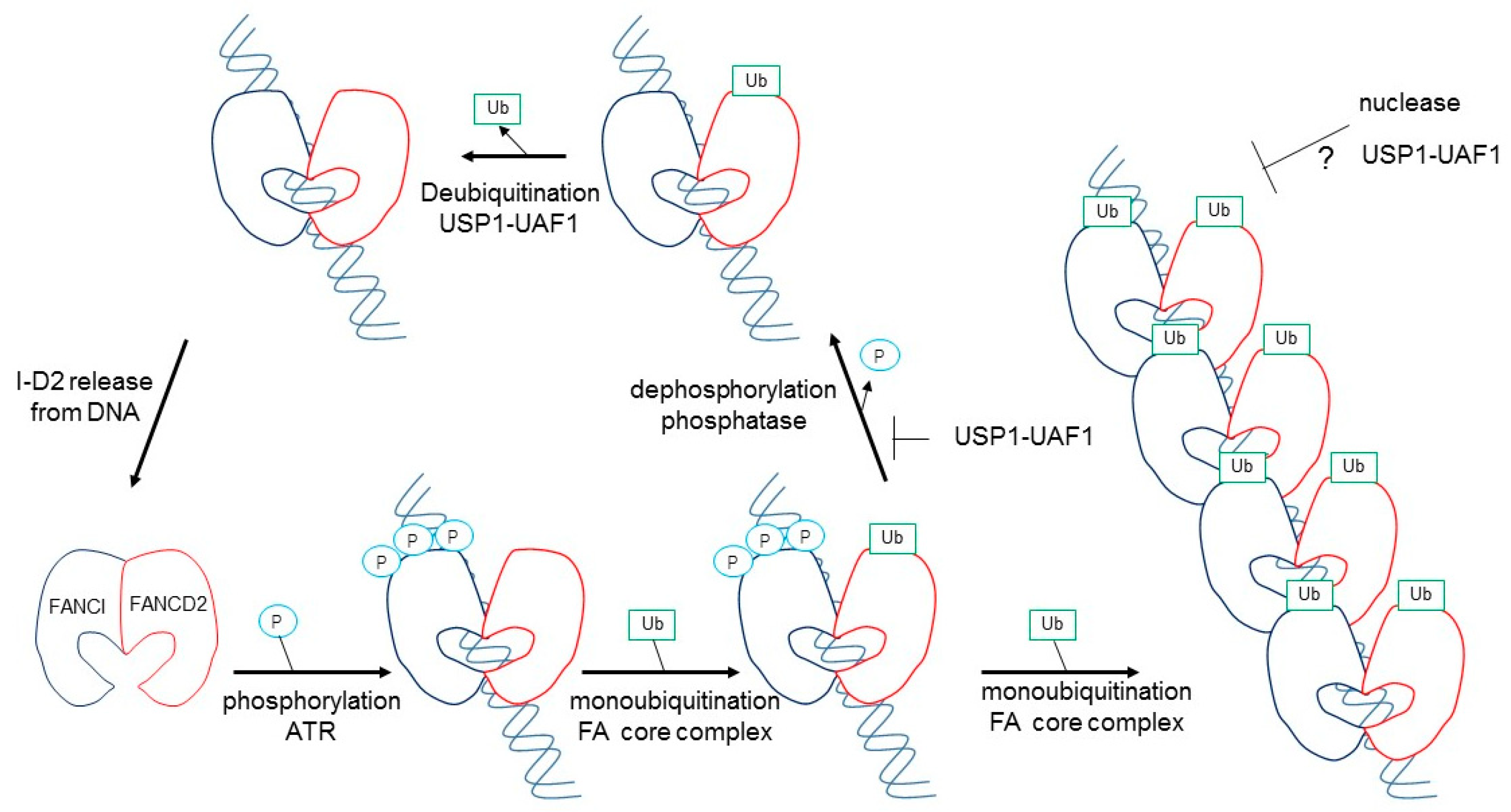
2. FANCD2 Monoubiquitination Is Key to Its Function in the FA Pathway
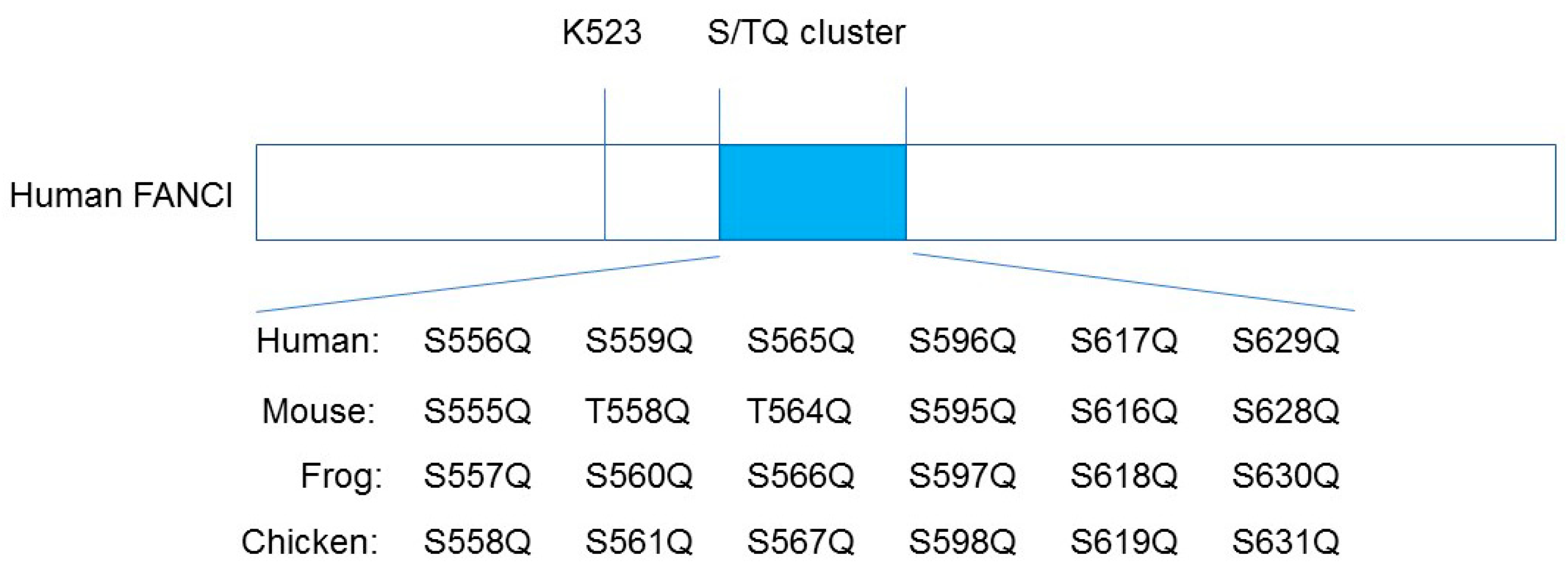
3. FANCI Phosphorylation Is a Molecular Switch Triggering FANCD2 and FANCI Monoubiquitination
4. Re-Evaluation of FANCI Phosphorylation in the Regulation of FA Pathway
5. Structure of Monoubiqutinated FANCI–FANCD2 Reveals DNA Clamping
6. Other Kinases Involved in the FA Pathway
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nalepa, G.; Clapp, D.W. Fanconi anemia and cancer: An intricate relationship. Nat. Rev. Cancer 2018, 18, 168–185. [Google Scholar] [CrossRef]
- Niraj, J.; Färkkilä, A.; D’Andrea, A. The Fanconi anemia pathway in cancer. Annu. Rev. Cancer Biol. 2019, 3, 457–478. [Google Scholar] [CrossRef]
- Renaudin, X.; Rosselli, F. The FANC/BRCA pathway release replication blockades by eliminating DNA interstrand cross-links. Genes 2020, 11, 585. [Google Scholar] [CrossRef]
- Semlow, D.R.; Walter, J.C. Mechanisms of vertebrate DNA interstrand cross-link repair. Annu. Rev. Biochem. 2021, 90, 107–135. [Google Scholar] [CrossRef]
- Sharp, M.F.; Bythell-Douglas, R.; Deans, A.J.; Crismani, W. The Fanconi anemia ubioquitin E3 ligase complex as an anti-cancer target. Mol. Cell 2021, 81, 2278–2289. [Google Scholar] [CrossRef]
- Inano, S.; Sato, K.; Katsuki, Y.; Kobayashi, W.; Tanaka, H.; Nakajima, K.; Nakada, S.; Miyoshi, H.; Knies, K.; Takaori-Kondo, A.; et al. RFWD3-mediated ubiquitination promotes timely removal of both RPA and RAD51 from DNA damage sites to facilitate homologous recombination. Mol. Cell 2017, 66, 622–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knies, K.; Inano, S.; Ramírez, M.J.; Ishiai, M.; Sarrallés, J.; Takata, M.; Schindler, D. Bialleic mutations in the ubiquitin ligase RFWD3 cause Fanconi anemia. J. Clin. Investig. 2017, 127, 3013–3027. [Google Scholar] [CrossRef] [Green Version]
- Madireddy, A.; Kosiyatrakul, S.T.; Boisvert, R.A.; Herrera-Moyano, E.; García-Rubio, M.L.; Gerhardt, J.; Vuono, E.A.; Owen, N.; Yan, Z.; Olson, S.; et al. FANCD2 facilitates replication through common fragile sites. Mol. Cell 2016, 64, 388–404. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, Y.; Iwasaki, W.M.; Kugou, K.; Takahashi, K.K.; Oda, A.; Sato, K.; Kobayashi, W.; Kawai, H.; Sakasai, R.; Takaori-Kondo, A.; et al. Replication stress induces accumulation of FANCD2 at central region of large fragile genes. Nucleic Acids Res. 2018, 46, 2932–2944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, Y.; Abe, M.; Itaya, A.; Tomida, J.; Ishiai, M.; Takaori-Kondo, A.; Taoka, M.; Isobe, T.; Takata, M. FANCD2 protects genome stability by recruiting RNA processing enzymes to resolve R-loops during mild replication stress. FEBS J. 2019, 286, 139–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, Y.; Hejna, J.; Takata, M. Regulation of R-loops and genome instability in Fanconi anemia. J. Biochem. 2019, 165, 465–470. [Google Scholar] [CrossRef]
- Li, L.; Tan, W.; Deans, A.J. Structural insight into FANCI-FANCD2 monoubiquitination. Essays Biochem. 2020, 64, 807–817. [Google Scholar]
- Shakeel, S.; Rajendra, E.; Alcón, P.; O’Reilly, F.; Chorev, D.S.; Maslen, S.; Degliesposti, G.; Russo, C.J.; He, S.; Hill, C.H.; et al. Structure of the Fanconi anemia monoubiquitin ligase complex. Nature 2019, 575, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, R.; Peralta, C.; Yaseen, A.; Pavletich, N.P. Structure of the FA core ubiquitin ligase closing the ID clamp on DNA. Nat. Struct. Mol. Biol. 2021, 28, 300–309. [Google Scholar] [CrossRef]
- Lemonidis, K.; Arkinson, C.; Rennie, M.L.; Walden, H. Mechanism, specificity, and function of FANCD2-FANCI ubiquitination and deubiquitination. FEBS J. 2021. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Ishiai, M.; Sato, K.; Tomida, J.; Kitao, H.; Kurumizaka, H.; Takata, M. Activation of the FA pathway mediated by phosphorylation and ubiquitination. Mutat. Res. 2017, 803–805, 89–95. [Google Scholar] [CrossRef]
- Matsushita, N.; Kitao, H.; Ishiai, M.; Nagashima, N.; Hirano, S.; Okawa, K.; Ohta, T.; Yu, D.S.; McHugh, P.; Hickson, I.D.; et al. A FANCD2-monoubiquitin fusion reveals hidden functions of Fanconi anemia core complex in DNA repair. Mol. Cell 2005, 19, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Ishiai, M.; Kitao, H.; Smogorzewska, A.; Tomida, J.; Kinomura, A.; Uchida, E.; Saberi, A.; Kinoshita, E.; Kinoshita-Kikuta, E.; Koike, T.; et al. FANCI phosphorylation functions as a molecular switch to turn on the Fanconi anemia pathway. Nat. Struct. Mol. Biol. 2018, 11, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Shigechi, T.; Tomida, J.; Sato, K.; Kobayashi, M.; Eykelenboom, J.K.; Pessina, F.; Zhang, Y.; Uchida, E.; Ishiai, M.; Lowndes, N.F.; et al. ATR-ATRIP kinase complex triggers activation of the Fanconi anemia DNA repair pathway. Cancer Res. 2012, 72, 1149–1156. [Google Scholar] [CrossRef] [Green Version]
- Tomida, J.; Itaya, A.; Shigechi, T.; Unno, J.; Uchida, E.; Ikura, M.; Masuda, T.; Matsuda, S.; Adachi, J.; Kobayashi, M.; et al. A novel interplay between the Fanconi anemia core complex and ATR-ATRIP kinase during DNA cross-link repair. Nucleic Acids Res. 2013, 41, 6930–6941. [Google Scholar] [CrossRef] [Green Version]
- Joo, W.; Xu, G.; Persky, N.S.; Smogorzewska, A.; Rudge, D.G.; Buzovetsky, O.; Elledge, S.J.; Pavletich, N.P. Structure of FANCI-FANCD2 complex: Insights into the Fanconi anemia DNA repair pathway. Science 2011, 333, 312–316. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Toda, M.; Ishiai, M.; Takata, M.; Kurumizaka, H. DNA robustly stimulates FANCD2 monoubiquitination in the complex with FANCI. Nucleic Acids Res. 2012, 40, 4553–4561. [Google Scholar] [CrossRef] [Green Version]
- Longerich, S.; Kwon, Y.; Tsai, M.-S.; Hing, A.S.; Kupfer, G.M.; Sung, P. Regulation of FANCD2 and FANCI monoubiquitination by their interaction and by DNA. Nucleic Acids Res. 2014, 42, 5657–5670. [Google Scholar] [CrossRef] [Green Version]
- Rajendra, E.; Oestergaard, V.H.; Langevin, F.; Wang, W.; Dornan, G.L.; Patel, K.J.; Passmore, L.A. The genetic and biochemical basis of FANCD2 monoubiquitination. Mol. Cell 2014, 54, 858–869. [Google Scholar] [CrossRef]
- van Twest, S.; Murphy, V.J.; Hodson, C.; Tan, W.; Swuec, P.; O’Rourke, J.J.; Heierhorst, J.; Crismani, W.; Deans, A.J. Mechanism of ubiquitination and deubiquitination in the Fanconi anemia pathway. Mol. Cell. 2017, 65, 247–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swuec, P.; Renault, L.; Borg, A.; Shah, F.; Murphy, V.J.; van Twest, M.; Snijders, B.; Deans, A.J.; Costa, A. The FA core complex contains a homodimeric catalytic module for the symmetric mono-ubiquitination of FANCI-FANCD2. Cell Rep. 2017, 18, 611–623. [Google Scholar] [CrossRef] [Green Version]
- Cheung, R.S.; Castella, A.A.; Gafken, P.R.; Tucker, N.; Taniguchi, T. Ubiquitination-linked phosphorylation of FANCI S/TQ cluster contributes to activation of the Fanconi anemia I/D2 complex. Cell Rep. 2017, 19, 2432–2440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, W.; van Twest, S.; Murphy, V.J.; Deans, A.J. ATR-mediated FANCI phosphorylation regulates both ubiquitination and deubiquitination of FANCD2. Front. Cell. Dev. Biol. 2020, 8, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarren, A.; Chadhury, I.; Adams, N.; Sobek, A. Fanconi anemia proteins FANCD2 and FANCI exhibited different DNA damage responses during S-phase. Nucleic Acids Res. 2012, 40, 8425–8439. [Google Scholar] [CrossRef]
- Tan, W.; van Twest, S.; Leis, A.; Bythell-Douglas, R.; Murphy, V.J.; Sharp, M.; Parker, M.W.; Crismani, W.; Deans, A. Monoubiquiination by human Fanconi anemia core complex clamps FANCI:FANCD2 on DNA in filamentous arrays. eLife 2020, 9, e54128. [Google Scholar] [CrossRef]
- Wang, R.; Wang, S.; Dhar, A.; Peralta, C.; Pavletich, N. DNA clamp function of the monoubiquitinated Fanconi anemia ID complex. Nature 2020, 580, 278–282. [Google Scholar] [CrossRef]
- Alcón, P.; Shakeel, S.; Chen, Z.A.; Rappsilber, J.; Patel, K.J.; Passmore, L.A. FANCD2-FANCI is a clamp stabilized on DNA by monoubiquitination of FANCD2 during DNA repair. Nat. Struct. Mol. Biol. 2020, 27, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Rennie, M.; Lemonidis, K.; Arkinson, C.; Chaugule, V.K.; Clarke, M.; Steetley, J.; Spagnolo, L.; Walden, H. Differential functions of FANCI and FANCD2 ubiquitination stabilize ID2complex on DNA. EMBO Rep. 2020, 21, e50133. [Google Scholar] [CrossRef] [PubMed]
- Andreassen, P.R.; D’Andrea, A.D.; Taniguchi, T. ATR couples FANCD2 monoubiquitination to the DNA-damage response. Genes Dev. 2004, 18, 1958–1963. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Martinez, D.; Kupculak, M.; Yang, D.; Yoshikawa, Y.; Liang, C.-C.; Wu, R.; Gygi, S.P.; Cohn, M.A. Phosphorylation of FANCD2 inhibits the FANCD2/FANCI complex and suppresses the Fanconi Anemia pathway in the absence of DNA damage. Cell Rep. 2019, 27, 2990–3005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meetei, A.R.; Medhurst, A.L.; Ling, C.; Xue, Y.; Singh, T.R.; Bier, P.; Steltenpool, J.; Stone, S.; Dokal, I.; Mathew, C.G. A human ortholog of archaeal DNA repair protein Hef is defective in Fancini anemia complementation group M. Nat. Genet. 2005, 37, 958–963. [Google Scholar] [CrossRef] [Green Version]
- Singh, T.R.; Ali, A.M.; Paramasivam, M.; Pradhan, A.; Wahengbam, K.; Seidman, M.M.; Meetei, A.R. ATR-dependent phosphoryklation of FANCM at serine 1045 is essential for FANCM functions. Cancer Res. 2013, 73, 4300–4310. [Google Scholar] [CrossRef] [Green Version]
- Deans, A.J.; Khanna, K.K.; McNees, C.J.; Mercurio, C.; Heierhorst, J.; McArthur, G.A. Cyclin-dependent kinase 2 functions in normal DNA repair and is a therapeutic target in BRCA1-deficient cancers. Cancer Res. 2006, 66, 8219–8226. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Kennedy, R.D.; Ray, K.; Stuckert, P.; Ellenberger, T.; D’Andrea, A.D. Chk1 mediated phosphorylation of FANCE is required for the Fanconi anemia/BRCA pathway. Mol. Cell Biol. 2007, 27, 3098–3108. [Google Scholar] [CrossRef] [Green Version]
- Collins, N.B.; Wilson, J.B.; Bush, T.; Thomashevski, A.; Roberts, K.J.; Jones, N.J.; Kupfer, G.M. ATR-dependent phosphorylation of FANCA on serine 1449 after DNA damage is important for FA pathway function. Blood 2009, 113, 2181–2190. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, T.; Kupfer, G.M.; Naf, D.; Suliman, A.; Joenje, H.; Asano, S.; D’Andrea, A.D. The Fanconi anemia pathway requires FAA phosphorylation and FAA/FAC nuclear accumulation. Proc. Natl. Acad. Sci. USA 1998, 95, 13085–13090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, J.; Qiao, F.; Wilson, J.B.; Zhi, G.; Bucheimer, N.R.; Jones, N.J.; Kupfer, G.M. Phosphorylation of Fanconi anemia (FA) complementation group G protein, FANCG, at serine 7 is important for function of the FA pathway. J. Biol. Chem. 2004, 279, 46035–46045. [Google Scholar]
- Hu, W.-F.; Krieger, K.L.; Lagundžin, D.; Cheung, R.S.; Taniguchi, T.; Jonson, K.R.; Bessho, T.; Monteiro, N.A.; Woods, N.T. CTDP1 regulates breast cancer survival and DNA repair through BRCT-specific interactions with FANCI. Cell Death Discov. 2019, 5, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishiai, M. Regulation of the Fanconi Anemia DNA Repair Pathway by Phosphorylation and Monoubiquitination. Genes 2021, 12, 1763. https://doi.org/10.3390/genes12111763
Ishiai M. Regulation of the Fanconi Anemia DNA Repair Pathway by Phosphorylation and Monoubiquitination. Genes. 2021; 12(11):1763. https://doi.org/10.3390/genes12111763
Chicago/Turabian StyleIshiai, Masamichi. 2021. "Regulation of the Fanconi Anemia DNA Repair Pathway by Phosphorylation and Monoubiquitination" Genes 12, no. 11: 1763. https://doi.org/10.3390/genes12111763
APA StyleIshiai, M. (2021). Regulation of the Fanconi Anemia DNA Repair Pathway by Phosphorylation and Monoubiquitination. Genes, 12(11), 1763. https://doi.org/10.3390/genes12111763