
Molecular Pathways and Pigments Underlying the Colors of the Pearl Oyster Pinctada margaritifera var. cumingii (Linnaeus 1758)

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. RNA Extraction, Purification, and Sequencing
2.3. Bioinformatics Analysis
2.4. Transcriptome Functional Annotation
2.5. Differential Molecular Function and Gene Expression
2.6. Enzymatic Structure Analysis by Homology Modeling
2.7. Raman Spectroscopy
2.8. Raman Spectra Analysis
3. Results
3.1. Sequencing Results
3.2. Transcriptome-Wide Functional Analysis
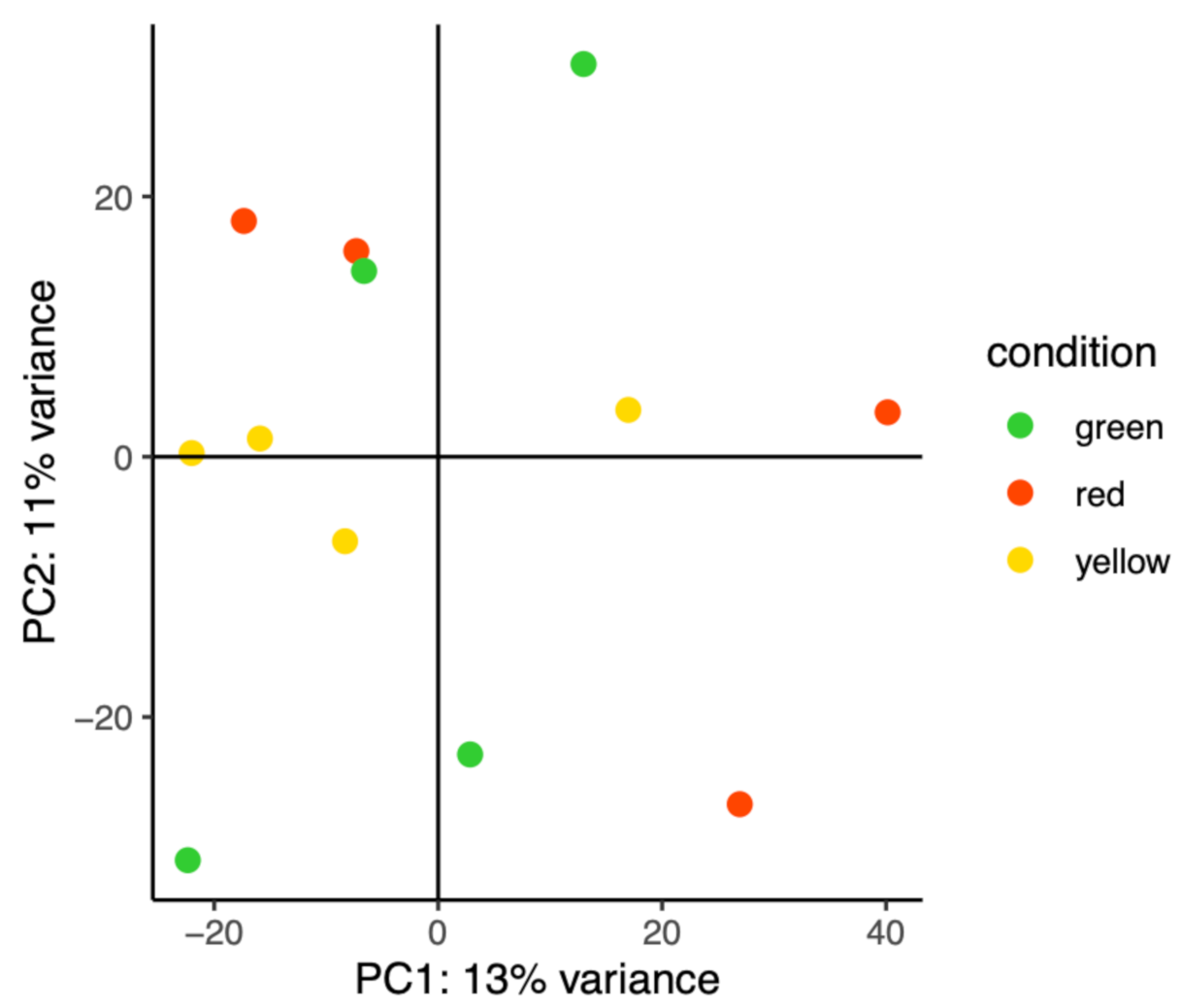
3.3. DEG Functional Analysis
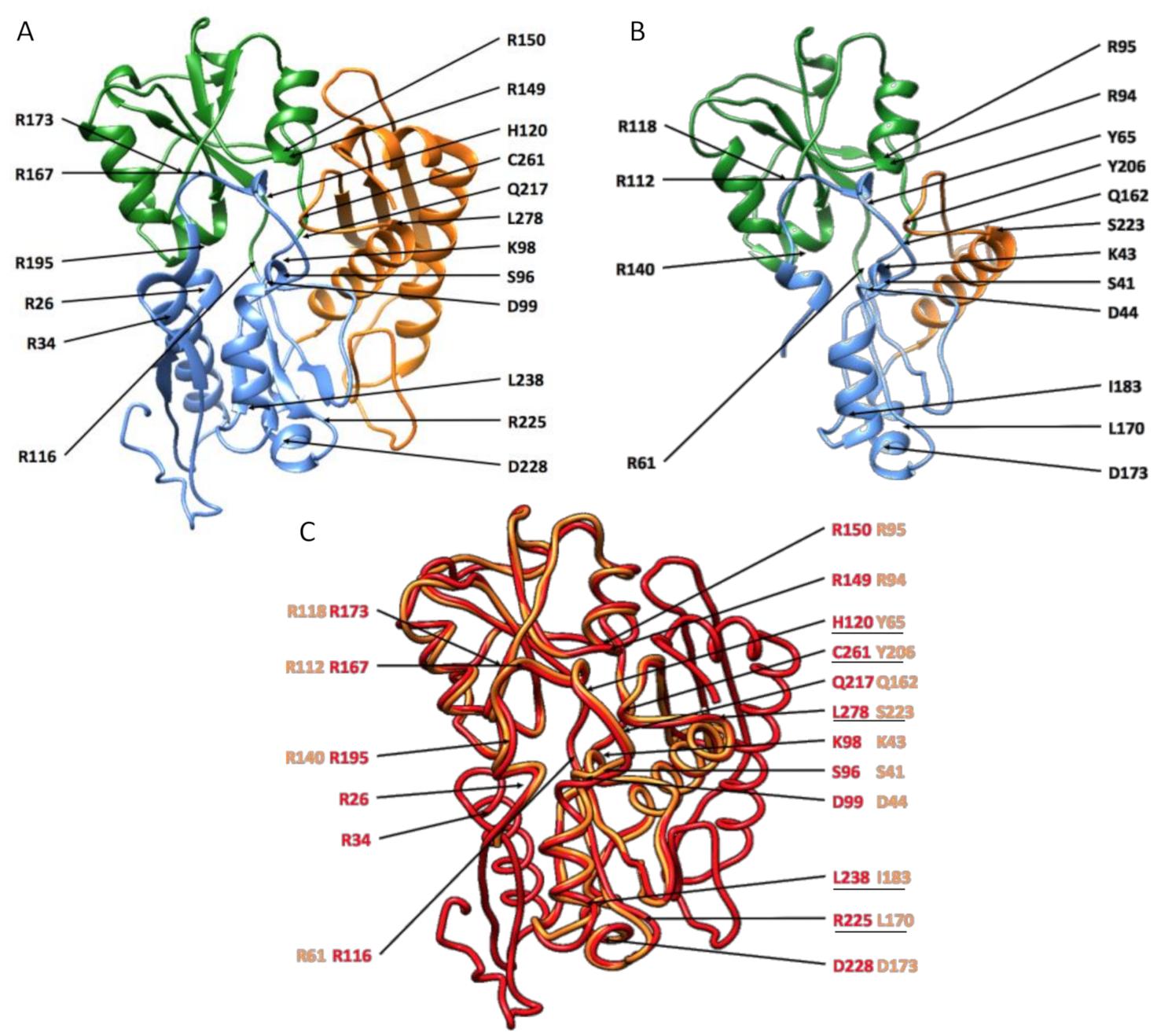
3.4. Enzymatic Structure Analysis with Homology Modeling
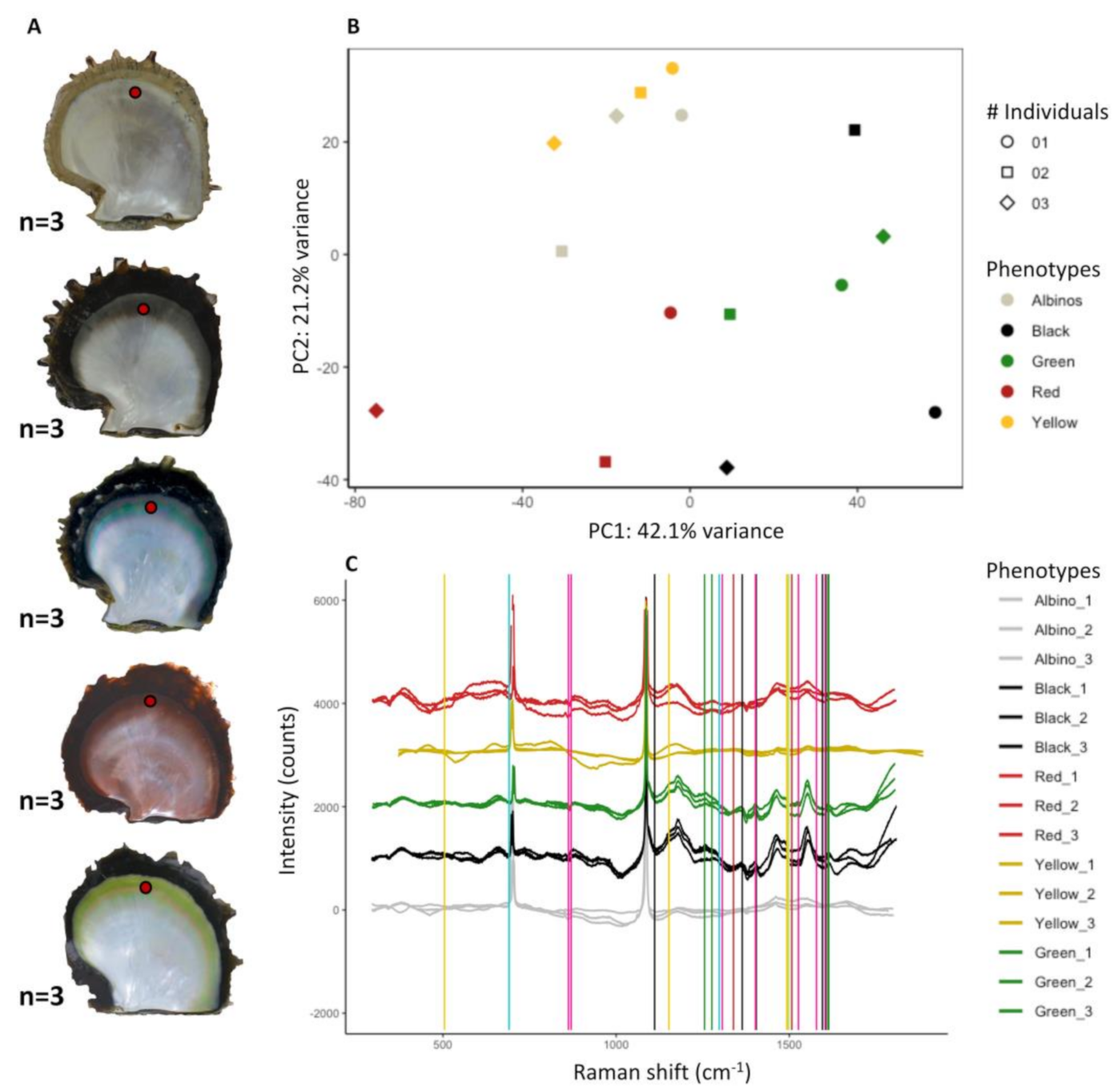
3.5. Raman Spectra Analysis
4. Discussion
4.1. The Inner Shell of P. margaritifera: Chemically Complex, Polychromic, but Containing Dominant Pigments
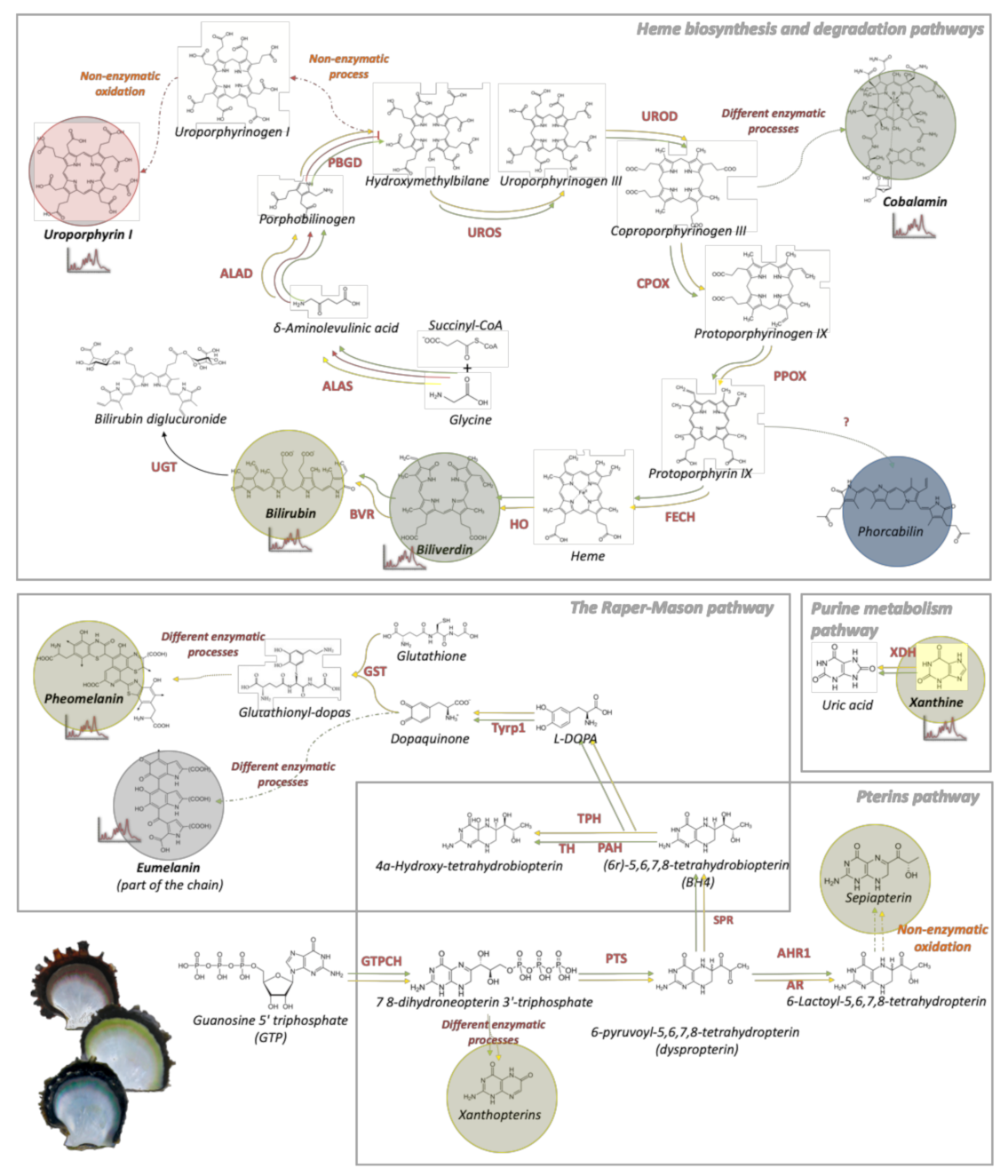
4.2. Dysfunctions of Porphobilinogen Deaminase in the Heme Pathways Produce Red Uroporphyrin and its Derivates in Red Individuals
4.3. Heme, Raper–Mason, and Purine Metabolism Pathways Produce Bilirubin, Pheomelanin, and Xanthine Pigments Underlying the Yellow Phenotype
4.4. Heme Pathways Are Central to the Green Phenotype through the Production of Biliverdin and Green Forms of Cobalamin
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosenberg, G. A New Critical Estimate of Named Species-Level Diversity of the Recent Mollusca. Am. Malacol. Bull. 2014, 32, 308–322. [Google Scholar] [CrossRef]
- Williams, S.T. Molluscan shell colour. Biol. Rev. 2016. [Google Scholar] [CrossRef] [PubMed]
- Cuthill, I.C.; Allen, W.L.; Arbuckle, K.; Caspers, B.; Chaplin, G.; Hauber, M.E.; Hill, G.E.; Jablonski, N.G.; Jiggins, C.D.; Kelber, A.; et al. The biology of color. Science (80-) 2017, 357. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Kawahara, I.; Hasekura, O.; Kijima, A. Genetic control of bluish shell color variation in the Pacific abalone, Haliotis discus hannai. J. Shellfish Res. 2004, 23, 1153–1157. [Google Scholar]
- Liu, X.; Wu, F.; Zhao, H.; Zhang, G.; Guo, X. A Novel Shell Color Variant of the Pacific Abalone Haliotis Discus Hannai Ino Subject to Genetic Control and Dietary Influence. J. Shellfish Res. 2009, 28, 419–424. [Google Scholar] [CrossRef]
- Canales-Gómez, E.; Correa, G.; Viana, M.T. Effect of commercial carotene pigments (astaxanthin, cantaxanthin and β-carotene) in juvenile abalone Haliotis rufescens diets on the color of the shell or nacre. Vet. Mex. 2010, 41, 191–200. [Google Scholar]
- Creese, R.G.; Underwood, A.J. Observations on the biology of the trochid gastropod Austrocochlea constricta (Lamarck) (Prosobranchia). I. Factors affecting shell-banding pattern. J. Exp. Mar. Biol. Ecol. 1976, 23, 211–228. [Google Scholar] [CrossRef]
- Leighton, D.L. Observations on the effect of diet on shell coloration on the red abalone Haliotis rufescens Swainson. Veliger 1961, 4, 29–32. [Google Scholar]
- Fox, D.L. Biochromy, Natural Coloration of Living Things; University of California Press: Berkeley, CA, USA, 1979. [Google Scholar]
- Comfort, A. Acid-soluble pigments of shells; the distribution of porphyrin fluorescence in molluscan shells. Biochem. J. 1949, 44, 111–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comfort, A. Acid-soluble Pigments of Molluscan Shells. Biochem. J. 1947, 45, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Comfort, A. The pigmentation of molluscan shells. Biochem. J. 1950, 45, 208. [Google Scholar] [CrossRef]
- Feng, D.; Li, Q.; Yu, H.; Zhao, X.; Kong, L. Comparative transcriptome analysis of the pacific oyster Crassostrea gigas characterized by shell colors: Identification of genetic bases potentially involved in pigmentation. PLoS ONE 2015, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, X.; Nie, Q.; Xiao, G.; Liu, B. Transcriptome Analysis of Shell Color-Related Genes in the Clam Meretrix meretrix. Mar. Biotechnol. 2015, 17, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Zhao, L.; Chang, Y.; Zhao, W.; Du, Z.; Hao, Z. Transcriptome sequencing and characterization of Japanese scallop Patinopecten yessoensis from different shell color lines. PLoS ONE 2015, 10, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinohara, M.; Kinoshita, S.; Tang, E.; Funabara, D.; Kakinuma, M. Comparison of two pearl sacs formed in the same recipient oyster with different genetic background Involved in yellow pigmentation in Pinctada fucata. Mar. Biotechnol. 2018, 20, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Lemer, S.; Saulnier, D.; Gueguen, Y.; Planes, S. Identification of genes associated with shell color in the black-lipped pearl oyster, Pinctada margaritifera. BMC Genomics 2015, 16, 568. [Google Scholar] [CrossRef] [PubMed]
- Iwahashi, Y.; Akamatsu, S. Porphyrin pigment in Black-Lip pearls and Its application to pearl identification. Fish. Sci. 1994, 60, 69–71. [Google Scholar] [CrossRef]
- Karampelas, S.; Fritsch, E.; Makhlooq, F.; Mohamed, F.; Al-Alawi, A. Raman spectroscopy of natural and cultured pearls and pearl producing mollusc shells. J. Raman Spectrosc. 2019, 1–9. [Google Scholar] [CrossRef]
- Elen, S. Identification of yellow cultured pearls from the Black-lipped oyster Pinctada margaritifera. Gems Gemol. 2002, 38, 66–72. [Google Scholar] [CrossRef]
- Caseiro, J. L’huître Perlière de Polynésie: Biominéralisation, Paramètres et Processus de Croissance, Effets Chromatiques Dans la Coquille et la Perle de Pinctada margaritifera. Ph.D. Thesis, Claude Bernard University Lyon 1, Lyon, France, 1993. [Google Scholar]
- Ky, C.L.; Demmer, J.; Blay, C.; Lo, C. Age-dependence of cultured pearl grade and colour in the black-lipped pearl oyster Pinctada margaritifera. Aquac. Res. 2015, 48, 955–968. [Google Scholar] [CrossRef] [Green Version]
- Ky, C.-L.; Le Pabic, L.; Sham Koua, M.; Nicolas, M.; Seiji, N.; Devaux, D. Is pearl colour produced from Pinctada margaritifera predictable through shell phenotypes and rearing environments selections? Aquaculture 2017, 48, 1041–1057. [Google Scholar] [CrossRef] [Green Version]
- Ky, C.-L.; Nakasai, S.; Pommier, S.; Sham Koua, M.; Devaux, D. The Mendelian inheritance of rare flesh and shell colour variants in the black-lipped pearl oyster (Pinctada margaritifera). Anim. Genet. 2016, 47, 610–614. [Google Scholar] [CrossRef] [Green Version]
- Ky, C.-L.; Blay, C.; Sham-Koua, M.; Vanaa, V.; Lo, C.; Cabral, P. Family effect on cultured pearl quality in black-lipped pearl oyster Pinctada margaritifera and insights for genetic improvement. Aquat. Living Resour. 2013, 26, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Le Moullac, G.; Soyez, C.; Vidal-Dupiol, J.; Belliard, C.; Fievet, J.; Sham-Koua, M.; Lo-Yat, A.; Saulnier, D.; Gaertner-Mazouni, N.; Gueguen, Y. Impact of pCO2 on the energy, reproduction and growth of the shell of the pearl oyster Pinctada margaritifera. Estuar. Coast. Shelf Sci. 2016, 182, 274–282. [Google Scholar] [CrossRef] [Green Version]
- Ky, C.-L.; Nakasai, S.; Molinari, N.; Devaux, D. Influence of grafter skill and season on cultured pearl shape, circles and rejects in Pinctada margaritifera aquaculture in Mangareva lagoon. Aquaculture 2014, 435, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Le Moullac, G.; Soyez, C.; Latchere, O.; Vidal-Dupiol, J.; Fremery, J.; Saulnier, D.; Lo Yat, A.; Belliard, C.; Mazouni-Gaertner, N.; Gueguen, Y. Pinctada margaritifera responses to temperature and pH: Acclimation capabilities and physiological limits. Estuar. Coast. Shelf Sci. 2016, 182, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Genome analysis Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Luyer, J.; Auffret, P.; Quillien, V.; Leclerc, N.; Reisser, C.; Ky, C. Whole transcriptome sequencing and biomineralization gene architecture associated with cultured pearl quality traits in the pearl oyster, Pinctada margaritifera. BMC Genom. 2019, 20, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. Genome analysis HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Nguyen, V.H.; Lavenier, D. PLAST: Parallel local alignment search tool for database comparison. BMC Bioinformatics 2009, 10, 329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Madden, T.L.; Alejandro, S.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Bairoch, A.; Apweiler, R. The SWISS-PROT protein sequence database and its supplement TrEMBL in 2000. Nucleic Acids Res. 2000, 28, 45–48. [Google Scholar] [CrossRef]
- Zdobnov, E.M.; Apweiler, R. InterProScan—An integration platform for the signature-recognition methods in InterPro. Bioinformatics 2001, 17, 847–848. [Google Scholar] [CrossRef] [Green Version]
- Conesa, A.; Götz, S.; García-gómez, J.M.; Terol, J.; Talón, M.; Genómica, D.; Valenciano, I.; Agrarias, D.I.; Valencia, U.P. De Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. BMC Bioinform. 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Wright, R.M.; Aglyamova, G.V.; Meyer, E.; Matz, M. V Gene expression associated with white syndromes in a reef building coral, Acropora hyacinthus. BMC Genom. 2015, 16, 371. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Struct. Biol. 2004, 157, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Song, G.; Li, Y.; Cheng, C.; Zhao, Y.; Gao, A.; Zhang, R.; Joachimiak, A.; Shaw, N.; Liu, Z. Structural insight into acute intermittent porphyria. FASEB J. 2009, 23, 396–404. [Google Scholar] [CrossRef]
- Maiti, R.; Van Domselaar, G.H.; Zhang, H.; Wishart, D.S. SuperPose: A simple server for sophisticated structural superposition. Nucleic Acids Res. 2004, 32, 590–594. [Google Scholar] [CrossRef] [Green Version]
- Bonnier, F.; Byrne, H.J. Understanding the molecular information contained in principal component analysis of vibrational spectra of biological systems. Analyst 2012, 137, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Buzgar, N.; Ionut Apopei, A. The Raman study of certain carbonates. Geologie 2009, 55, 97–112. [Google Scholar]
- Aphalo, P.J. The r4photobiology Suite: Spectral Irradiance; Package ‘ggspectra’; CRAN: Auckland, New-Zealand, 2015. [Google Scholar] [CrossRef]
- Barnard, W.; De Waal, D. Raman investigation of pigmentary molecules in the molluscan biogenic matrix. J. Raman Spectrosc. 2006, 37, 342–352. [Google Scholar] [CrossRef]
- Williams, S.T.; Lockyer, A.E.; Dyal, P.; Nakano, T.; Churchill, C.K.C.; Speiser, D.I. Colorful seashells: Identification of haem pathway genes associated with the synthesis of porphyrin shell color in marine snails. Ecol. Evol. 2017, 7, 10379–10397. [Google Scholar] [CrossRef] [Green Version]
- Pirone, C.; Quirke, J.M.E.; Priestap, H.A.; Lee, D.W. Animal pigment bilirubin discovered in plants. J. Am. Chem. Soc. 2009, 131, 2830. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, G.; Yoshida, T.; Noguchi, M. Heme oxygenase and heme degradation. Biochem. Biophys. Res. Commun. 2005, 338, 558–567. [Google Scholar] [CrossRef]
- Shamim, G.; Ranjan, S.K.; Pandey, D.M.; Ramani, R. Biochemistry and biosynthesis of insect pigments. Eur. J. Entomol. 2014, 111, 149–164. [Google Scholar] [CrossRef] [Green Version]
- Schallreuter, K.U.; Schulz-Douglas, V.; Bünz, A.; Beazley, W.; Körner, C. Pteridines in the control of pigmentation. J. Invest. Dermatol. 1997, 109, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Siracusa, L.D. The agouti gene: Turned on to yellow. Trends Genet. 1994, 10, 423–428. [Google Scholar] [CrossRef]
- Bagnara, J.T.; Matsumoto, J.; Ferris, W.; Frost, S.K.; William, A.; Tchen, T.T.; Taylor, J.D. Common origin of pigment cells. Science (80-) 1979, 203, 410–415. [Google Scholar] [CrossRef]
- Sonthalia, S.; Daulatabad, D.; Sarkar, R. Glutathione as a skin whitening agent: Facts, myths, evidence and controversies. Indian J. Dermatol. 2016, 82, 262. [Google Scholar] [CrossRef]
- Williams, S.T.; Ito, S.; Wakamatsu, K.; Goral, T.; Edwards, N.P.; Wogelius, R.A. Identification of shell colour pigments in marine snails Clanculus pharaonius and C. margaritarius (Trochoidea; Gastropoda). PLoS ONE 2016, 11, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Pearson, W.R. An introduction to sequence similarity (“homology”) searching. Curr. Protoc. Bioinform. 2013, 42, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Furuichi, T.; Yoshikawa, S.; Miyawakft, A.; Wadat, K.; Maedat, N.; Mikoshiba, K. Primary structure and functional expression of the inositol 1, 4, 5-trisphosphate-binding protein P 400. Nature 1989, 342, 714–716. [Google Scholar] [CrossRef] [PubMed]
- Wek, R.C.; Cannon, J.F.; Dever, T.E.; Hinnebuschl, A.G. Truncated protein phosphatase GLC7 restores translational activation of GCN4 expression in yeast mutants defective for the eIF-2a kinase GCN2. Mol. Cell. Biol. 1992, 12, 5700–5710. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.H.E.; Marassi, F.M.; Jones, D.H.; Straus, S.K.; Bour, S.; Strebel, K.; Schubert, U.; Oblatt-montal, M.; Montal, M.; Opella, S.J. Expression, purification, and activities of full-length and truncated versions of the integral membrane protein Vpu from HIV-1. Protein Sci. 2002, 11, 546–557. [Google Scholar] [CrossRef] [Green Version]
- Farre, B.; Dauphin, Y. Lipids from the nacreous and prismatic layers of two Pteriomorpha Mollusc shells. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 152, 103–109. [Google Scholar] [CrossRef]
- Marie, B.; Joubert, C.; Tayalé, A.; Zanella-cléon, I. Different secretory repertoires control the biomineralization processes of prism and nacre deposition of the pearl oyster shell. Proc. Natl. Acad. Sci. USA 2013, 109, 20986–20991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, F.; Li, G.; Haws, M.; Niu, T. Element concentrations in shell of Pinctada margaritifera from French Polynesia and evaluation for using as a food supplement. Food Chem. 2007, 104, 1171–1176. [Google Scholar] [CrossRef]
- Wang, H.; Osseiran, S.; Igras, V.; Nichols, A.J.; Roider, E.M.; Pruessner, J.; Tsao, H.; Fisher, D.E.; Evans, C.L. In vivo coherent Raman imaging of the melanomagenesis-associated pigment pheomelanin. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ky, C.L.; Lo, C.; Planes, S. Mono- and polychromatic inner shell phenotype diversity in Pinctada margaritifera donor pearl oysters and its relation with cultured pearl colour. Aquaculture 2017, 468, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Stenger, P.-L.; Vidal-Dupiol, J.; Reisser, C.; Planes, S.; Ky, C. Colour plasticity in the shells and pearls of animal graft model Pinctada margaritifera through colour quantification with the HSV system. Sci. Rep. 2019, 75, 20. [Google Scholar] [CrossRef]
- Ky, C.L.; Quillien, V.; Broustal, F.; Soyez, C.; Devaux, D. Phenome of pearl quality traits in the mollusc transplant model Pinctada margaritifera. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Paoli, M.; Marles-wright, J.O.N.; Smith, A.N.N. Structure—Function relationships in heme-proteins. DNA Cell Biol. 2002, 21. [Google Scholar] [CrossRef] [Green Version]
- Jordan, P.M.; Warren, M.J. Evidence for a dipyrromethane cofactor at the catalytic of E. coli porphobilinogen deaminase. FEBS Lett. 1987, 225, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.K.; Peters, T.J. Urine and faecal porphyrin profiles by reversed-phase high-performance liquid chromatography in the porphyrias. Clin. Chim. Acta 1984, 139, 55–63. [Google Scholar] [CrossRef]
- Doss, M.O. Dual porphyria in double heterozygotes with porphobilinogen deaminase and uroporphyrinogen decarboxylase deficiencies. Clin. Genet. 1989, 35, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Shoolingin-Jordan, P.M. Porphobilinogen deaminase and uroporphyrinogen III synthase: Structure, molecular biology, and mechanism. J. Bioenerg. Biomembr. 1995, 27, 181–195. [Google Scholar] [CrossRef]
- Kainrath, S.; Stadler, M.; Reichhart, E.; Distel, M.; Janovjak, H. Green-light-induced inactivation of receptor signaling using cobalamin-binding domains. Angew. Chem. Int. Ed. 2017, 56, 4608–4611. [Google Scholar] [CrossRef]
- Ajioka, R.S.; Phillips, J.D.; Kushner, J.P. Biosynthesis of heme in mammals. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2006, 1763, 723–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravera, O.; Cenci, R.; Beone, G.M.; Dantas, M.; Lodigiani, P. Trace element concentrations in freshwater mussels and macrophytes as related to those in their environment. J. Limnol. 2003, 62, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Meyer, U.A.; Strand, L.J.; Doss, M.; Rees, A.C.; Marver, H.S. Intermittent acute porphyria—demonstration of a genetic defect in porphobilinogen metabolism. N. Engl. J. Med. 1972, 286, 1277–1282. [Google Scholar] [CrossRef]
- Fox, D.L. Animal Biochromes and Structural Colours: Physical, Chemical, Distributional & Physiological Features of Coloured Bodies in the Animal World; University of California: Oakland, CA, USA, 1976. [Google Scholar]
- Tenhunen, R.; Marver, H.S.; Schmid, R. The enzymatic conversion of heme to bilirubin by microsomal heme oxygenase. Proc. Natl. Acad. Sci. 1968, 61, 748–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singleton, J.W.; Laster, L. Biliverdin reductase of guinea pig liver. J. Biol. Chem. 1965, 240, 4780–4789. [Google Scholar] [CrossRef]
- Bouétard, A.; Noirot, C.; Besnard, A.L.; Bouchez, O.; Choisne, D.; Robe, E.; Klopp, C.; Lagadic, L.; Coutellec, M.A. Pyrosequencing-based transcriptomic resources in the pond snail Lymnaea stagnalis, with a focus on genes involved in molecular response to diquat-induced stress. Ecotoxicology 2012, 21, 2222–2234. [Google Scholar] [CrossRef]
- Jansen, P. The enzyme-catalyzed formation of bilirubin diglucuronide by a solubilized preparation from cat liver microsomes. Biochim. Biophys. Acta (BBA) General Subj. 1974, 338, 170–182. [Google Scholar] [CrossRef]
- Nawa, S. The Structure of the Yellow Pigment from Drosophila. Bull. Chem. Soc. Jpn. 1960, 33, 1555–1560. [Google Scholar] [CrossRef]
- Blay, C.; Planes, S.; Ky, C.L. Donor and recipient contribution to phenotypic traits and the expression of biomineralisation genes in the pearl oyster model Pinctada margaritifera. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- McGraw, K.J. Mechanics of uncommon colors: Pterins, porphyrins, and psittacofulvins. In Bird Coloration; Harvard University Press: Cambridge, MA, USA, 2006; pp. 354–398. [Google Scholar]
- Grant, H.E.; Williams, S.T. Phylogenetic distribution of shell colour in Bivalvia (Mollusca). Biol. J. Linn. Soc. 2018, 125, 377–391. [Google Scholar] [CrossRef]
- Mills, M.G.; Patterson, L.B. Not just black and white: Pigment pattern development and evolution in vertebrates. Semin. Cell Dev. Biol. 2009, 20, 72–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrier, C.; Dupuy, C.; Jardon, P.; Gautron, R. The configurations in solution of the biliverdin TXγ phorcabilin and isophorcabilin dimethyl esters. Tetrahedron Lett. 1981, 22, 855–858. [Google Scholar] [CrossRef]
- Choussy, M.; Barbier, M. Photo-oxidation and photoprotection in the IXγ bile pigment series: A comparison of the photoprotective roles of pterobilin, phorcabilin and sarpedobilin in vitro. Tetrahedron 1983, 39, 1915–1918. [Google Scholar] [CrossRef]
- Ju, Z.Y.; Viljoen, C.; Hutchinson, P.; Reinicke, J.; Horgen, F.D.; Howard, L.; Lee, C. Effects of diets on the growth performance and shell pigmentation of Pacific abalone. Aquac. Res. 2016, 47, 4004–4014. [Google Scholar] [CrossRef]
- Sassa, S. Why heme needs to be degraded to iron, biliverdin IX, and carbon monoxide? Antioxid. Redox Signal. 2004, 6, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Kikkawa, H.; Fujito, S. Nature of pigments derived from tyrosine and tryptophan in Animals. Science (80-) 1955, 121, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Kumudha, A.; Sarada, R. Effect of different extraction methods on vitamin B12 from blue green algae, Spirulina platensis. Pharm. Anal. Acta 2015, 6, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Roper, J.M.; Raux, E.; Brindley, A.A.; Schubert, H.L.; Gharbia, S.E.; Shah, H.N.; Warren, M.J. The enigma of cobalamin (vitamin B12) biosynthesis in Porphyromonas gingivalis: Identification and characterization of a functional corrin pathway. J. Biol. Chem. 2000, 275, 40316–40323. [Google Scholar] [CrossRef] [Green Version]
- Kondo, H.; Kolhouse, J.F.; Allen, R.H. Presence of cobalamin analogues in animal tissues. Proc. Natl. Acad. Sci. USA 1980, 77, 817–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, J.; Lawrence, J.; Bobik, T. Cobalamin (Coenzyme B12): Synthesis and Biological Significance. Ann. Rev. Microbiol. 1996, 50, 137–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croft, M.T.; Lawrence, A.D.; Raux-Deery, E.; Warren, M.J.; Smith, A.G. Algae acquire vitamin B12 through a symbiotic relationship with bacteria. Nature 2005, 438, 90–93. [Google Scholar] [CrossRef]
- Xie, B.; Bishop, S.; Stessman, D.; Wright, D.; Spalding, M.H.; Halverson, L.J. Chlamydomonas reinhardtii thermal tolerance enhancement mediated by a mutualistic interaction with vitamin B12-producing bacteria. ISME J. 2013, 7, 1544–1555. [Google Scholar] [CrossRef]
- Ruaux, C.G. Cobalamin in companion animals: Diagnostic marker, deficiency states and therapeutic implications. Vet. J. 2013, 196, 145–152. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Milani, C.; de Giori, G.S.; Sesma, F.; van Sinderen, D.; Ventura, M. Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef]
- Pathare, P.M.; Wilbur, D.S.; Heusser, S.; Quadros, E.V.; Mcloughlin, P.; Morgan, A.C. Synthesis of Cobalamin—Biotin Conjugates That Vary in the Position of Cobalamin Coupling. Evaluation of Cobalamin Derivative Binding to Transcobalamin II. Bioconjug. Chem. 1996, 7, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, B.; Pong-Wong, R.; Woolliams, J.A. Marker assisted selection with optimised contributions of the candidates to selection. Genet. Sel. Evol. 2002, 34, 679–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekkers, J.C.M. Marker-assisted selection for commercial crossbred performance. J. Anim. Sci. 2007, 85, 2104–2114. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Human’s_PBGD | PBGD_A | PBGD_B | PBGD_C | PBGD_D |
|---|---|---|---|---|
| R26 | NA | NA | H23 | NA |
| Q34 | NA | NA | L32 | NA |
| N88 | K33 | NA | K82 | NA |
| S96 | S41 | NA | S90 | NA |
| K98 | K43 | NA | K92 | NA |
| D99 | D44 | NA | D93 | NA |
| R116 | R61 | NA | R110 | NA |
| H120 | Y65 | NA | Y114 | NA |
| R149 | R94 | NA | R143 | NA |
| R150 | R95 | NA | R144 | NA |
| R167 | R112 | NA | V161 | R14 |
| R173 | R118 | NA | NA | R20 |
| R195 | R140 | NA | NA | R42 |
| Q217 | Q162 | NA | NA | Q64 |
| R225 | L170 | NA | NA | R72 |
| D228 | D173 | NA | NA | D75 |
| L238 | I183 | I8 | NA | Y85 |
| C261 | Y206 | Y31 | NA | Y108 |
| L278 | S223 | L48 | NA | NA |
| Pigment | Peaks | Publication | Red individual 1 | Red individual 2 | Red individual 3 | Yellow individual 1 | Yellow individual 2 | Yellow individual 3 | Green individual 1 | Green individual 2 | Green individual 3 | Black individual 1 | Black individual 2 | Black individual 3 | Albino individual 1 | Albino individual 2 | Albino individual 3 | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Peak | Intensity | Ratio | Peak | Intensity | Ratio | Peak | Intensity | Ratio | Peak | Intensity | Ratio | Peak | Intensity | Ratio | Peak | Intensity | Ratio | Peak | Intensity | Ratio | Peak | Intensity | Ratio | Peak | Intensity | Ratio | Peak | Intensity | Ratio | Peak | Intensity | Ratio | Peak | Intensity | Ratio | Peak | Intensity | Ratio | Peak | Intensity | Ratio | Peak | Intensity | Ratio | |||
| Uropor--phyrin I | 1335 | Williams et al. 2017 | yes | 29.26 | 0.66 | yes | 19.70 | 0.49 | yes | 16.05 | 0.50 | ||||||||||||||||||||||||||||||||||||
| 1445 | yes | 44.39 | 0.83 | yes | 29.10 | 0.72 | yes | 120.49 | 0.77 | yes | 46.51 | 0.64 | |||||||||||||||||||||||||||||||||||
| 1556 | yes | 13.18 | 0.57 | ||||||||||||||||||||||||||||||||||||||||||||
| Copperuroporphyrin | 1310 | Shelnutt 1981, 1982, et al. 1984 | yes | 6.52 | 0.66 | yes | 5.63 | 0.46 | yes | 5.37 | 0.22 | ||||||||||||||||||||||||||||||||||||
| 1379 | yes | 35.73 | 0.25 | ||||||||||||||||||||||||||||||||||||||||||||
| 1403 | yes | 11.34 | 0.27 | yes | 106.76 | 0.51 | yes | 99.57 | 0.41 | yes | 62.13 | 0.43 | |||||||||||||||||||||||||||||||||||
| 1499 | yes | 27.21 | 0.73 | yes | 19.07 | 0.74 | yes | 8.45 | 0.31 | ||||||||||||||||||||||||||||||||||||||
| 1582 | yes | 17.18 | 0.71 | ||||||||||||||||||||||||||||||||||||||||||||
| 1637 | yes | 10.28 | 0.66 | ||||||||||||||||||||||||||||||||||||||||||||
| FeIIIuroporphyrin | 1307 | Shelnutt 1982 | yes | 5.48 | 0.66 | yes | 11.81 | 0.45 | |||||||||||||||||||||||||||||||||||||||
| 1376 | yes | 9.29 | 0.62 | yes | 29.43 | 0.51 | yes | 21.71 | 0.23 | yes | 59.22 | 0.49 | |||||||||||||||||||||||||||||||||||
| 1402 | yes | 11.29 | 0.67 | yes | 16.37 | 0.54 | yes | 11.34 | 0.26 | yes | 114.98 | 0.51 | |||||||||||||||||||||||||||||||||||
| 1489 | yes | 12.80 | 0.76 | yes | 16.93 | 0.74 | yes | 8.04 | 0.31 | yes | 111.01 | 0.73 | yes | 42.97 | 0.51 | ||||||||||||||||||||||||||||||||
| 1581 | yes | 15.64 | 0.70 | yes | 4.23 | 0.30 | |||||||||||||||||||||||||||||||||||||||||
| 1627 | yes | 16.99 | 0.72 | ||||||||||||||||||||||||||||||||||||||||||||
| Biliverdin | 1252 | Margulies and Toporowicz 1984, Holt et al. 1989 | yes | 40.58 | 0.81 | ||||||||||||||||||||||||||||||||||||||||||
| 1273 | yes | 24.18 | 0.56 | ||||||||||||||||||||||||||||||||||||||||||||
| 1606 | yes | 11.07 | 0.73 | yes | 20.05 | 0.56 | yes | 12.97 | 0.60 | ||||||||||||||||||||||||||||||||||||||
| 1608 | yes | 38.36 | 0.73 | yes | 7.60 | 0.56 | yes | 21.08 | 0.60 | ||||||||||||||||||||||||||||||||||||||
| Cobala--min | 1495 | Puckett et al 1996, Galluzzi et al. 1974 | yes | 16.70 | 0.72 | yes | 14.17 | 0.57 | |||||||||||||||||||||||||||||||||||||||
| 1502 | yes | 19.50 | 0.75 | yes | 18.87 | 0.72 | yes | 15.99 | 0.56 | yes | 24.57 | 0.53 | |||||||||||||||||||||||||||||||||||
| Xanthine | 862 | Muniz-Miranda et al. 2018 | yes | 25.06 | 0.48 | yes | 29.51 | 0.71 | yes | 17.56 | 0.60 | ||||||||||||||||||||||||||||||||||||
| 870 | yes | 79.86 | 0.71 | yes | 35.32 | 0.57 | yes | 81.36 | 0.21 | yes | 30.95 | 0.72 | yes | 38.01 | 0.53 | yes | 111.64 | 0.66 | yes | 45.10 | 0.51 | ||||||||||||||||||||||||||
| 1303 | yes | 7.64 | 0.54 | yes | 9.09 | 0.53 | |||||||||||||||||||||||||||||||||||||||||
| 1398 | yes | 5.58 | 0.68 | yes | 2.73 | 0.57 | yes | 7.76 | 0.55 | ||||||||||||||||||||||||||||||||||||||
| 1521 | yes | 25.67 | 0.75 | yes | 8.88 | 0.48 | yes | 41.97 | 0.51 | ||||||||||||||||||||||||||||||||||||||
| 1573 | yes | 28.38 | 0.56 | yes | 6.44 | 0.55 | yes | 42.71 | 0.60 | ||||||||||||||||||||||||||||||||||||||
| 1593 | |||||||||||||||||||||||||||||||||||||||||||||||
| 1598 | yes | 9.26 | 0.53 | ||||||||||||||||||||||||||||||||||||||||||||
| Pheomelanin | 507 | Galvan et al. 2013 | yes | 3.67 | 0.54 | yes | 26.49 | 0.48 | |||||||||||||||||||||||||||||||||||||||
| 1150 | yes | 17.71 | 0.56 | yes | 42.43 | 1.00 | |||||||||||||||||||||||||||||||||||||||||
| 1488 | yes | 2.08 | 0.53 | yes | 2.69 | 0.55 | yes | 0.43 | 0.48 | ||||||||||||||||||||||||||||||||||||||
| 1490 | yes | 1.11 | 0.53 | yes | 1.92 | 0.55 | yes | 1.48 | 0.47 | yes | 92.35 | 0.72 | yes | 70.77 | 0.63 | ||||||||||||||||||||||||||||||||
| Bilirubin | 692 | Dybas et al. 2018 | yes | 22.07 | 0.55 | yes | 13.70 | 0.56 | |||||||||||||||||||||||||||||||||||||||
| 1294 | yes | 5.46 | 0.53 | yes | 4.40 | 0.51 | yes | 32.05 | 0.70 | yes | 23.52 | 0.66 | |||||||||||||||||||||||||||||||||||
| Melanin | 1109 | Centeno and Shamir 2008, Williams et al. 2016 | yes | 27.08 | 0.71 | yes | 34.65 | 0.24 | yes | 20.82 | 0.79 | ||||||||||||||||||||||||||||||||||||
| 1360 | yes | 12.87 | 0.49 | yes | 12.00 | 0.67 | |||||||||||||||||||||||||||||||||||||||||
| 1400 | yes | 71.58 | 0.67 | yes | 54.95 | 0.57 | yes | 27.01 | 0.54 | yes | 39.33 | 0.43 | yes | 44.05 | 0.46 | ||||||||||||||||||||||||||||||||
| 1590 | yes | 13.25 | 0.52 | ||||||||||||||||||||||||||||||||||||||||||||
| 1600 | yes | 23.14 | 0.53 | ||||||||||||||||||||||||||||||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stenger, P.-L.; Ky, C.-L.; Reisser, C.; Duboisset, J.; Dicko, H.; Durand, P.; Quintric, L.; Planes, S.; Vidal-Dupiol, J. Molecular Pathways and Pigments Underlying the Colors of the Pearl Oyster Pinctada margaritifera var. cumingii (Linnaeus 1758). Genes 2021, 12, 421. https://doi.org/10.3390/genes12030421
Stenger P-L, Ky C-L, Reisser C, Duboisset J, Dicko H, Durand P, Quintric L, Planes S, Vidal-Dupiol J. Molecular Pathways and Pigments Underlying the Colors of the Pearl Oyster Pinctada margaritifera var. cumingii (Linnaeus 1758). Genes. 2021; 12(3):421. https://doi.org/10.3390/genes12030421
Chicago/Turabian StyleStenger, Pierre-Louis, Chin-Long Ky, Céline Reisser, Julien Duboisset, Hamadou Dicko, Patrick Durand, Laure Quintric, Serge Planes, and Jeremie Vidal-Dupiol. 2021. "Molecular Pathways and Pigments Underlying the Colors of the Pearl Oyster Pinctada margaritifera var. cumingii (Linnaeus 1758)" Genes 12, no. 3: 421. https://doi.org/10.3390/genes12030421
APA StyleStenger, P. -L., Ky, C. -L., Reisser, C., Duboisset, J., Dicko, H., Durand, P., Quintric, L., Planes, S., & Vidal-Dupiol, J. (2021). Molecular Pathways and Pigments Underlying the Colors of the Pearl Oyster Pinctada margaritifera var. cumingii (Linnaeus 1758). Genes, 12(3), 421. https://doi.org/10.3390/genes12030421