
The FLA4-FEI Pathway: A Unique and Mysterious Signaling Module Related to Cell Wall Structure and Stress Signaling

Abstract
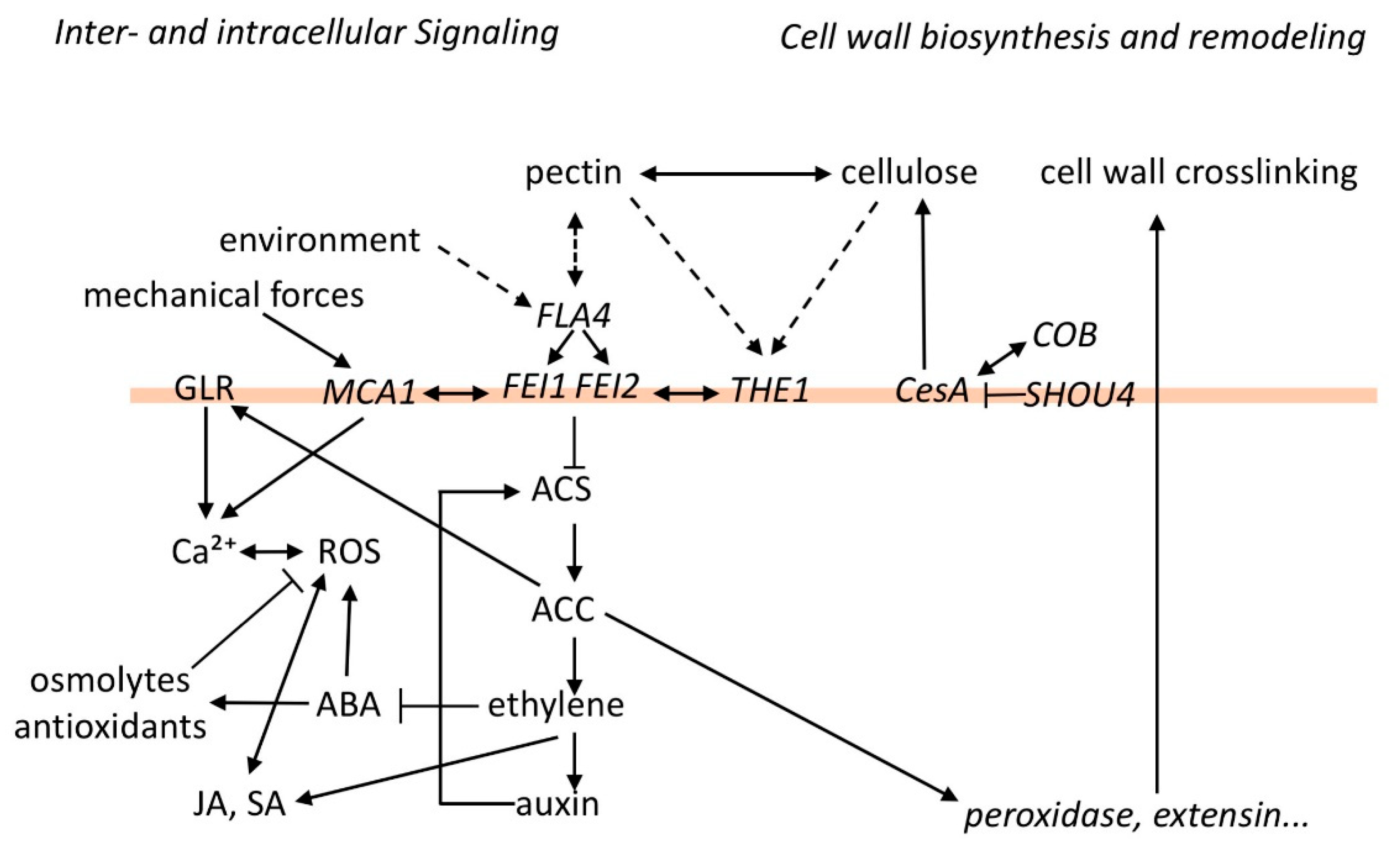
1. Introduction
2. FLA4 and the Partially Redundant FEI1 and FEI2 Loci Act in a Linear Genetic Pathway Affecting Both Cellulose and Pectin
3. The FLA4-FEI Pathway Genetically Interacts with Primary Cell Wall Biosynthesis and Damage at Various Levels
4. Interactions with Growth Regulator Signaling
4.1. Control of Ethylene-Independent ACC Signaling
4.2. Interactions with Abscisic Acid (ABA) and Auxin Signaling
5. The FLA4-FEI Pathway Seen from the Transcriptomic Perspective
6. The Relevance of FLA4-FEI Pathway for Agronomic Traits
7. Too Many FEI Interactors! Which Ones Are Relevant?
8. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Somerville, C.; Coburn, P.S.; Pillar, C.M.; Jett, B.D.; Haas, W.; Gilmore, M.S. Toward a Systems Approach to Understanding Plant Cell Walls. Science 2004, 306, 2206–2211. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.E. Cell Wall Integrity Signaling in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2005, 69, 262–291. [Google Scholar] [CrossRef] [PubMed]
- Kechagia, J.Z.; Ivaska, J.; Roca-Cusachs, P. Integrins as biomechanical sensors of the microenvironment. Nat. Rev. Mol. Cell Biol. 2019, 20, 457–473. [Google Scholar] [CrossRef] [PubMed]
- Nazemi, M.; Rainero, E. Cross-Talk between the Tumor Microenvironment, Extracellular Matrix, and Cell Metabolism in Cancer. Front. Oncol. 2020, 10, 239. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. Integrins. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef]
- Wolf, S.; Hématy, K.; Höfte, H. Growth Control and Cell Wall Signaling in Plants. Annu. Rev. Plant Biol. 2012, 63, 381–407. [Google Scholar] [CrossRef]
- Wolf, S. Plant cell wall signalling and receptor-like kinases. Biochem. J. 2017, 474, 471–492. [Google Scholar] [CrossRef]
- Vaahtera, L.; Schulz, J.; Hamann, T. Cell wall integrity maintenance during plant development and interaction with the environment. Nat. Plants 2019, 5, 924–932. [Google Scholar] [CrossRef]
- Seifert, G.J.; Blaukopf, C. Irritable Walls: The Plant Extracellular Matrix and Signaling. Plant Physiol. 2010, 153, 467–478. [Google Scholar] [CrossRef]
- Boisson-Dernier, A.; Kessler, S.A.; Grossniklaus, U. The walls have ears: The role of plant CrRLK1Ls in sensing and transducing extracellular signals. J. Exp. Bot. 2011, 62, 1581–1591. [Google Scholar] [CrossRef]
- Novaković, L.; Guo, T.; Bacic, A.; Sampathkumar, A.; Johnson, K.L. Hitting the Wall—Sensing and Signaling Pathways Involved in Plant Cell Wall Remodeling in Response to Abiotic Stress. Plants 2018, 7, 89. [Google Scholar] [CrossRef] [PubMed]
- Kohorn, B.D.; Kohorn, S.L. The cell wall-associated kinases, WAKs, as pectin receptors. Front. Plant Sci. 2012, 3, 88. [Google Scholar] [CrossRef] [PubMed]
- Lindner, H.; Müller, L.M.; Boisson-Dernier, A.; Grossniklaus, U. CrRLK1L receptor-like kinases: Not just another brick in the wall. Curr. Opin. Plant Biol. 2012, 15, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Van Der Does, D.; Ladwig, F.; Sticht, C.; Kolbeck, A.; Schürholz, A.-K.; Augustin, S.; Keinath, N.; Rausch, T.; Greiner, S.; et al. A receptor-like protein mediates the response to pectin modification by activating brassinosteroid signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 15261–15266. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Mravec, J.; Greiner, S.; Mouille, G.; Höfte, H. Plant Cell Wall Homeostasis Is Mediated by Brassinosteroid Feedback Signaling. Curr. Biol. 2012, 22, 1732–1737. [Google Scholar] [CrossRef]
- Van Der Does, D.; Boutrot, F.; Engelsdorf, T.; Rhodes, J.; McKenna, J.F.; Vernhettes, S.; Koevoets, I.T.; Tintor, N.; Veerabagu, M.; Miedes, E.; et al. The Arabidopsis leucine-rich repeat receptor kinase MIK2/LRR-KISS connects cell wall integrity sensing, root growth and response to abiotic and biotic stresses. PLoS Genet. 2017, 13, e1006832. [Google Scholar] [CrossRef]
- Chaudhary, A.; Chen, X.; Gao, J.; Lesniewska, B.; Hammerl, R.; Dawid, C.; Schneitz, K. The Arabidopsis receptor kinase STRUBBELIG regulates the response to cellulose deficiency. PLoS Genet. 2020, 16, e1008433. [Google Scholar] [CrossRef]
- Zhu, J. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Johnson, K.L.; Jones, B.J.; Bacic, A.; Schultz, C.J. The Fasciclin-Like Arabinogalactan Proteins of Arabidopsis. A Multigene Family of Putative Cell Adhesion Molecules. Plant Physiol. 2003, 133, 1911–1925. [Google Scholar] [CrossRef]
- Shi, H.; Kim, Y.; Guo, Y.; Stevenson, B.; Zhu, J. The Arabidopsis SOS5 Locus Encodes a Putative Cell Surface Adhesion Protein and Is Required for Normal Cell Expansion. Plant Cell 2003, 15, 19–32. [Google Scholar] [CrossRef]
- Hauser, M.-T.; Morikami, A.; Benfey, P.N. Conditional root expansion mutants of Arabidopsis. Development 1995, 121, 1237–1252. [Google Scholar] [PubMed]
- Baskin, T.; Betzner, A.; Hoggart, R.; Cork, A.; Williamson, R. Root Morphology Mutants in Arabidopsis thaliana. Funct. Plant Biol. 1992, 19, 427–437. [Google Scholar] [CrossRef]
- Mosher, D.F.; Johansson, M.W.; Gillis, M.E.; Annis, U.S. Periostin and TGF-β-induced protein: Two peas in a pod? Crit. Rev. Biochem. Mol. Biol. 2015, 50, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Squadrito, M.L.; De Palma, M. A niche role for periostin and macrophages in glioblastoma. Nat. Cell Biol. 2015, 17, 107–109. [Google Scholar] [CrossRef] [PubMed]
- Seifert, G.J. Fascinating Fasciclins: A Surprisingly Widespread Family of Proteins that Mediate Interactions between the Cell Exterior and the Cell Surface. Int. J. Mol. Sci. 2018, 19, 1628. [Google Scholar] [CrossRef]
- Chae, H.S.; Kieber, J.J. Eto Brute? Role of ACS turnover in regulating ethylene biosynthesis. Trends Plant Sci. 2005, 10, 291–296. [Google Scholar] [CrossRef]
- Xu, S.-L.; Rahman, A.; Baskin, T.I.; Kieber, J.J. Two Leucine-Rich Repeat Receptor Kinases Mediate Signaling, Linking Cell Wall Biosynthesis and ACC Synthase in Arabidopsis. Plant Cell 2008, 20, 3065–3079. [Google Scholar] [CrossRef]
- Schindelman, G.; Morikami, A.; Jung, J.; Baskin, T.I.; Carpita, N.C.; Derbyshire, P.; McCann, M.C.; Benfey, P.N. COBRA encodes a putative GPI-anchored protein, which is polarly localized and necessary for oriented cell expansion in Arabidopsis. Genes Dev. 2001, 15, 1115–1127. [Google Scholar] [CrossRef]
- Arioli, T.; Peng, L.; Betzner, A.S.; Burn, J.; Wittke, W.; Herth, W.; Camilleri, C.; Höfte, H.; Plazinski, J.; Birch, R.; et al. Molecular Analysis of Cellulose Biosynthesis inArabidopsis. Science 1998, 279, 717–720. [Google Scholar] [CrossRef]
- Fagard, M.; Desnos, T.; Desprez, T.; Goubet, F.; Réfrégier, G.; Mouille, G.; McCann, M.; Rayon, C.; Vernhettes, S.; Höfte, H. PROCUSTE1 Encodes a Cellulose Synthase Required for Normal Cell Elongation Specifically in Roots and Dark-Grown Hypocotyls of Arabidopsis. Plant Cell 2000, 12, 2409–2424. [Google Scholar] [CrossRef][Green Version]
- Voiniciuc, C.; Yang, B.; Schmidt, M.H.-W.; Günl, M.; Usadel, B. Starting to Gel: How Arabidopsis Seed Coat Epidermal Cells Produce Specialized Secondary Cell Walls. Int. J. Mol. Sci. 2015, 16, 3452–3473. [Google Scholar] [CrossRef] [PubMed]
- North, H.M.; Berger, A.; Saez-Aguayo, S.; Ralet, M.-C. Understanding polysaccharide production and properties using seed coat mutants: Future perspectives for the exploitation of natural variants. Ann. Bot. 2014, 114, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Harpaz-Saad, S.; McFarlane, H.E.; Xu, S.-L.; Divi, U.K.; Forward, B.; Western, T.L.; Kieber, J.J. Cellulose synthesis via the FEI2 RLK/SOS5 pathway and CELLULOSE SYNTHASE 5 is required for the structure of seed coat mucilage in Arabidopsis. Plant J. 2011, 68, 941–953. [Google Scholar] [CrossRef] [PubMed]
- McCann, M.C.; Bush, M.; Milioni, D.; Sado, P.; Stacey, N.J.; Catchpole, G.; Defernez, M.; Carpita, N.C.; Hofte, H.; Ulvskov, P.; et al. Approaches to understanding the functional architecture of the plant cell wall. Phytochemistry 2001, 57, 811–821. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Re-constructing our models of cellulose and primary cell wall assembly. Curr. Opin. Plant Biol. 2014, 22, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Alberts, B.; Johnson, A.; Lewis, J.; Morgan, D.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell, 6th ed.; Garland Science: New York, NY, USA, 2015. [Google Scholar]
- Buchanan, B.B.; Gruissem, W.; Jones, R.L. Biochemistry & Molecular Biology of Plants; American Society of Plant Physiologists: Rockville, MD, USA, 2000. [Google Scholar]
- Park, Y.B.; Cosgrove, D.J. A Revised Architecture of Primary Cell Walls Based on Biomechanical Changes Induced by Substrate-Specific Endoglucanases. Plant Physiol. 2012, 158, 1933–1943. [Google Scholar] [CrossRef]
- Du, J.; Kirui, A.; Huang, S.; Wang, L.; Barnes, W.J.; Kiemle, S.N.; Zheng, Y.; Rui, Y.; Ruan, M.; Qi, S.; et al. Mutations in the Pectin Methyltransferase QUASIMODO2 Influence Cellulose Biosynthesis and Wall Integrity in Arabidopsis. Plant Cell 2020, 32, 3576–3597. [Google Scholar] [CrossRef] [PubMed]
- Bradley, D.J.; Kjellbom, P.; Lamb, C.J. Elicitor- and wound-induced oxidative cross-linking of a proline-rich plant cell wall protein: A novel, rapid defense response. Cell 1992, 70, 21–30. [Google Scholar] [CrossRef]
- Kjellbom, P.; Snogerup, L.; Stohr, C.; Reuzeau, C.; McCabe, P.F.; Pennell, R.I. Oxidative cross-linking of plasma membrane arabinogalactan proteins. Plant J. 1997, 12, 1189–1196. [Google Scholar] [CrossRef]
- Raggi, S.; Ferrarini, A.; Delledonne, M.; Dunand, C.; Ranocha, P.; De Lorenzo, G.; Cervone, F.; Ferrari, S. The Arabidopsis thaliana Class III Peroxidase AtPRX71 Negatively Regulates Growth under Physiological Conditions and in Response to Cell Wall Damage. Plant Physiol. 2015, 169, 2513–2525. [Google Scholar] [CrossRef]
- Polko, J.K.; Barnes, W.J.; Voiniciuc, C.; Doctor, S.; Steinwand, B.; Hill, J.L.; Tien, M.; Pauly, M.; Anderson, C.T.; Kieber, J.J. SHOU4 Proteins Regulate Trafficking of Cellulose Synthase Complexes to the Plasma Membrane. Curr. Biol. 2018, 28, 3174–3182.e6. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, J.S.; Tsai, A.Y.-L.; Xue, H.; Voiniciuc, C.; Šola, K.; Seifert, G.J.; Mansfield, S.D.; Haughn, G.W. SALT-OVERLY SENSITIVE5 Mediates Arabidopsis Seed Coat Mucilage Adherence and Organization through Pectins. Plant Physiol. 2014, 165, 991–1004. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, J.S.; Crepeau, M.-J.; Ralet, M.-C.; Seifert, G.J.; North, H.M. Dissecting Seed Mucilage Adherence Mediated by FEI2 and SOS5. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Ben-Tov, D.; Idan-Molakandov, A.; Hugger, A.; Ben-Shlush, I.; Günl, M.; Yang, B.; Usadel, B.; Harpaz-Saad, S. The role of COBRA - LIKE 2 function, as part of the complex network of interacting pathways regulating Arabidopsis seed mucilage polysaccharide matrix organization. Plant J. 2018, 94, 497–512. [Google Scholar] [CrossRef] [PubMed]
- Basu, D.; Tian, L.; Debrosse, T.; Poirier, E.; Emch, K.; Herock, H.; Travers, A.; Showalter, A.M. Glycosylation of a Fasciclin-Like Arabinogalactan-Protein (SOS5) Mediates Root Growth and Seed Mucilage Adherence via a Cell Wall Receptor-Like Kinase (FEI1/FEI2) Pathway in Arabidopsis. PLoS ONE 2016, 11, e0145092. [Google Scholar] [CrossRef]
- Xue, H.; Veit, C.; Abas, L.; Tryfona, T.; Maresch, D.; Ricardi, M.M.; Estevez, J.M.; Strasser, R.; Seifert, G.J. Arabidopsis thaliana FLA4 functions as a glycan-stabilized soluble factor via its carboxy-proximal Fasciclin 1 domain. Plant J. 2017, 91, 613–630. [Google Scholar] [CrossRef]
- Engelsdorf, T.; Gigli-Bisceglia, N.; Veerabagu, M.; McKenna, J.F.; Vaahtera, L.; Augstein, F.; Van Der Does, D.; Zipfel, C.; Hamann, T. The plant cell wall integrity maintenance and immune signaling systems cooperate to control stress responses in Arabidopsis thaliana. Sci. Signal. 2018, 11, eaao3070. [Google Scholar] [CrossRef]
- Feng, W.; Kita, D.; Peaucelle, A.; Cartwright, H.N.; Doan, V.; Duan, Q.; Liu, M.-C.; Maman, J.; Steinhorst, L.; Schmitz-Thom, I.; et al. The FERONIA Receptor Kinase Maintains Cell-Wall Integrity during Salt Stress through Ca2+ Signaling. Curr. Biol. 2018, 28, 666–675.e5. [Google Scholar] [CrossRef]
- Denness, L.; McKenna, J.F.; Segonzac, C.; Wormit, A.; Madhou, P.; Bennett, M.; Mansfield, J.; Zipfel, C.; Hamann, T. Cell Wall Damage-Induced Lignin Biosynthesis Is Regulated by a Reactive Oxygen Species- and Jasmonic Acid-Dependent Process in Arabidopsis. Plant Physiol. 2011, 156, 1364–1374. [Google Scholar] [CrossRef]
- Xue, H.; Seifert, G.J. FASCICLIN LIKE ARABINOGALACTAN PROTEIN 4 and RESPIRATORY BURST OXIDASE HOMOLOG D and F independently modulate abscisic acid signaling. Plant Signal. Behav. 2015, 10. [Google Scholar] [CrossRef]
- Polko, J.K.; Kieber, J.J. 1-Aminocyclopropane 1-Carboxylic Acid and Its Emerging Role as an Ethylene-Independent Growth Regulator. Front. Plant Sci. 2019, 10, 1602. [Google Scholar] [CrossRef] [PubMed]
- Tsang, D.L.; Edmond, C.; Harrington, J.L.; Nühse, T.S. Cell Wall Integrity Controls Root Elongation via a General 1-Aminocyclopropane-1-Carboxylic Acid-Dependent, Ethylene-Independent Pathway. Plant Physiol. 2011, 156, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Vanderstraeten, L.; Depaepe, T.; Bertrand, S.; Van Der Straeten, D. The Ethylene Precursor ACC Affects Early Vegetative Development Independently of Ethylene Signaling. Front. Plant Sci. 2019, 10, 1591. [Google Scholar] [CrossRef] [PubMed]
- Mou, W.; Kao, Y.-T.; Michard, E.; Simon, A.A.; Li, D.; Wudick, M.M.; Lizzio, M.A.; Feijó, J.A.; Chang, C. Ethylene-independent signaling by the ethylene precursor ACC in Arabidopsis ovular pollen tube attraction. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Argueso, C.T.; Hansen, M.; Kieber, J.J. Regulation of Ethylene Biosynthesis. J. Plant Growth Regul. 2007, 26, 92–105. [Google Scholar] [CrossRef]
- Turupcu, A.; Almohamed, W.; Oostenbrink, C.; Seifert, G.J. A speculation on the tandem fasciclin 1 repeat of FLA4 proteins in angiosperms. Plant Signal. Behav. 2018, 13, e1507403-5. [Google Scholar] [CrossRef]
- Guan, Q.; Wu, J.; Yue, X.; Zhang, Y.; Zhu, J. A Nuclear Calcium-Sensing Pathway Is Critical for Gene Regulation and Salt Stress Tolerance in Arabidopsis. PLoS Genet. 2013, 9, e1003755. [Google Scholar] [CrossRef]
- Steinhorst, L.; Kudla, J. Signaling in cells and organisms—Calcium holds the line. Curr. Opin. Plant Biol. 2014, 22, 14–21. [Google Scholar] [CrossRef]
- Kwon, Y.; Shen, J.; Lee, M.H.; Geem, K.R.; Jiang, L.; Hwang, I. AtCAP2 is crucial for lytic vacuole biogenesis during germination by positively regulating vacuolar protein trafficking. Proc. Natl. Acad. Sci. USA 2018, 115, E1675–E1683. [Google Scholar] [CrossRef]
- Markakis, M.N.; De Cnodder, T.; Lewandowski, M.; Simon, D.; Boron, A.; Balcerowicz, D.; Doubbo, T.; Soubigou-Taconnat, L.; Renou, J.-P.; Höfte, H.; et al. Identification of genes involved in the ACC-mediated control of root cell elongation in Arabidopsis thaliana. BMC Plant Biol. 2012, 12, 208. [Google Scholar] [CrossRef]
- Steinwand, B.J.; Xu, S.-L.; Polko, J.K.; Doctor, S.M.; Westafer, M.; Kieber, J.J. Alterations in Auxin Homeostasis Suppress Defects in Cell Wall Function. PLoS ONE 2014, 9, e98193. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Schumaker, K.; Zhu, J. Molecular genetic perspectives on cross-talk and specificity in abiotic stress signalling in plants. J. Exp. Bot. 2004, 55, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Seifert, G.J.; Xue, H.; Acet, T. The Arabidopsis thaliana FASCICLIN LIKE ARABINOGALACTAN PROTEIN 4 gene acts synergistically with abscisic acid signalling to control root growth. Ann. Bot. 2014, 114, 1125–1133. [Google Scholar] [CrossRef] [PubMed]
- Acet, T.; Kadıoğlu, A. SOS5 gene-abscisic acid crosstalk and their interaction with antioxidant system in Arabidopsis thaliana under salt stress. Physiol. Mol. Biol. Plants 2020, 26, 1831–1845. [Google Scholar] [CrossRef] [PubMed]
- Seki, M.; Umezawa, T.; Urano, K.; Shinozaki, K. Regulatory metabolic networks in drought stress responses. Curr. Opin. Plant Biol. 2007, 10, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Postiglione, A.E.; Muday, G.K. The Role of ROS Homeostasis in ABA-Induced Guard Cell Signaling. Front. Plant Sci. 2020, 11, 968. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Chan, Z. ROS Regulation During Abiotic Stress Responses in Crop Plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Tuan, P.A.; Izydorczyk, M.S.; Ayele, B.T. Ethylene regulates post-germination seedling growth in wheat through spatial and temporal modulation of ABA/GA balance. J. Exp. Bot. 2020, 71, 1985–2004. [Google Scholar] [CrossRef]
- Tao, J.-J.; Chen, H.-W.; Ma, B.; Zhang, W.-K.; Chen, S.-Y.; Zhang, J.-S. The Role of Ethylene in Plants Under Salinity Stress. Front. Plant Sci. 2015, 6, 1059. [Google Scholar] [CrossRef]
- Persson, S.; Wei, H.; Milne, J.; Page, G.P.; Somerville, C.R. Identification of genes required for cellulose synthesis by regression analysis of public microarray data sets. Proc. Natl. Acad. Sci. USA 2005, 102, 8633–8638. [Google Scholar] [CrossRef]
- Brown, D.M.; Zeef, L.A.; Ellis, J.; Goodacre, R.; Turner, S.R. Identification of Novel Genes in Arabidopsis Involved in Secondary Cell Wall Formation Using Expression Profiling and Reverse Genetics. Plant Cell 2005, 17, 2281–2295. [Google Scholar] [CrossRef] [PubMed]
- Usadel, B.; Obayashi, T.; Mutwil, M.; Giorgi, F.M.; Bassel, G.W.; Tanimoto, M.; Chow, A.; Steinhauser, D.; Persson, S.; Provart, N.J. Co-expression tools for plant biology: Opportunities for hypothesis generation and caveats. Plant Cell Environ. 2009, 32, 1633–1651. [Google Scholar] [CrossRef] [PubMed]
- Ruprecht, C.; Persson, S. Co-expression of cell-wall related genes: New tools and insights. Front. Plant Sci. 2012, 3, 83. [Google Scholar] [CrossRef]
- Klepikova, A.V.; Kasianov, A.S.; Gerasimov, E.S.; Logacheva, M.D.; Penin, A.A. A high resolution map of the Arabidopsis thaliana developmental transcriptome based on RNA-seq profiling. Plant J. 2016, 88, 1058–1070. [Google Scholar] [CrossRef]
- Mutwil, M.; Øbro, J.; Willats, W.G.T.; Persson, S. GeneCAT—Novel webtools that combine BLAST and co-expression analyses. Nucleic Acids Res. 2008, 36, W320–W326. [Google Scholar] [CrossRef]
- Xi, L.; Na Wu, X.; Gilbert, M.; Schulze, W.X. Classification and Interactions of LRR Receptors and Co-receptors Within the Arabidopsis Plasma Membrane—An Overview. Front. Plant Sci. 2019, 10, 472. [Google Scholar] [CrossRef]
- Smakowska-Luzan, E.; Mott, G.A.; Parys, K.; Stegmann, M.; Howton, T.C.; Layeghifard, M.; Neuhold, J.; Lehner, A.; Kong, J.; Grünwald, K.; et al. An extracellular network of Arabidopsis leucine-rich repeat receptor kinases. Nat. Cell Biol. 2018, 553, 342–346. [Google Scholar] [CrossRef] [PubMed]
- Myśków, B.; Góralska, M.; Lenarczyk, N.; Czyczyło-Mysza, I.; Stojałowski, S. Putative candidate genes responsible for leaf rolling in rye (Secale cereale L.). BMC Genet. 2018, 19, 57. [Google Scholar] [CrossRef]
- Weiberg, A.; Wang, M.; Lin, F.-M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.-D.; Jin, H. Fungal Small RNAs Suppress Plant Immunity by Hijacking Host RNA Interference Pathways. Science 2013, 342, 118–123. [Google Scholar] [CrossRef]
- Wang, M.; Weiberg, A.; Dellota, E., Jr.; Yamane, D.; Jin, H. Botrytis small RNA Bc-siR37 suppresses plant defense genes by cross-kingdom RNAi. RNA Biol. 2017, 14, 421–428. [Google Scholar] [CrossRef]
- Qu, S.; Shan, X.; Lin, J.; Song, Y.; Zhang, X. THESEUS1 positively modulates plant defense responses against Botrytis cinerea through GUANINE EXCHANGE FACTOR4 signaling. J. Integr. Plant Biol. 2017, 59, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Xuan, Y.; Xu, M.; Wang, R.-S.; Ho, C.-H.; LaLonde, S.; You, C.H.; Sardi, M.I.; Parsa, S.A.; Smith-Valle, E.; et al. Border Control--A Membrane-Linked Interactome of Arabidopsis. Science 2014, 344, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Hooper, C.M.; Castleden, I.R.; Tanz, S.K.; Aryamanesh, N.; Millar, A.H. SUBA4: The interactive data analysis centre for Arabidopsis subcellular protein locations. Nucleic Acids Res. 2017, 45, D1064–D1074. [Google Scholar] [CrossRef] [PubMed]
- Bryant, N.; Lloyd, J.; Sweeney, C.; Myouga, F.; Meinke, D. Identification of Nuclear Genes Encoding Chloroplast-Localized Proteins Required for Embryo Development in Arabidopsis. Plant Physiol. 2011, 155, 1678–1689. [Google Scholar] [CrossRef] [PubMed]
- Moody, R.G.; Williamson, M.P. Structure and function of a bacterial Fasciclin I Domain Protein elucidates function of related cell adhesion proteins such as TGFBIp and periostin. FEBS Open Bio 2013, 3, 71–77. [Google Scholar] [CrossRef]
- Kohorn, B.D. Cell wall-associated kinases and pectin perception. J. Exp. Bot. 2016, 67, 489–494. [Google Scholar] [CrossRef]
- Du, S.; Qu, L.-J.; Xiao, J. Crystal structures of the extracellular domains of the CrRLK1L receptor-like kinases ANXUR1 and ANXUR2. Protein Sci. 2018, 27, 886–892. [Google Scholar] [CrossRef]
- Moussu, S.; Augustin, S.; Roman, A.-O.; Broyart, C.; Santiago, J. Crystal structures of two tandem malectin-like receptor kinases involved in plant reproduction. Acta Crystallogr. Sect. D Struct. Biol. 2018, 74, 671–680. [Google Scholar] [CrossRef]
- Herger, A.; Gupta, S.; Kadler, G.; Franck, C.M.; Boisson-Dernier, A.; Ringli, C. Overlapping functions and protein-protein interactions of LRR-extensins in Arabidopsis. PLoS Genet. 2020, 16, e1008847. [Google Scholar] [CrossRef] [PubMed]
- Ringli, C. The hydroxyproline-rich glycoprotein domain of the Arabidopsis LRX1 requires Tyr for function but not for insolubilization in the cell wall. Plant J. 2010, 63, 662–669. [Google Scholar] [CrossRef]


{kind=link}
| AGI | Name | Molecular Function *) | Type of Interaction x) | Localization @) | Co-Expression §) R2 | ||
|---|---|---|---|---|---|---|---|
| FLA4 | FEI1 | FEI2 | |||||
| AT3G46550 | FLA4 | unknown | D + [27] | PM | 1 | 0.58 | 0.70 |
| AT2G35620 | FEI2 | co-receptor | D + [27] | PM | 0.70 | 0.42 | 1 |
| AT5G54380 | THE1 | RLK | D + [49] | PM | 0.59 | 0.55 | 0.33 |
| AT1G31420 | FEI1 | co-receptor | D + [27] | PM | 0.58 | 1 | 0.42 |
| AT5G64900 | PROPEP1 | PTI peptide | I − [49] | vacuole | 0.52 | 0.02 | 0.59 |
| AT5G53340 | HPGT1 1) | AGP biosynt. | n.d. | med-Golgi | 0.46 | 0.17 | 0.37 |
| AT5G09870 | CESA5 | seed coat secondary cell wall cellulose | I + [46] | PM | 0.46 | 0.81 | 0.34 |
| AT1G73080 | PEPR1 | receptor | I − [49] | PM | 0.44 | −0.02 | 0.45 |
| AT4G39350 | CesA2 1) | seed coat secondary cell wall cellulose | n.d. | PM | 0.44 | 0.57 | 0.63 |
| AT4G21060 | GALT2 | AGP biosynt. | D + [47] | Golgi | 0.42 | 0.09 | 0.35 |
| AT2G25300 | HPGT3 1) | AGP biosynt. | n.d. | Golgi | 0.41 | −0.05 | 0.35 |
| AT4G32410 | CesA1 1) | primary cell wall cellulose | n.d. | PM | 0.41 | 0.75 | 0.25 |
| AT5G05170 | CesA3 1) | primary cell wall cellulose | n.d. | PM | 0.37 | 0.69 | 0.36 |
| AT5G64740 | CesA6 | primary cell wall cellulose | I + [27] | PM | 0.37 | 0.68 | 0.23 |
| AT3G29810 | COBL2 | seed coat secondary cell wall cellulose | I + [46] | PM | 0.34 | −0.05 | 0.57 |
| AT1G17750 | PEPR2 | receptor | I − [49] | PM | 0.32 | 0.16 | 0.19 |
| AT4G22290 | SHOU4L2 | trafficking | I − [43] | PM | 0.24 | 0.10 | 0.57 |
| AT4G35920 | MCA1 | Ca2+ transport | D + [49] | PM | 0.24 | 0.17 | 0.17 |
| AT5G60920 | COB | primary cell wall cellulose | I + [27] | PM | 0.23 | 0.68 | 0.11 |
| AT4G32120 | HPGT2 1) | AGP biosynt. | n.d. | Golgi | 0.15 | -0.07 | 0.31 |
| At1g70560 | WEI8 | auxin level | I − [63] | cytosol | 0.08 | −0.25 | −0.05 |
| AT4G18780 | CesA8 1) | secondary cell wall cellulose | n.d. | PM | 0.08 | 0.35 | 0.22 |
| At4g24670 | TAR2 | auxin level | I − [63] | vac. | 0.07 | −0.33 | 0.28 |
| AT5G17420 | CesA7 1) | secondary cell wall cellulose | n.d. | PM | 0.07 | 0.33 | 0.22 |
| AT5G44030 | CesA4 1) | secondary cell wall cellulose | n.d. | PM | 0.05 | 0.32 | 0.20 |
| AT2G21770 | CesA9 1) | seed coat secondary cell wall cellulose | n.d. | PM | 0.05 | −0.13 | 0.01 |
| AT1G78880 | SHOU4 | trafficking | I − [43] | PM | 0.01 | −0.21 | 0.09 |
| AT5G64905 | PROPEP3 | PTI peptide | I − [49] | vac. | 0.00 | −0.09 | 0.12 |
| AT1G74800 | GALT5 | AGP biosynt. | D + [47] | Golgi | −0.02 | −0.13 | 0.00 |
| AT5G48870 | SAD1 | ABA signaling (neg. regulator) | I − [65] | cytosol, nuc. | −0.16 | −0.39 | 0.01 |
| AT4G21670 | CPL1 | ABA signaling (neg. regulator) | I − [65] | nuc. | −0.22 | -0.26 | −0.31 |
| AT1G24180 | IAR4 | auxin level | I − [63] | mitoch. | −0.23 | −0.26 | −0.33 |
| AT2G40220 | ABI4 | ABA signaling | I + [65] | nuc. | −0.28 | −0.20 | −0.20 |
| AT2G36270 | ABI5 | ABA signaling | I + [65] | nuc. | −0.29 | −0.3 | −0.35 |
| AT5G63800 | MUM2 | pectin in seed coat mucilage | D − [44] | apopl. | −0.31 | 0.17 | −0.39 |
| AT4G28370 | FLY1 | pectin in seed coat mucilage | D − [44] | unkown/novel compartment | −0.35 | −0.15 | −0.16 |
| AGI | Name | Molecular Function *) | Partner in Pathway | Expected Localization @) | Co-Expression §) R2 | ||
|---|---|---|---|---|---|---|---|
| FLA4 | FEI1 | FLA2 | |||||
| AT5G11890 | EMB3135 | unknown | FEI1, FEI2 [84] | PM | 0.71 | 0.35 | 0.61 |
| AT2G02780 | LRR IV | co-receptor | FEI1, FEI2 [83] | PM | 0.49 | 0.23 | 0.30 |
| AT1G34470 | unknown | unknown | FEI1 [84] | mt, cp | 0.38 | 0.49 | 0.36 |
| AT2G40316 | autophagy-like | unknown | FEI1 [84] | vac. | 0.36 | 0.21 | 0.419 |
| AT2G29180 | unknown | unknown | FEI1 [84] | cp | 0.28 | 0.551 | 0.179 |
| AT5G59650 | LRR I | receptor | FEI2 [84] | PM | 0.20 | 0.207 | 0.307 |
| AT5G49540 | Rab5-interacting fam. | trafficking | FEI1, FEI2 [84] | PM | 0.17 | −0.1 | 0.272 |
| AT5G06320 | NHL3 | unknown | FEI2 [84] | PM | 0.145 | 0.112 | −0.1 |
| AT3G28220 | TRAF-like fam. | unknown | FEI1 [84] | cp | 0.074 | −0.01 | -0.2 |
| AT1G47640 | seven transmembrane domain fam. | receptor-like | FEI1, FEI2 [84] | PM, apopl. | 0.07 | −0.17 | 0.19 |
| AT2G31360 | acyl-CoA desaturase like | enzyme | FEI1 [84] | ER | 0.06 | −0.06 | 0.42 |
| AT5G47530 | auxin-responsive fam. | unknown | FEI1 [84] | PM, apopl. | 0.05 | 0.16 | 0.39 |
| AT5G14030 | translocon-β | unknown | FEI1 [84] | ER | 0.01 | −0.25 | 0.087 |
| AT3G12180 | cornichon fam. | trafficking | FEI2 [84] | apopl. | 0.00 | −0.21 | 0.13 |
| AT4G25360 | TBL18 | O-acetyl transferase | FEI2 [84] | mt | 0.00 | 0.07 | −0.31 |
| AT4G14455 | BET12 | trafficking | FEI1, FEI2 [84] | Golgi | 0.00 | −0.1 | 0.08 |
| AT1G70520 | CRK2 | RLK | FEI1 [84] | PM | −0.03 | 0.25 | −0.20 |
| AT4G05370 | unknown | unknown | FEI1, FEI2 [84] | apopl., PM | −0.07 | −0.17 | −0.2 |
| AT5G65800 | ACS5 | ACC biosynt. | FEI1, FEI2 [27] | cytosol, PM | −0.10 | 0.02 | −0.03 |
| AT4G20790 | LRR VI | co-receptor | FEI2 [84] | PM | −0.10 | −0.18 | −0.18 |
| AT3G49700 | ACS9 | ACC biosyn. | FEI1, FEI2 [27] | cytosol, PM | −0.11 | −0.02 | −0.02 |
| AT5G47180 | VAMP fam. | trafficking | FEI2 [84] | ER | −0.16 | −0.15 | −0.26 |
| AT2G26180 | IQD6 | microtubule organization | FEI1, FEI2 [84] | nuc., cytosol | −0.18 | -0.29 | 0.15 |
| AT5G40640 | unknown | unknown | FEI1 [84] | PM | −0.18 | −0.37 | −0.11 |
| AT1G33100 | MATE efflux fam. | ion transport | FEI1 [84] | PM, vac. | −0.19 | −0.13 | −0.06 |
| AT4G23220 | CRK14 | RLK | FEI1 [84] | PM | −0.20 | −0.02 | −0.26 |
| AT1G17230 | LRR XI | receptor | FEI1 [79] | PM | −0.20 | 0.24 | −0.18 |
| AT1G21240 | WAK3 | RLK | FEI1, FEI2 [84] | PM | −0.23 | −0.07 | −0.29 |
| AT4G37680 | HHP4 | receptor-like | FEI1, FEI2 [84] | Golgi, vac., PM | −0.24 | −0.19 | −0.36 |
| AT2G41705 | CrcB fam. | fluoride transporter | FEI1, FEI2 [84] | PM | −0.30 | −0.23 | −0.31 |
| AT5G27350 | major facilitator fam. | sugar transporter | FEI1 [84] | PM, vac. | −0.30 | −0.08 | −0.35 |
| AT3G10640 | VPS60.1 | trafficking | FEI2 [84] | nuc. | −0.31 | −0.21 | −0.44 |
| AT1G17280 | UBC34 | ubiquitination | FEI1, FEI2 [84] | perox.,cytosol, | −0.34 | −0.19 | −0.48 |
| AT3G17000 | UBC32 | ubiquitination | FEI1, FEI2 [84] | cytosol | −0.35 | −0.29 | −0.47 |
| AT1G29060 | SFT12 | QcSNARE trafficking | FEI2 [84] | Golgi | −0.39 | −0.25 | −0.40 |
| AT2G04040 | MATE efflux fam. | ion transport | FEI1 [84] | PM | −0.40 | −0.30 | −0.39 |
| AT4G04860 | DER2.2 | ubiquitination | FEI1 [84] | ER, PM | −0.41 | −0.35 | −0.39 |
| AT3G66654 | CYP21-4 | ABA signaling | FEI1 [84] | Golgi | −0.5 | −0.13 | −0.56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seifert, G.J. The FLA4-FEI Pathway: A Unique and Mysterious Signaling Module Related to Cell Wall Structure and Stress Signaling. Genes 2021, 12, 145. https://doi.org/10.3390/genes12020145
Seifert GJ. The FLA4-FEI Pathway: A Unique and Mysterious Signaling Module Related to Cell Wall Structure and Stress Signaling. Genes. 2021; 12(2):145. https://doi.org/10.3390/genes12020145
Chicago/Turabian StyleSeifert, Georg J. 2021. "The FLA4-FEI Pathway: A Unique and Mysterious Signaling Module Related to Cell Wall Structure and Stress Signaling" Genes 12, no. 2: 145. https://doi.org/10.3390/genes12020145
APA StyleSeifert, G. J. (2021). The FLA4-FEI Pathway: A Unique and Mysterious Signaling Module Related to Cell Wall Structure and Stress Signaling. Genes, 12(2), 145. https://doi.org/10.3390/genes12020145