
Participation of TFIIIB Subunit Brf1 in Transcription Regulation in the Human Pathogen Leishmania major


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatic Analyses
2.2. Cell Culture and Transfection
2.3. Generation of Plasmids
2.4. Indirect Immunofluorescence Assays
2.5. Western Blot Analysis
2.6. Chromatin Immunoprecipitation Assays
2.7. Quantitative Real-Time PCR Experiments
2.8. Northern and Southern Blot Analyses
2.9. Tandem Affinity Purifications and Mass Spectrometry Analysis
3. Results
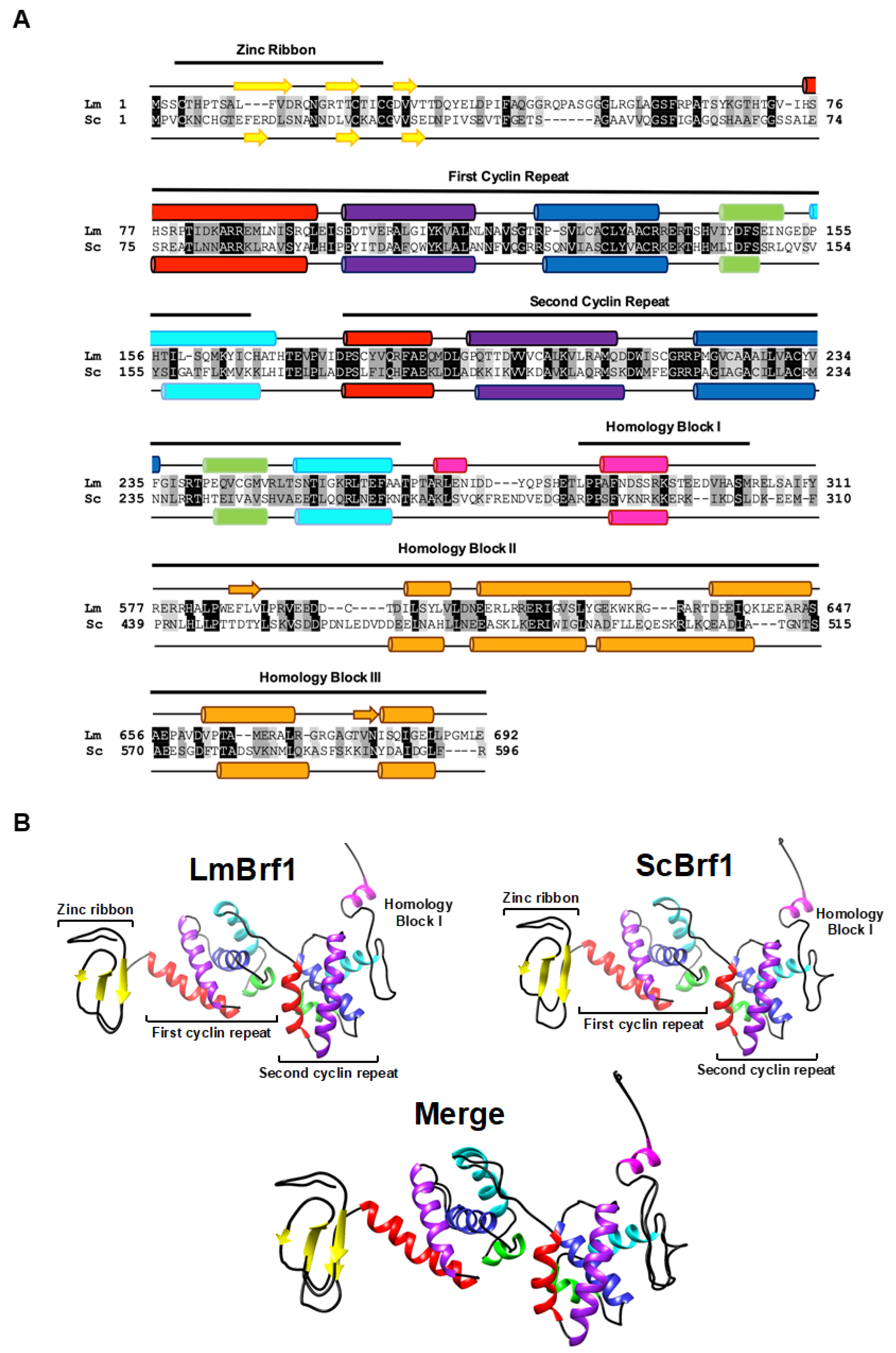
3.1. The Sequence and Predicted Structure of LmBrf1 Are Conserved
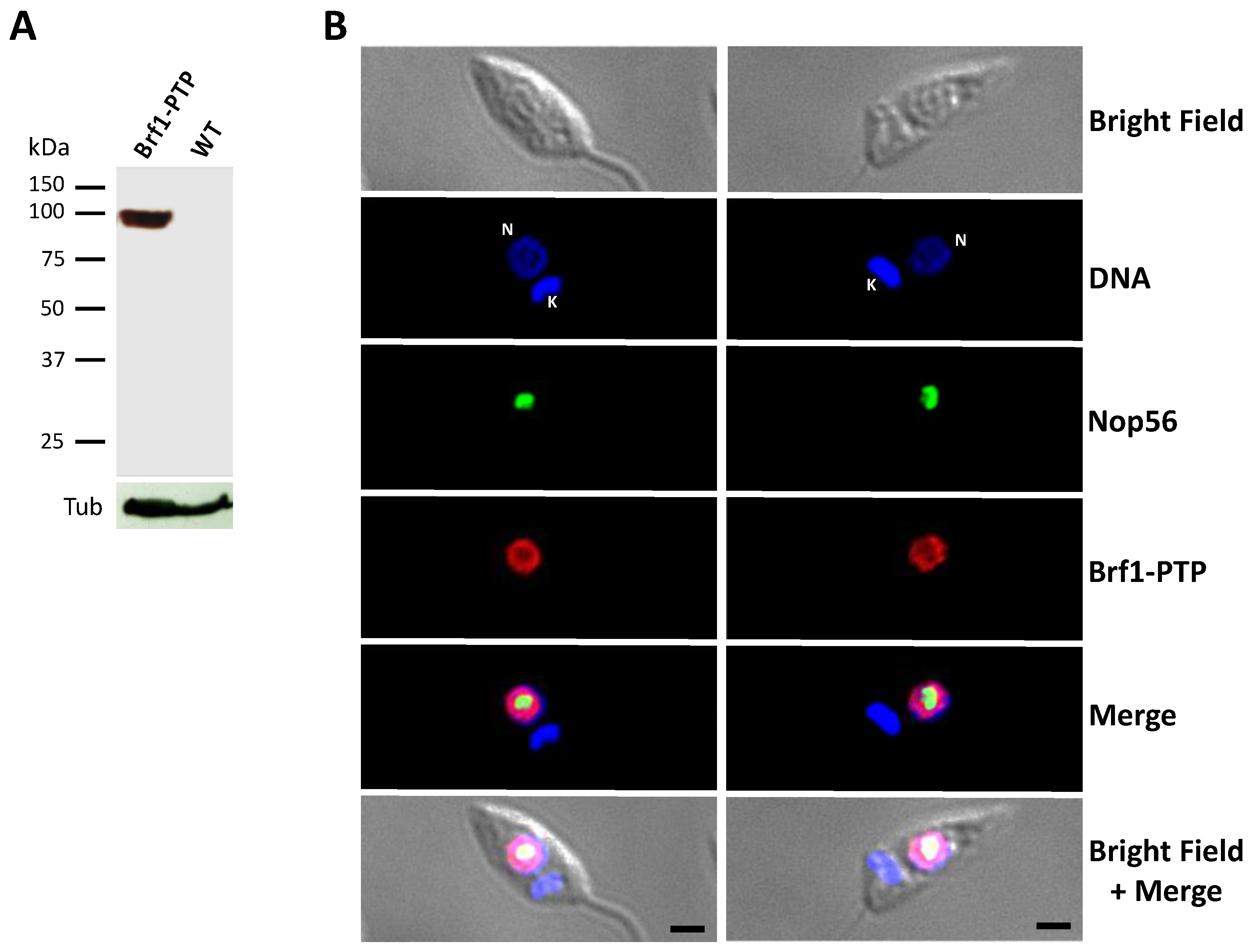
3.2. LmBRF1 Is a Nuclear Protein
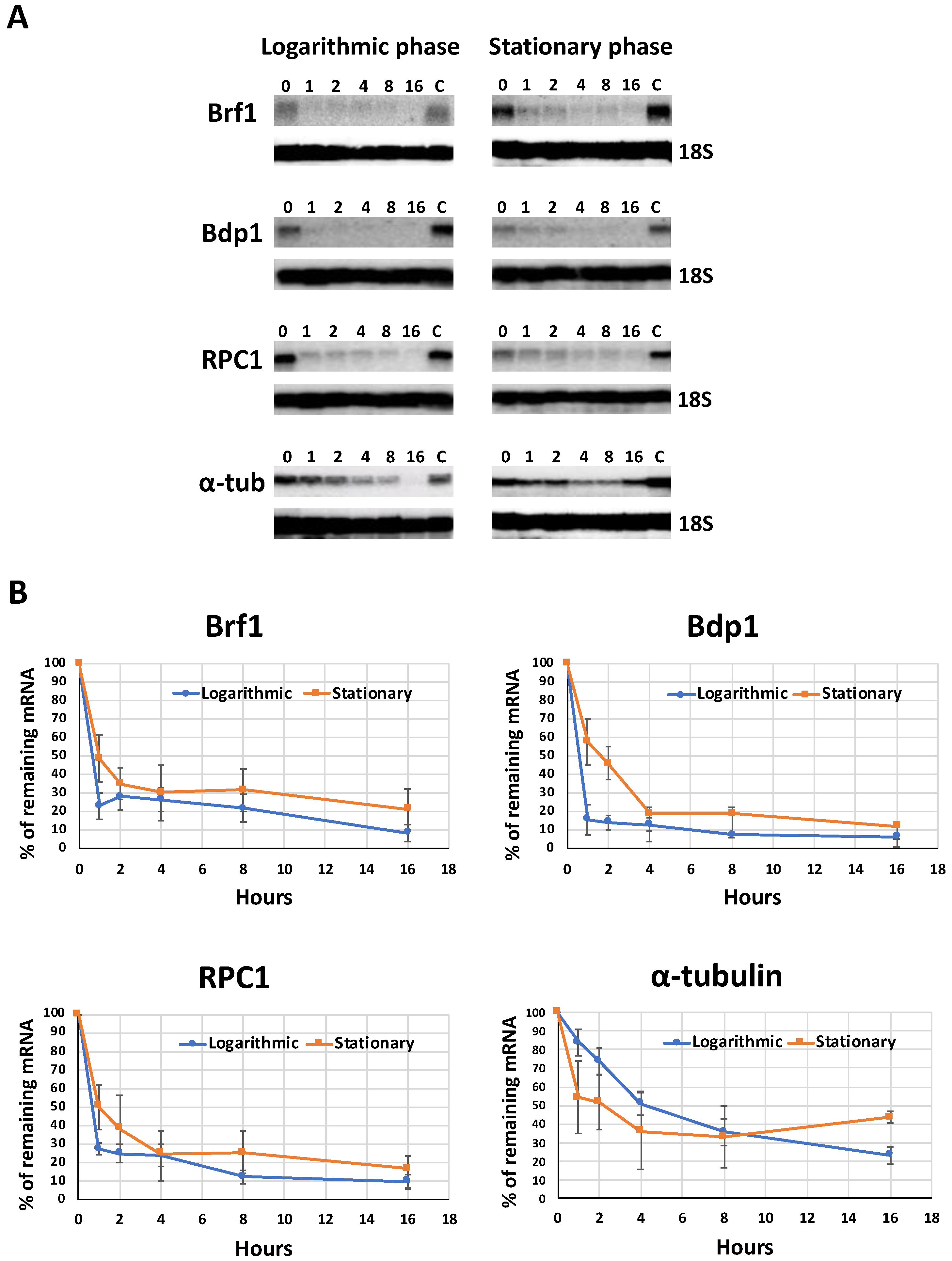
3.3. The Half-Lives of the mRNAs from LmBrf1 and Other Components of the Pol III Transcription Machinery Are Very Similar
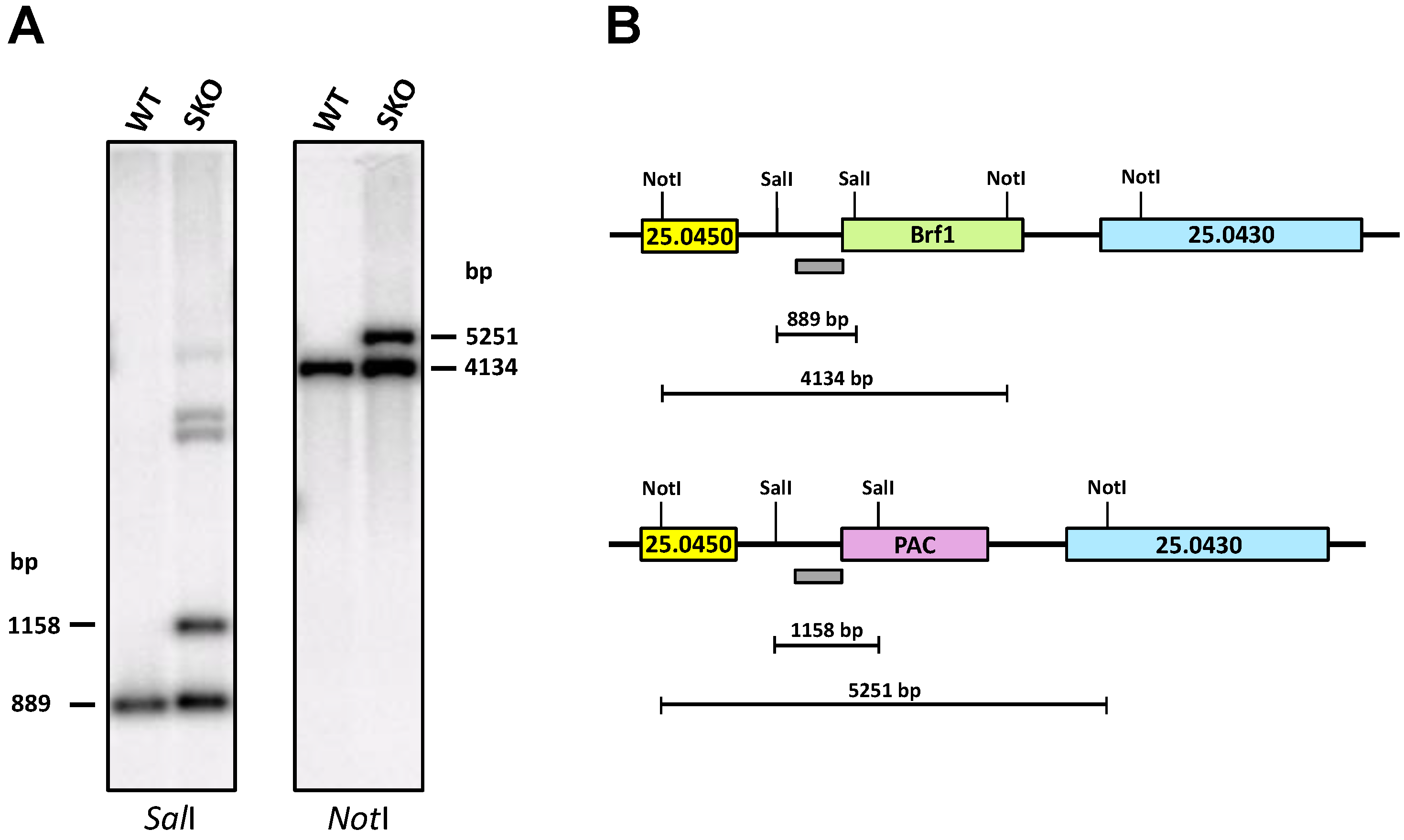
3.4. Promastigotes Need at Least One Copy of the LmBrf1 Gene to Survive
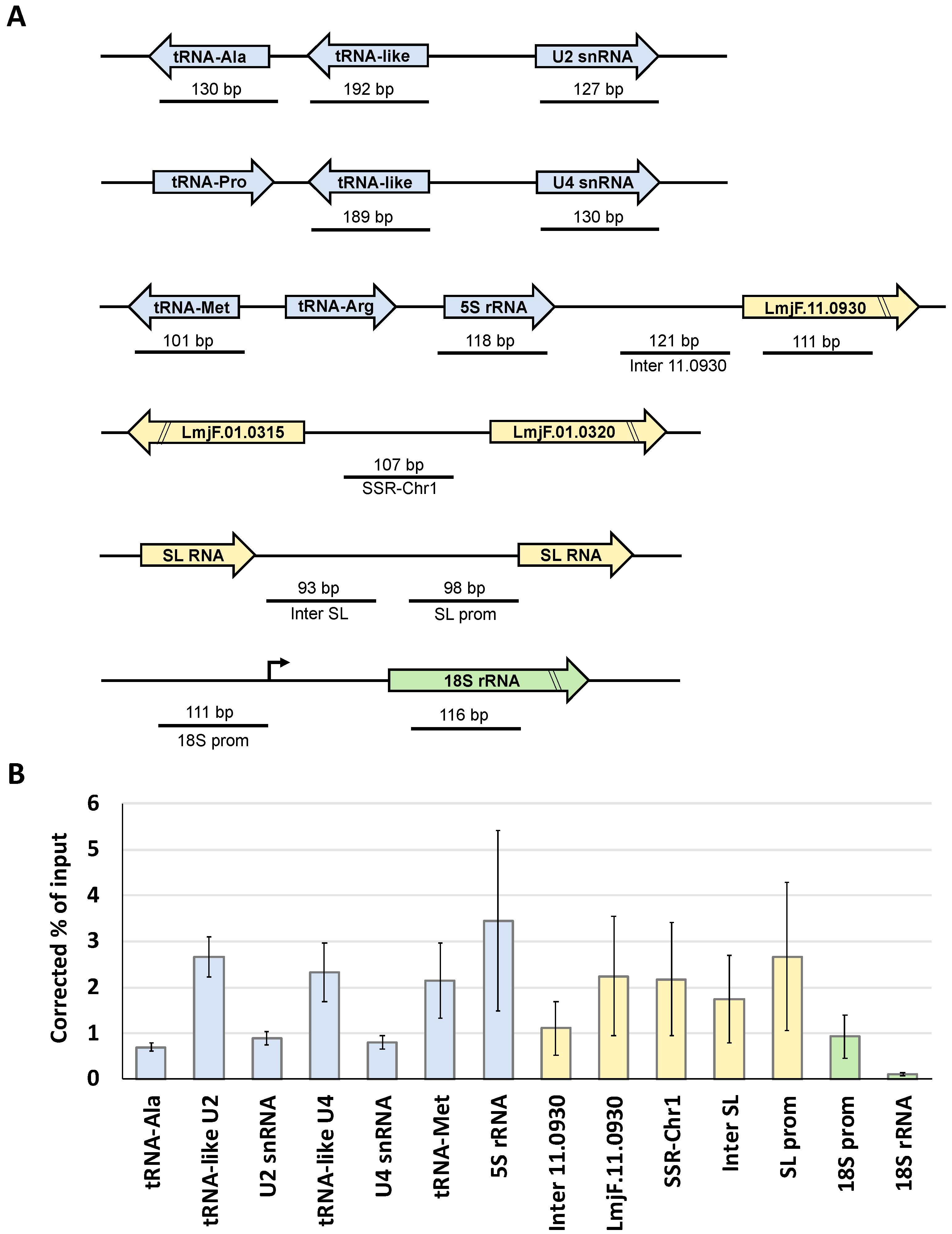
3.5. LmBrf1 Binds to Pol III Promoters and Other Genomic Regions
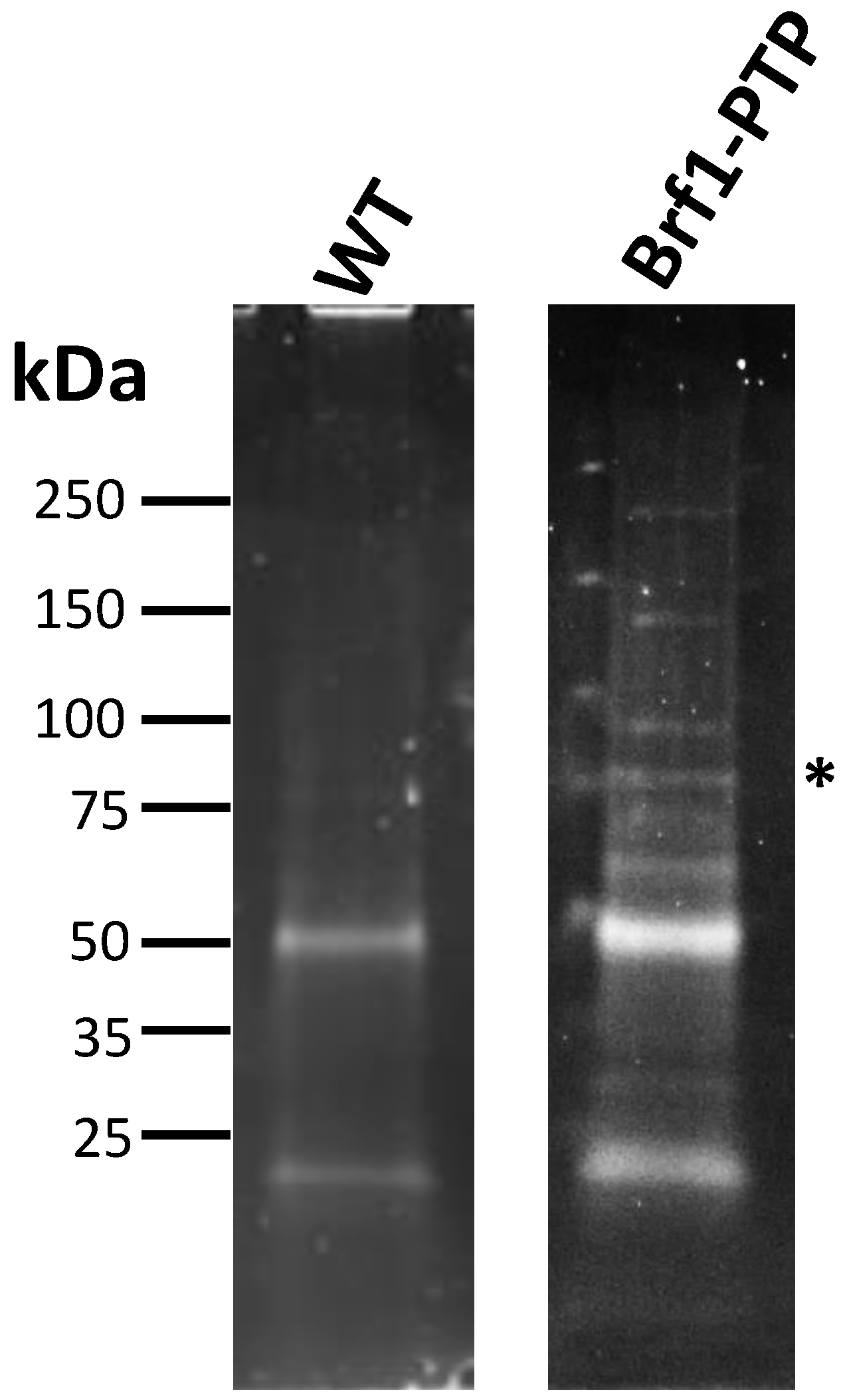
3.6. Proteins That Participate in Pol I, Pol II, and Pol III Transcription Were Co-Purified with LmBrf1-PTP
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tuon, F.F.; Neto, V.A.; Amato, V.S. Leishmania: Origin, evolution and future since the Precambrian. FEMS Immunol. Med. Microbiol. 2008, 54, 158–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunzl, A.; Vanhamme, L.; Myler, P.J. Transcription in trypanosomes: A different means to the end. In Trypanosomes: After the Genome; Barry, J.D., McCulloch, R., Mottram, J.C., Acosta-Serrano, A., Eds.; Horizon Bioscience: Wymonham, UK, 2007; pp. 177–208. [Google Scholar]
- Martinez-Calvillo, S.; Vizuet-de-Rueda, J.C.; Florencio-Martinez, L.E.; Manning-Cela, R.G.; Figueroa-Angulo, E.E. Gene expression in trypanosomatid parasites. J. Biomed. Biotechnol. 2010, 2010, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clayton, C. The Regulation of Trypanosome Gene Expression by RNA-Binding Proteins. PLoS Pathog. 2013, 9, e1003680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roeder, R.G. 50+ years of eukaryotic transcription: An expanding universe of factors and mechanisms. Nat. Struct. Mol. Biol. 2019, 26, 783–791. [Google Scholar] [CrossRef]
- Grummt, I. Life on a planet of its own: Regulation of RNA polymerase I transcription in the nucleolus. Genes Dev. 2003, 17, 1691–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Bushnell, D.A.; Kornberg, R.D. RNA polymerase II transcription: Structure and mechanism. Biochim. Biophys. Acta 2013, 1829, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Dieci, G.; Fiorino, G.; Castelnuovo, M.; Teichmann, M.; Pagano, A. The expanding RNA polymerase III transcriptome. Trends Genet. 2007, 23, 614–622. [Google Scholar] [CrossRef]
- Lesniewska, E.; Boguta, M. Novel layers of RNA polymerase III control affecting tRNA gene transcription in eukaryotes. Open Biol. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Dieci, G.; Bosio, M.C.; Fermi, B.; Ferrari, R. Transcription reinitiation by RNA polymerase III. Biochim. Biophys. Acta 2013, 1829, 331–341. [Google Scholar] [CrossRef]
- Geiduschek, E.P.; Kassavetis, G.A. The RNA polymerase III transcription apparatus. J. Mol. Biol. 2001, 310, 1–26. [Google Scholar] [CrossRef]
- Kassavetis, G.A.; Han, S.; Naji, S.; Geiduschek, E.P. The role of transcription initiation factor IIIB subunits in promoter opening probed by photochemical cross-linking. J. Biol. Chem. 2003, 278, 17912–17917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huet, J.; Conesa, C.; Manaud, N.; Chaussivert, N.; Sentenac, A. Interactions between yeast TFIIIB components. Nucleic Acids Res. 1994, 22, 3433–3439. [Google Scholar] [CrossRef] [Green Version]
- Schramm, L.; Pendergrast, P.S.; Sun, Y.; Hernandez, N. Different human TFIIIB activities direct RNA polymerase III transcription from TATA-containing and TATA-less promoters. Genes Dev. 2000, 14, 2650–2663. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Yang, W.; Yu, H.; Fu, C.; Liu, X.; Liu, J. Double mutation of BRF1 and BRF2 leads to sterility in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2019, 516, 969–975. [Google Scholar] [CrossRef]
- Vannini, A.; Cramer, P. Conservation between the RNA polymerase I, II, and III transcription initiation machineries. Mol. Cell 2012, 45, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Lopez-de-Leon, A.; Librizzi, M.; Puglia, K.; Willis, I.M. PCF4 encodes an RNA polymerase III transcription factor with homology to TFIIB. Cell 1992, 71, 211–220. [Google Scholar] [CrossRef]
- Khoo, S.K.; Wu, C.C.; Lin, Y.C.; Lee, J.C.; Chen, H.T. Mapping the protein interaction network for TFIIB-related factor Brf1 in the RNA polymerase III preinitiation complex. Mol. Cell Biol. 2014, 34, 551–559. [Google Scholar] [PubMed] [Green Version]
- Hsieh, Y.J.; Kundu, T.K.; Wang, Z.; Kovelman, R.; Roeder, R.G. The TFIIIC90 subunit of TFIIIC interacts with multiple components of the RNA polymerase III machinery and contains a histone-specific acetyltransferase activity. Mol. Cell Biol. 1999, 19, 7697–7704. [Google Scholar] [PubMed] [Green Version]
- Hsieh, Y.J.; Wang, Z.; Kovelman, R.; Roeder, R.G. Cloning and characterization of two evolutionarily conserved subunits (TFIIIC102 and TFIIIC63) of human TFIIIC and their involvement in functional interactions with TFIIIB and RNA polymerase III. Mol. Cell Biol. 1999, 19, 4944–4952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moir, R.D.; Puglia, K.V.; Willis, I.M. A gain-of-function mutation in the second tetratricopeptide repeat of TFIIIC131 relieves autoinhibition of Brf1 binding. Mol. Cell Biol. 2002, 22, 6131–6141. [Google Scholar] [CrossRef] [Green Version]
- Khoo, S.K.; Wu, C.C.; Lin, Y.C.; Chen, H.T. The TFIIE-related Rpc82 subunit of RNA polymerase III interacts with the TFIIB-related transcription factor Brf1 and the polymerase cleft for transcription initiation. Nucleic Acids Res. 2018, 46, 1157–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abascal-Palacios, G.; Ramsay, E.P.; Beuron, F.; Morris, E.; Vannini, A. Structural basis of RNA polymerase III transcription initiation. Nature 2018, 553, 301–306. [Google Scholar] [CrossRef]
- Vorlander, M.K.; Khatter, H.; Wetzel, R.; Hagen, W.J.H.; Muller, C.W. Molecular mechanism of promoter opening by RNA polymerase III. Nature 2018, 553, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Vorlander, M.K.; Jungblut, A.; Karius, K.; Baudin, F.; Grotsch, H.; Kosinski, J.; Muller, C.W. Structure of the TFIIIC subcomplex tauA provides insights into RNA polymerase III pre-initiation complex formation. Nat. Commun. 2020, 11, 4905. [Google Scholar] [CrossRef] [PubMed]
- Fantoni, A.; Dare, A.O.; Tschudi, C. RNA polymerase III-mediated transcription of the trypanosome U2 small nuclear RNA gene is controlled by both intragenic and extragenic regulatory elements. Mol. Cell Biol. 1994, 14, 2021–2028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas-Sanchez, S.; Figueroa-Angulo, E.; Moreno-Campos, R.; Florencio-Martinez, L.E.; Manning-Cela, R.G.; Martinez-Calvillo, S. Transcription of Leishmania major U2 small nuclear RNA gene is directed by extragenic sequences located within a tRNA-like and a tRNA-Ala gene. Parasit. Vectors 2016, 9, 401. [Google Scholar]
- Padilla-Mejia, N.E.; Florencio-Martinez, L.E.; Figueroa-Angulo, E.E.; Manning-Cela, R.G.; Hernandez-Rivas, R.; Myler, P.J.; Martinez-Calvillo, S. Gene organization and sequence analyses of transfer RNA genes in Trypanosomatid parasites. BMC Genom. 2009, 10, 232. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Campos, R.; Florencio-Martinez, L.E.; Nepomuceno-Mejia, T.; Rojas-Sanchez, S.; Velez-Ramirez, D.E.; Padilla-Mejia, N.E.; Figueroa-Angulo, E.; Manning-Cela, R.; Martinez-Calvillo, S. Molecular characterization of 5S ribosomal RNA genes and transcripts in the protozoan parasite Leishmania major. Parasitology 2016, 143, 1917–1929. [Google Scholar] [CrossRef]
- Nakaar, V.; Gunzl, A.; Ullu, E.; Tschudi, C. Structure of the Trypanosoma brucei U6 snRNA gene promoter. Mol. Biochem. Parasitol. 1997, 88, 13–23. [Google Scholar] [CrossRef]
- Thomas, S.; Yu, M.C.; Sturm, N.R.; Campbell, D.A. A non-universal transcription factor? The Leishmania tarentolae TATA box-binding protein LtTBP associates with a subset of promoters. Int. J. Parasitol. 2006, 36, 1217–1226. [Google Scholar] [CrossRef]
- Thomas, S.; Green, A.; Sturm, N.R.; Campbell, D.A.; Myler, P.J. Histone acetylations mark origins of polycistronic transcription in Leishmania major. BMC Genom. 2009, 10, 152. [Google Scholar] [CrossRef] [Green Version]
- Roman-Carraro, F.C.; Florencio-Martinez, L.E.; Romero-Meza, G.; Nepomuceno-Mejia, T.; Carrero, J.C.; Arroyo, R.; Ortega-Lopez, J.; Manning-Cela, R.G.; Martinez-Calvillo, S. TFIIIB Subunit Bdp1 Participates in RNA Polymerase III Transcription in the Protozoan Parasite Leishmania major. BioMed Res. Int. 2019, 2019, 1425281. [Google Scholar] [CrossRef] [Green Version]
- Schimanski, B.; Nguyen, T.N.; Gunzl, A. Characterization of a multisubunit transcription factor complex essential for spliced-leader RNA gene transcription in Trypanosoma brucei. Mol. Cell Biol. 2005, 25, 7303–7313. [Google Scholar] [CrossRef] [Green Version]
- Velez-Ramirez, D.E.; Florencio-Martinez, L.E.; Romero-Meza, G.; Rojas-Sanchez, S.; Moreno-Campos, R.; Arroyo, R.; Ortega-Lopez, J.; Manning-Cela, R.; Martinez-Calvillo, S. BRF1, a subunit of RNA polymerase III transcription factor TFIIIB, is essential for cell growth of Trypanosoma brucei. Parasitology 2015, 142, 1563–1573. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, L.; Stephens, A.; Nam, S.Z.; Rau, D.; Kubler, J.; Lozajic, M.; Gabler, F.; Soding, J.; Lupas, A.N.; Alva, V. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at its Core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Calvillo, S.; Stuart, K.; Myler, P.J. Ploidy changes associated with disruption of two adjacent genes on Leishmania major chromosome 1. Int. J. Parasitol. 2005, 35, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Nepomuceno-Mejia, T.; Florencio-Martinez, L.E.; Martinez-Calvillo, S. Nucleolar Division in the Promastigote Stage of Leishmania major Parasite: A Nop56 Point of View. BioMed Res. Int. 2018, 2018, 1641839. [Google Scholar] [CrossRef]
- Ji, H. Lysis of cultured cells for immunoprecipitation. Cold Spring Harb. Protoc. 2010, 2010. [Google Scholar] [CrossRef]
- Romero-Meza, G.; Velez-Ramirez, D.E.; Florencio-Martinez, L.E.; Roman-Carraro, F.C.; Manning-Cela, R.; Hernandez-Rivas, R.; Martinez-Calvillo, S. Maf1 is a negative regulator of transcription in Trypanosoma brucei. Mol. Microbiol. 2017, 103, 452–468. [Google Scholar] [CrossRef] [Green Version]
- Vizuet-de-Rueda, J.C.; Florencio-Martinez, L.E.; Padilla-Mejia, N.E.; Manning-Cela, R.; Hernandez-Rivas, R.; Martinez-Calvillo, S. Ribosomal RNA Genes in the Protozoan Parasite Leishmania major Possess a Nucleosomal Structure. Protist 2016, 167, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Schimanski, B.; Nguyen, T.N.; Gunzl, A. Highly efficient tandem affinity purification of trypanosome protein complexes based on a novel epitope combination. Eukaryot. Cell 2005, 4, 1942–1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Calvillo, S.; Saxena, A.; Green, A.; Leland, A.; Myler, P.J. Characterization of the RNA polymerase II and III complexes in Leishmania major. Int. J. Parasitol. 2007, 37, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juo, Z.S.; Kassavetis, G.A.; Wang, J.; Geiduschek, E.P.; Sigler, P.B. Crystal structure of a transcription factor IIIB core interface ternary complex. Nature 2003, 422, 534–539. [Google Scholar] [CrossRef]
- Chen, H.T.; Legault, P.; Glushka, J.; Omichinski, J.G.; Scott, R.A. Structure of a (Cys3His) zinc ribbon, a ubiquitous motif in archaeal and eucaryal transcription. Protein Sci. 2000, 9, 1743–1752. [Google Scholar] [CrossRef] [Green Version]
- Noble, M.E.; Endicott, J.A.; Brown, N.R.; Johnson, L.N. The cyclin box fold: Protein recognition in cell-cycle and transcription control. Trends Biochem. Sci. 1997, 22, 482–487. [Google Scholar] [CrossRef]
- Rogers, M.B.; Hilley, J.D.; Dickens, N.J.; Wilkes, J.; Bates, P.A.; Depledge, D.P.; Harris, D.; Her, Y.; Herzyk, P.; Imamura, H.; et al. Chromosome and gene copy number variation allow major structural change between species and strains of Leishmania. Genome Res. 2011, 21, 2129–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Cai, G.; Panigrahi, A.K.; Dunham-Ems, S.; Nguyen, T.N.; Radolf, J.D.; Asturias, F.J.; Gunzl, A. A TFIIH-associated mediator head is a basal factor of small nuclear spliced leader RNA gene transcription in early-diverged trypanosomes. Mol. Cell Biol. 2010, 30, 5502–5513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Calvillo, S.; Yan, S.; Nguyen, D.; Fox, M.; Stuart, K.; Myler, P.J. Transcription of Leishmania major Friedlin chromosome 1 initiates in both directions within a single region. Mol. Cell 2003, 11, 1291–1299. [Google Scholar] [CrossRef]
- Mellacheruvu, D.; Wright, Z.; Couzens, A.L.; Lambert, J.P.; St-Denis, N.A.; Li, T.; Miteva, Y.V.; Hauri, S.; Sardiu, M.E.; Low, T.Y.; et al. The CRAPome: A contaminant repository for affinity purification-mass spectrometry data. Nat. Methods 2013, 10, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Kassavetis, G.A.; Bartholomew, B.; Blanco, J.A.; Johnson, T.E.; Geiduschek, E.P. Two essential components of the Saccharomyces cerevisiae transcription factor TFIIIB: Transcription and DNA-binding properties. Proc. Natl. Acad. Sci. USA 1991, 88, 7308–7312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabani, M.; Levin, J.Z.; Fan, L.; Adiconis, X.; Raychowdhury, R.; Garber, M.; Gnirke, A.; Nusbaum, C.; Hacohen, N.; Friedman, N.; et al. Metabolic labeling of RNA uncovers principles of RNA production and degradation dynamics in mammalian cells. Nat. Biotechnol. 2011, 29, 436–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadda, A.; Ryten, M.; Droll, D.; Rojas, F.; Farber, V.; Haanstra, J.R.; Merce, C.; Bakker, B.M.; Matthews, K.; Clayton, C. Transcriptome-wide analysis of trypanosome mRNA decay reveals complex degradation kinetics and suggests a role for co-transcriptional degradation in determining mRNA levels. Mol. Microbiol. 2014, 94, 307–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharova, L.V.; Sharov, A.A.; Nedorezov, T.; Piao, Y.; Shaik, N.; Ko, M.S. Database for mRNA half-life of 19 977 genes obtained by DNA microarray analysis of pluripotent and differentiating mouse embryonic stem cells. DNA Res. 2009, 16, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Cevallos, A.M.; Perez-Escobar, M.; Espinosa, N.; Herrera, J.; Lopez-Villasenor, I.; Hernandez, R. The stabilization of housekeeping transcripts in Trypanosoma cruzi epimastigotes evidences a global regulation of RNA decay during stationary phase. FEMS Microbiol. Lett. 2005, 246, 259–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coulson, R.M.; Connor, V.; Chen, J.C.; Ajioka, J.W. Differential expression of Leishmania major β-tubulin genes during the acquisition of promastigote infectivity. Mol. Biochem. Parasitol. 1996, 82, 227–236. [Google Scholar] [CrossRef]
- Brittingham, A.; Miller, M.A.; Donelson, J.E.; Wilson, M.E. Regulation of GP63 mRNA stability in promastigotes of virulent and attenuated Leishmania chagasi. Mol. Biochem. Parasitol. 2001, 112, 51–59. [Google Scholar] [PubMed]
- Gouge, J.; Guthertz, N.; Kramm, K.; Dergai, O.; Abascal-Palacios, G.; Satia, K.; Cousin, P.; Hernandez, N.; Grohmann, D.; Vannini, A. Molecular mechanisms of Bdp1 in TFIIIB assembly and RNA polymerase III transcription initiation. Nat. Commun. 2017, 8, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivens, A.C.; Peacock, C.S.; Worthey, E.A.; Murphy, L.; Aggarwal, G.; Berriman, M.; Sisk, E.; Rajandream, M.A.; Adlem, E.; Aert, R.; et al. The genome of the kinetoplastid parasite, Leishmania major. Science 2005, 309, 436–442. [Google Scholar] [CrossRef] [Green Version]
- Collart, M.A. The Ccr4-Not complex is a key regulator of eukaryotic gene expression. Wiley Interdiscip. Rev. RNA 2016, 7, 438–454. [Google Scholar] [CrossRef] [Green Version]
- Schwede, A.; Ellis, L.; Luther, J.; Carrington, M.; Stoecklin, G.; Clayton, C. A role for Caf1 in mRNA deadenylation and decay in trypanosomes and human cells. Nucleic Acids Res. 2008, 36, 3374–3388. [Google Scholar] [CrossRef] [Green Version]
- Ouna, B.A.; Nyambega, B.; Manful, T.; Helbig, C.; Males, M.; Fadda, A.; Clayton, C. Depletion of trypanosome CTR9 leads to gene expression defects. PLoS ONE 2012, 7, e34256. [Google Scholar] [CrossRef] [Green Version]
- Bhalla, P.; Shukla, A.; Vernekar, D.V.; Arimbasseri, A.G.; Sandhu, K.S.; Bhargava, P. Yeast PAF1 complex counters the pol III accumulation and replication stress on the tRNA genes. Sci. Rep. 2019, 9, 12892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohdate, H.; Lim, C.R.; Kokubo, T.; Matsubara, K.; Kimata, Y.; Kohno, K. Impairment of the DNA binding activity of the TATA-binding protein renders the transcriptional function of Rvb2p/Tih2p, the yeast RuvB-like protein, essential for cell growth. J. Biol. Chem. 2003, 278, 14647–14656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez-Beltran, E.; Denisenko, T.V.; Zhivotovsky, B.; Bozhkov, P.V. Tudor staphylococcal nuclease: Biochemistry and functions. Cell Death. Differ. 2016, 23, 1739–1748. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, M.S.; Rudenko, G. TDP1 is an HMG chromatin protein facilitating RNA polymerase I transcription in African trypanosomes. Nucleic Acids Res. 2013, 41, 2981–2992. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.H.; Sharrocks, A.D.; Whitmarsh, A.J. MAP kinase signalling cascades and transcriptional regulation. Gene 2013, 513, 1–13. [Google Scholar] [CrossRef]
- Lee, J.; Moir, R.D.; Willis, I.M. Differential Phosphorylation of RNA Polymerase III and the Initiation Factor TFIIIB in Saccharomyces cerevisiae. PLoS ONE 2015, 10, e0127225. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Moscinski, L.C. Cdc2: A monopotent or pluripotent CDK? Cell Prolif. 2011, 44, 205–211. [Google Scholar] [PubMed]
- Chen, Y.H.; Keegan, S.; Kahli, M.; Tonzi, P.; Fenyo, D.; Huang, T.T.; Smith, D.J. Transcription shapes DNA replication initiation and termination in human cells. Nat. Struct. Mol. Biol. 2019, 26, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Raha, D.; Wang, Z.; Moqtaderi, Z.; Wu, L.; Zhong, G.; Gerstein, M.; Struhl, K.; Snyder, M. Close association of RNA polymerase II and many transcription factors with Pol III genes. Proc. Natl. Acad. Sci. USA 2010, 107, 3639–3644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, F.N.; Canella, D.; Praz, V.; Michaud, J.; Romascano, D.; Hernandez, N. Genomic study of RNA polymerase II and III SNAPc-bound promoters reveals a gene transcribed by both enzymes and a broad use of common activators. PLoS Genet. 2012, 8, e1003028. [Google Scholar]
- Johnson, S.S.; Zhang, C.; Fromm, J.; Willis, I.M.; Johnson, D.L. Mammalian Maf1 is a negative regulator of transcription by all three nuclear RNA polymerases. Mol. Cell 2007, 26, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Gerber, A.; Ito, K.; Chu, C.S.; Roeder, R.G. Gene-Specific Control of tRNA Expression by RNA Polymerase II. Mol. Cell 2020, 78, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Abraham, K.J.; Khosraviani, N.; Chan, J.N.Y.; Gorthi, A.; Samman, A.; Zhao, D.Y.; Wang, M.; Bokros, M.; Vidya, E.; Ostrowski, L.A.; et al. Nucleolar RNA polymerase II drives ribosome biogenesis. Nature 2020, 585, 298–302. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TriTrypDB Name | Protein Function (Known or Putative) | Predicted Size(kDa) | Peptides b |
|---|---|---|---|
| Transcription factors | |||
| LmjF.19.1390 | TATA-binding protein | 31.1 | 43 (3) |
| LmjF.33.2810 | Transcription elongation factor TFIIS | 50.5 | 5 (1) |
| LmjF.12.0560 | TFIIIC subunit Tau131, putative | 149 | 1 (1) |
| LmjF.33.2820 | TFIIS-like | 63.9 | 1 (1) |
| LmjF.33.3090 | Transcription elongation factor SPT6 | 158 | 1 (1) |
| RNA polymerase subunits | |||
| LmjF.19.0660 | RPAC1 (Pol I and Pol III) | 47.3 | 6 (2) |
| LmjF.31.0160 | RPB2 (Pol II) | 133.8 | 6 (2) |
| LmjF.16.1350 | RPA1 (Pol I) | 199.7 | 4 (2) |
| LmjF.18.0780 | RPB5 (Pol II and Pol III) | 27.2 | 2 (2) |
| LmjF.34.0360 | RPC1 (Pol III) | 173.6 | 1 (1) |
| LmjF.18.0790 | RPB5z (Pol I) | 37.6 | 1 (1) |
| LmjF.31.2610 | RPB1 (Pol II) | 184.5 | 1 (1) |
| Regulators of transcription and/or chromatin remodelers | |||
| LmjF.34.2610 | RuvB-like DNA helicase, putative | 53.6 | 11 (3) |
| LmjF29.0850 | High-mobility group protein TDP1 | 33.6 | 8 (2) |
| LmjF.35.2850 | PAF1 complex subunit LEO1 | 61.7 | 8 (2) |
| LmjF.29.1110 | PAF1 complex novel subunit | 68.4 | 7 (2) |
| LmjF.14.0890 | PAF1 complex novel subunit (putative RTF1) | 71.8 | 7 (3) |
| LmjF.19.0440 | Nucleosome assembly protein, putative | 39.7 | 5 (2) |
| LmjF.32.0950 | Staphylococcal nuclease homolog/Tudor domain-containing protein, putative | 102.3 | 5 (2) |
| LmjF29.0020 | FACT complex subunit SPT16 | 114.7 | 4 (2) |
| LmjF.31.1750 | Nucleosome assembly protein-like | 45.4 | 3 (1) |
| LmjF.29.2340 | Nucleosome assembly protein (NAP), putative | 21.2 | 2 (2) |
| LmjF.29.2550 | PAF1 complex subunit CTR9 | 96.6 | 2 (2) |
| LmjF.22.0450 | Acetyltransferase (GNAT) family | 32.3 | 1 (1) |
| LmjF.25.1840 | CCR4-NOT transcription complex subunit Not5 | 72.4 | 1 (1) |
| LmjF.21.0800 | CCR4-NOT transcription complex subunit Not1 | 248.7 | 1 (1) |
| DNA or RNA binding proteins | |||
| LmjF27.1300 | KH domain-containing protein, putative | 59.9 | 13 (2) |
| LmjF.21.1552 | ATP-dependent RNA helicase SUB2, putative | 49.5 | 12 (3) |
| LmjF.21.0540 | La protein homolog | 37.2 | 11 (3) |
| LmjF.34.2580 | ALBA-domain protein 3 | 22.6 | 6 (3) |
| LmjF.06.0010 | Histone H4 | 11.4 | 6 (3) |
| LmjF.28.1530 | ATP-dependent RNA helicase FAL1, putative | 44 | 6 (3) |
| LmjF.07.1000 | RNA-binding protein-like protein | 35 | 5 (3) |
| LmjF.18.0700 | HEAT repeats, putative | 77.3 | 5 (3) |
| LmjF.30.3090 | RNA-binding protein 42 (RNA-binding motif protein 42), putative | 36.3 | 5 (2) |
| LmjF.30.3430 | Protein Mkt1, putative | 90.2 | 4 (2) |
| LmjF.12.1220 | WD repeat and HMG-box DNA-binding protein, putative | 150.2 | 4 (1) |
| LmjF.24.1490 | WD domain, G-beta repeat, putative | 47.5 | 3 (1) |
| LmjF.22.0470 | tRNA-binding domain-containing protein | 44.8 | 2 (1) |
| DNA replication | |||
| LmjF.28.0850 | DNA replication licensing factor MCM2, putative | 110.4 | 7 (1) |
| LmjF.09.0250 | DNA replication licensing factor MCM4, putative | 97.2 | 5 (1) |
| LmjF27.0550 | Replication factor C, subunit 4, putative | 39.5 | 5 (2) |
| LmjF.15.1450 | Proliferative cell nuclear antigen (PCNA), putative | 32.4 | 4 (2) |
| LmjF.36.6710 | Replication factor C subunit 3, putative | 39.3 | 3 (2) |
| Kinases or phosphatases | |||
| LmjF.32.2950 | Nucleoside diphosphate kinase B | 16.6 | 14 (3) |
| LmjF.25.0750 | Protein phosphatase 2C | 44.9 | 6 (3) |
| LmjF.10.0200 | Mitogen-activated protein kinase 10, putative | 46.3 | 2 (2) |
| LmjF.36.0550 | Cdc2-related kinase 3 | 35.6 | 2 (1) |
| LmjF.34.2820 | Regulatory subunit of protein kinase A-like | 71.6 | 2 (1) |
| Other functions | |||
| LmjF.08.1110 | Stress-induced protein STI1 | 62.1 | 20 (3) |
| LmjF.36.3210 | 14-3-3 protein 1, putative | 29.7 | 15 (3) |
| LmjF.17.0870 | META domain-containing protein | 48.4 | 8 (3) |
| LmjF.32.2150 | Hypothetical protein, conserved | 117.6 | 8 (3) |
| LmjF.21.0430 | Hypothetical protein, conserved | 44.7 | 8 (2) |
| LmjF.36.2510 | Nucleoporin NUP96 | 96.9 | 8 (1) |
| LmjF.35.0070 | Prohibitin, putative | 32.3 | 6 (2) |
| LmjF.21.1555 | Hypothetical protein, conserved | 46.4 | 5 (3) |
| LmjF.13.1360 | Hypothetical protein, conserved | 90.9 | 5 (3) |
| LmjF.32.0840 | Hypothetical protein, conserved | 57.4 | 4 (3) |
| LmjF.33.2270 | Hypothetical protein, conserved | 59.6 | 3 (2) |
| LmjF.32.0620 | Hypothetical protein, conserved | 41.8 | 3 (1) |
| LmjF.09.0840 | Uncharacterized protein family (UPF0160), putative | 42.5 | 3 (1) |
| LmjF.33.2800 | Hypothetical protein, conserved | 17 | 2 (2) |
| LmjF.22.0500 | Hypothetical protein, conserved | 54.9 | 2 (2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Florencio-Martínez, L.E.; Cano-Santiago, A.; Mondragón-Rosas, F.; Gómez-García, M.; Flores-Pérez, C.; Román-Carraro, F.C.; Barocio-Rodríguez, L.A.; Manning-Cela, R.G.; Nepomuceno-Mejía, T.; Martínez-Calvillo, S. Participation of TFIIIB Subunit Brf1 in Transcription Regulation in the Human Pathogen Leishmania major. Genes 2021, 12, 280. https://doi.org/10.3390/genes12020280
Florencio-Martínez LE, Cano-Santiago A, Mondragón-Rosas F, Gómez-García M, Flores-Pérez C, Román-Carraro FC, Barocio-Rodríguez LA, Manning-Cela RG, Nepomuceno-Mejía T, Martínez-Calvillo S. Participation of TFIIIB Subunit Brf1 in Transcription Regulation in the Human Pathogen Leishmania major. Genes. 2021; 12(2):280. https://doi.org/10.3390/genes12020280
Chicago/Turabian StyleFlorencio-Martínez, Luis E., Andrés Cano-Santiago, Fabiola Mondragón-Rosas, Maricarmen Gómez-García, Carlos Flores-Pérez, Fiordaliso C. Román-Carraro, Luis A. Barocio-Rodríguez, Rebeca G. Manning-Cela, Tomás Nepomuceno-Mejía, and Santiago Martínez-Calvillo. 2021. "Participation of TFIIIB Subunit Brf1 in Transcription Regulation in the Human Pathogen Leishmania major" Genes 12, no. 2: 280. https://doi.org/10.3390/genes12020280