
Odorant and Taste Receptors in Sperm Chemotaxis and Cryopreservation: Roles and Implications in Sperm Capacitation, Motility and Fertility


Abstract
:1. Introduction
2. Sperm Chemotaxis and Sperm–Egg Interaction
3. Odorant and Taste Receptors
3.1. Distribution of Odorant and Taste Receptors
3.2. Reproductive Odorant and Taste Receptors
4. Olfactory Transduction and Sperm Signaling Pathways
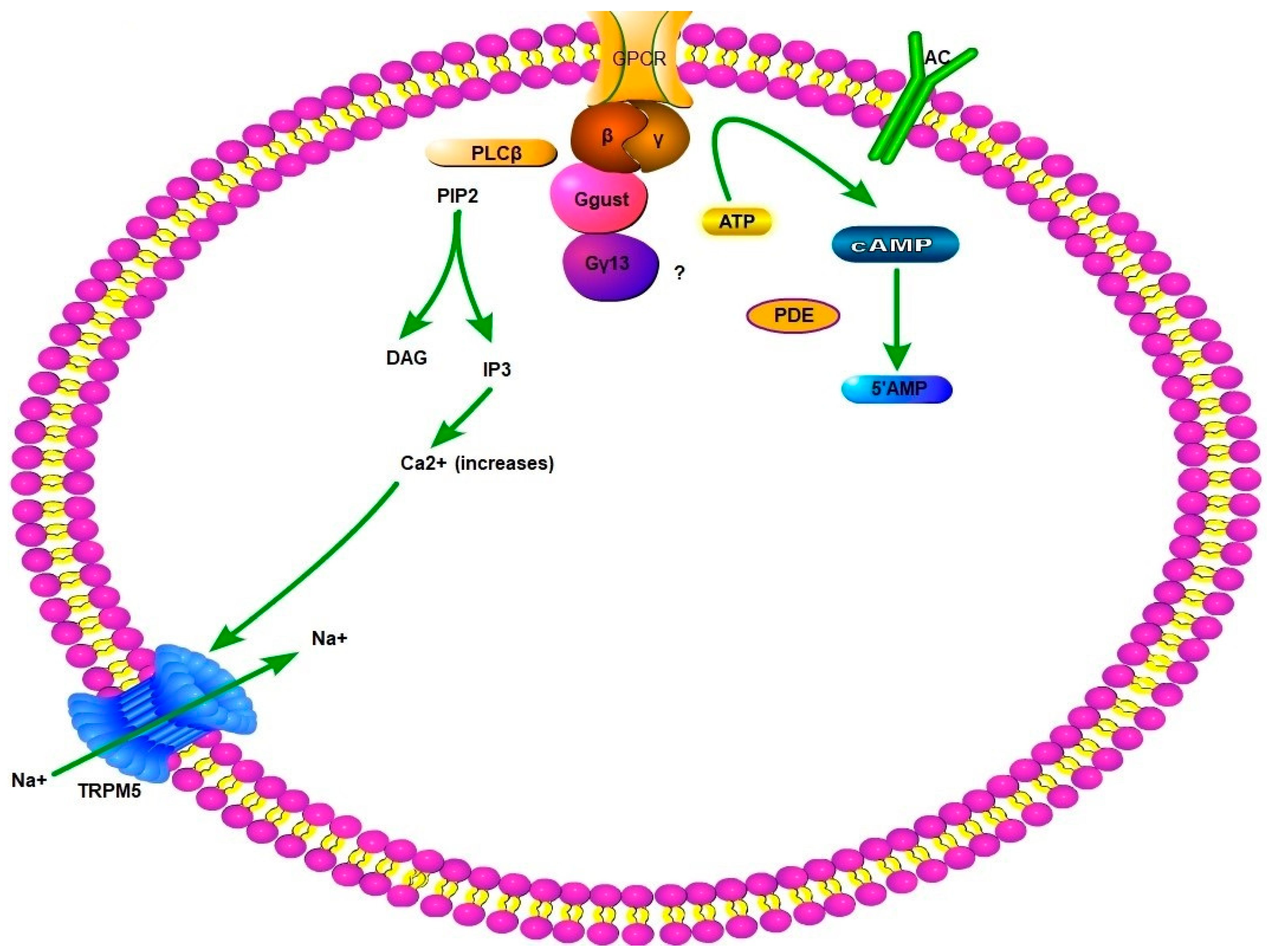
5. Gustatory Transduction
6. Cryopreservation and Olfactory Transduction
7. Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ihara, S.; Yoshikawa, K.; Touhara, K. Chemosensory signals and their receptors in the olfactory neural system. Neuroscience 2013, 254, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Dalton, R.P.; Lomvardas, S.J.A. Chemosensory receptor specificity and regulation. Annu. Rev. Neurosci. 2015, 38, 331–349. [Google Scholar] [CrossRef] [Green Version]
- Ralt, D.; Manor, M.; Cohen-Dayag, A.; Tur-Kaspa, I.; Ben-Shlomo, I.; Makler, A.; Yuli, I.; Dor, J.; Blumberg, S.; Mashiach, S.; et al. Chemotaxis and Chemokinesis of Human Spermatozoa to Follicular Factors1. Biol. Reprod. 1994, 50, 774–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaupp, U.B.; Kashikar, N.D.; Weyand, I. Mechanisms of Sperm Chemotaxis. Annu. Rev. Physiol. 2008, 70, 93–117. [Google Scholar] [CrossRef] [PubMed]
- Parmentier, M.; Libert, F.; Schurmans, S.; Schiffmann, S.; Lefort, A.; Eggerickx, D.; Ledent, C.; Mollereau, C.; Gérard, C.; Perret, J.; et al. Expression of members of the putative olfactory receptor gene family in mammalian germ cells. Nat. Cell Biol. 1992, 355, 453–455. [Google Scholar] [CrossRef] [PubMed]
- Spehr, M.; Gisselmann, G.; Poplawski, A.; Riffell, J.A.; Wetzel, C.H.; Zimmer, R.K.; Hatt, H. Identification of a Testicular Odorant Receptor Mediating Human Sperm Chemotaxis. Science 2003, 299, 2054–2058. [Google Scholar] [CrossRef]
- Braun, T.; Voland, P.; Kunz, L.; Prinz, C.; Gratzl, M. Enterochromaffin Cells of the Human Gut: Sensors for Spices and Odorants. Gastroenterology 2007, 132, 1890–1901. [Google Scholar] [CrossRef]
- Neuhaus, E.M.; Zhang, W.; Gelis, L.; Deng, Y.; Noldus, J.; Hatt, H. Activation of an Olfactory Receptor Inhibits Proliferation of Prostate Cancer Cells. J. Biol. Chem. 2009, 284, 16218–16225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flegel, C.; Manteniotis, S.; Osthold, S.; Hatt, H.; Gisselmann, G. Expression Profile of Ectopic Olfactory Receptors Determined by Deep Sequencing. PLoS ONE 2013, 8, e55368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milardi, D.; Colussi, C.; Grande, G.; Vincenzoni, F.; Pierconti, F.; Mancini, F.; Baroni, S.; Castagnola, M.; Marana, R.; Pontecorvi, A. Olfactory Receptors in Semen and in the Male Tract: From Proteome to Proteins. Front. Endocrinol. 2018, 8, 379. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, N.; Yomogida, K.; Okabe, M.; Touhara, K. Functional characterization of a mouse testicular olfactory receptor and its role in chemosensing and in regulation of sperm motility. J. Cell Sci. 2004, 117, 5835–5845. [Google Scholar] [CrossRef] [Green Version]
- Vanderhaeghen, P.; Schurmans, S.; Vassart, G.; Parmentier, M. Specific Repertoire of Olfactory Receptor Genes in the Male Germ Cells of Several Mammalian Species. Genomics 1997, 39, 239–246. [Google Scholar] [CrossRef]
- Walensky, L.D.; Roskams, A.J.; Lefkowitz, R.J.; Snyder, S.H.; Ronnett, G.V. Odorant Receptors and Desensitization Proteins Colocalize in Mammalian Sperm. Mol. Med. 1995, 1, 130–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Governini, L.; Semplici, B.; Pavone, V.; Crifasi, L.; Marrocco, C.; de Leo, V.; Arlt, E.; Gudermann, T.; Boekhoff, I.; Luddi, A.; et al. Expression of Taste Receptor 2 Subtypes in Human Testis and Sperm. J. Clin. Med. 2020, 9, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Cao, J.; Iguchi, N.; Riethmacher, D.; Huang, L. Functional characterization of bitter-taste receptors expressed in mammalian testis. Mol. Hum. Reprod. 2012, 19, 17–28. [Google Scholar] [CrossRef]
- Spinaci, M.; Bucci, D.; Mazzoni, M.; Giaretta, E.; Bernardini, C.; Vallorani, C.; Tamanini, C.; Clavenzani, P.; Galeati, G.J.T. Expression of α-gustducin and α-transducin, G proteins coupled with taste receptors, in boar sperm. Theriogenology 2014, 82, 144–151. [Google Scholar] [CrossRef]
- Pérez-Cerezales, S.; Laguna-Barraza, R.; de Castro, A.C.; Sánchez-Calabuig, M.J.; Cano-Oliva, E.; de Castro-Pita, F.J.; Montoro-Buils, L.; Pericuesta, E.; Fernández-González, R.; Gutiérrez-Adán, A. Sperm selection by thermotaxis improves ICSI outcome in mice. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.-G.; Wehry, U.P.; Buhren, B.A.; Schrumpf, H.; Oláh, P.; Bünemann, E.; Yu, C.-F.; Chen, S.-J.; Müller, A.; Hirchenhain, J.; et al. CCL20-CCR6 axis directs sperm–oocyte interaction and its dysregulation correlates/associates with male infertility. Biol. Reprod. 2020, 103, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Olaniyan, O.T.; Dare, A.; Okotie, G.E.; Adetunji, C.O.; Ibitoye, B.O.; Eweoya, O.; Dare, J.B.; Okoli, B.J. Ovarian odorant-like biomolecules in promoting chemotaxis behavior of spermatozoa olfactory receptors during migration, maturation, and fertilization. Middle East. Fertil. Soc. J. 2021, 26, 1–14. [Google Scholar] [CrossRef]
- Eisenbach, M.; Giojalas, L.C. Sperm guidance in mammals—An unpaved road to the egg. Nat. Rev. Mol. Cell Biol. 2006, 7, 276–285. [Google Scholar] [CrossRef]
- Kaupp, U. 100 years of sperm chemotaxis. J. Gen. Physiol. 2012, 140, 583–586. [Google Scholar] [CrossRef] [Green Version]
- Ran, M.-X.; Zhou, L.K.; Wang, Z.; Yang, J.-D.; Wu, K.; Huang, Y.; Luo, B.; Qazi, I.H. Comparative Analysis of MicroRNA and mRNA Profiles of Sperm with Different Freeze Tolerance Capacities in Boar (Sus scrofa) and Giant Panda (Ailuropoda melanoleuca). Biomolecules 2019, 9, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago-Moreno, J.; Esteso, M.C.; Pradieé, J.; Castano, C.; Toledano-Díaz, A.; O’Brien, E.; López-Sebastián, A.; Martinez-Nevado, E.; Delclaux, M.; Fernandez-Morán, J.; et al. Giant panda (Ailuropoda melanoleuca) sperm morphometry and function after repeated freezing and thawing. Andrologia 2015, 48, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.G.; Tomasi, L.; Rovasio, R.A.; Giojalas, L.C. Increased velocity and induction of chemotactic response in mouse spermatozoa by follicular and oviductal fluids. Reproduction 1999, 115, 23–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giojalas, L.; Fabro, G.; Eisenbach, M.; Rovasio, R.J.J.A. Capacitated and chemotactic rabbit spermatozoa appear to be shortly available around ovulation. J. Androl. 2001, 22, 92. [Google Scholar]
- Bahat, A.; Tur-Kaspa, I.; Gakamsky, A.; Giojalas, L.C.; Breitbart, H.; Eisenbach, M. Thermotaxis of mammalian sperm cells: A potential navigation mechanism in the female genital tract. Nat. Med. 2003, 9, 149–150. [Google Scholar] [CrossRef]
- Miki, K.; Clapham, D.E. Rheotaxis Guides Mammalian Sperm. Curr. Biol. 2013, 23, 443–452. [Google Scholar] [CrossRef] [Green Version]
- Mondal, A.; Takagi, Y.; Baba, S.A.; Hamano, K.-I. Involvement of calcium channels and intracellular calcium in bull sperm thermotaxis. J. Reprod. Dev. 2017, 63, 143–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.W.; Ki, M.S.; Kim, C.-L.; Hwang, I.-S.; Jeon, I.S. A Simple Confocal Microscopy-based Method for Assessing Sperm Movement. Dev. Reprod. 2017, 21, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Nagata, M.B.; Egashira, J.; Katafuchi, N.; Endo, K.; Ogata, K.; Yamanaka, K.; Yamanouchi, T.; Matsuda, H.; Hashiyada, Y.; Yamashita, K. Bovine sperm selection procedure prior to cryopreservation for improvement of post-thawed semen quality and fertility. J. Anim. Sci. Biotechnol. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.; Hill, C.; Scudamore, I.; Dunphy, B.; Cooke, I.; Barratt, C. Physiology: Sperm numbers and distribution within the human Fallopian tube around ovulation. Hum. Reprod. 1993, 8, 2019–2026. [Google Scholar] [CrossRef]
- Suarez, S.; Dai, X.J.B. Hyperactivation enhances mouse sperm capacity for penetrating viscoelastic media. Biol. Reprod. 1992, 46, 686–691. [Google Scholar] [CrossRef]
- Suarez, S.S.; Dai, X.B.; de Mott, R.P.; Redfern, K.; Mirando, M.A. Movement characteristics of boar sperm obtained from the oviduct or hyperactivated in vitro. J. Androl. 1992, 13, 75–80. [Google Scholar]
- Guidobaldi, H.A.; Hirohashi, N.; Cubilla, M.; Buffone, M.G.; Giojalas, L.C. An intact acrosome is required for the chemotactic response to progesterone in mouse spermatozoa. Mol. Reprod. Dev. 2017, 84, 310–315. [Google Scholar] [CrossRef]
- Fabro, G.; Rovasio, R.A.; Civalero, S.; Frenkel, A.; Caplan, S.R.; Eisenbach, M.; Giojalas, L.C. Chemotaxis of Capacitated Rabbit Spermatozoa to Follicular Fluid Revealed by a Novel Directionality-Based Assay1. Biol. Reprod. 2002, 67, 1565–1571. [Google Scholar] [CrossRef] [Green Version]
- Gatica, L.V.; Guidobaldi, H.A.; Montesinos, M.M.; Teves, M.E.; Moreno, A.I.; Unates, D.R.; Molina, R.I.; Giojalas, L.C. Picomolar gradients of progesterone select functional human sperm even in subfertile samples. Mol. Hum. Reprod. 2013, 19, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Uñates, D.R.; Guidobaldi, H.A.; Gatica, L.V.; Cubilla, M.A.; Teves, M.E.; Moreno, A.; Giojalas, L.C. Versatile Action of Picomolar Gradients of Progesterone on Different Sperm Subpopulations. PLoS ONE 2014, 9, e91181. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, E.M.; Moreno-Irusta, A.; Guidobaldi, H.A.; Tribulo, H.; Giojalas, L.C. Improved bovine in vitro embryo production with sexed and unsexed sperm selected by chemotaxis. Theriogenology 2018, 122, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Eisenbach, M. Sperm chemotaxis. Rev. Reprod. 1999, 4, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Suarez, S.S.J.B. The oviductal sperm reservoir in mammals: Mechanisms of formation. Biol. Reprod. 1998, 58, 1105–1107. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, R.; Suarez, S.S.J.B. Effect of capacitation on bull sperm binding to homologous oviductal epithelium. Biol. Reprod. 1996, 54, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Yanagimachi, R. Fertility of mammalian spermatozoa: Its development and relativity. Zygote 1994, 2, 371–372. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.H.F. Sperm: Egg ratios and putative molecular signals to modulate gamete interactions in polytocous mammals. Mol. Reprod. Dev. 1993, 35, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Vanderhyden, B.C.; Tonary, A.M.J.B. Differential regulation of progesterone and estradiol production by mouse cumulus and mural granulosa cells by A factor (s) secreted by the oocyte. Biol. Reprod. 1995, 53, 1243–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, Y.; Shimada, M.; Okazaki, T.; Maeda, T.; Terada, T. Production of Progesterone from De Novo-Synthesized Cholesterol in Cumulus Cells and Its Physiological Role During Meiotic Resumption of Porcine Oocytes1. Biol. Reprod. 2003, 68, 1193–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guidobaldi, H.A.; Teves, M.E.; Uñates, D.R.; Anastasía, A.; Giojalas, L.C.J. Progesterone from the cumulus cells is the sperm chemoattractant secreted by the rabbit oocyte cumulus complex. PLoS ONE 2008, 3, e3040. [Google Scholar] [CrossRef] [PubMed]
- Giojalas, L.C.; Guidobaldi, H.A.; Gatica, L.V.; Teves, M.E.; del Mar Montesinos, M.; Uñates, D.R. Device for Diagnosis of Physiologic Status and/or Selection of the Best Spermatozoa of a Semen Sample Based on Chemotaxis, and Procedure of use thereof. U.S. Patent No. 8,993,310, 31 March 2015. [Google Scholar]
- Teves, M.E.; Guidobaldi, H.A.; Uñates, D.R.; Sanchez, R.; Miska, W.; Giojalas, L.C. Progesterone sperm chemoattraction may be modulated by its corticosteroid-binding globulin carrier protein. Fertil. Steril. 2010, 93, 2450–2452. [Google Scholar] [CrossRef]
- Martinez, C.A.; Alvarez-Rodriguez, M.; Wright, D.; Rodriguez-Martinez, H. Does the Pre-Ovulatory Pig Oviduct Rule Sperm Capacitation In Vivo Mediating Transcriptomics of Catsper Channels? Int. J. Mol. Sci. 2020, 21, 1840. [Google Scholar] [CrossRef] [Green Version]
- Lissabet, J.F.B.; Belén, L.H.; Lee-Estevez, M.; Risopatrón, J.; Valdebenito, I.; Figueroa, E.; Farías, J.G. The CatSper channel is present and plays a key role in sperm motility of the Atlantic salmon (Salmo salar). Comp. Biochem. Physiol. Part. A Mol. Integr. Physiol. 2020, 241, 110634. [Google Scholar] [CrossRef]
- Raju, D.N.; Hansen, J.N.; Rassmann, S.; Stüven, B.; Jikeli, J.F.; Strünker, T.; Körschen, H.G.; Möglich, A.; Wachten, D. Cyclic Nucleotide-Specific Optogenetics Highlights Compartmentalization of the Sperm Flagellum into cAMP Microdomains. Cells 2019, 8, 648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darszon, A.; Nishigaki, T.; López-González, I.; Visconti, P.E.; Treviño, C.L. Differences and Similarities: The Richness of Comparative Sperm Physiology. Physiology 2020, 35, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Scott, I.; Reveco, A.R. The Restraint of Bovine Sperm Cell Motility Increases Survival: Role of Extracellular Calcium in the Phenomena. J. Veter. Sci. Technol. 2016, 7, 2. [Google Scholar] [CrossRef]
- Priego-Espinosa, D.A.; Darszon, A.; Guerrero, A.; González-Cota, A.L.; Nishigaki, T.; Martínez-Mekler, G.; Carneiro, J. Modular analysis of the control of flagellar Ca2+-spike trains produced by CatSper and CaV channels in sea urchin sperm. PLoS Comput. Biol. 2020, 16, e1007605. [Google Scholar] [CrossRef]
- Johnson, G.P.; English, A.-M.; Cronin, S.; Hoey, D.A.; Meade, K.G.; Fair, S. Genomic identification, expression profiling, and functional characterization of CatSper channels in the bovine. Biol. Reprod. 2017, 97, 302–312. [Google Scholar] [CrossRef]
- Lehti, M.S.; Sironen, A.J.B. Formation and function of sperm tail structures in association with sperm motility defects. Biol. Reprod. 2017, 97, 522–536. [Google Scholar] [CrossRef] [PubMed]
- Yeste, M.; Llavanera, M.; Mateo-Otero, Y.; Catalán, J.; Bonet, S.; Pinart, E. HVCN1 Channels Are Relevant for the Maintenance of Sperm Motility During In Vitro Capacitation of Pig Spermatozoa. Int. J. Mol. Sci. 2020, 21, 3255. [Google Scholar] [CrossRef]
- Castellano, L.E.; Treviño, C.L.; Rodríguez, D.; Serrano, C.J.; Pacheco, J.; Tsutsumi, V.; Felix, R.; Darszon, A. Transient receptor potential (TRPC) channels in human sperm: Expression, cellular localization and involvement in the regulation of flagellar motility. FEBS Lett. 2003, 541, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Minke, B. Drosophila mutant with a transducer defect. Eur. Biophys. J. 1977, 3, 59–64. [Google Scholar] [CrossRef]
- Montell, C.; Jones, K.; Hafen, E.; Rubin, G. Rescue of the Drosophila phototransduction mutation trp by germline transformation. Science 1985, 230, 1040–1043. [Google Scholar] [CrossRef] [PubMed]
- Wissenbach, U.; Niemeyer, B.A.; Flockerzi, V. TRP channels as potential drug targets. Biol. Cell 2004, 96, 47–54. [Google Scholar] [CrossRef]
- Goodwin, L.O.; Karabinus, D.S.; Pergolizzi, R.G.; Benoff, S. L-type voltage-dependent calcium channel alpha-1C subunit mRNA is present in ejaculated human spermatozoa. Mol. Hum. Reprod. 2000, 6, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Serrano, C.J.; Treviño, C.L.; Felix, R.; Darszon, A. Voltage-dependent Ca(2+) channel subunit expression and immunolocalization in mouse spermatogenic cells and sperm. FEBS Lett. 1999, 462, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, F.; López-González, I.; Serrano, C.; Gasque, G.; de la Vega-Beltrán, J.; Treviño, C.; Darszon, A. Anion channel blockers differentially affect t-type Ca2 currents of mouse spermatogenic cells, α1E currents expressed in Xenopus oocytes and the sperm acrosome reaction. Dev. Genet. 1999, 25, 103–114. [Google Scholar] [CrossRef]
- Liévano, A.; Santi, C.M.; Serrano, C.; Treviño, C.L.; Bellvé, A.R.; Hernández-Cruz, A.; Darszon, A. T-type Ca2+ channels and α 1E expression in spermatogenic cells, and their possible relevance to the sperm acrosome reaction. FEBS Lett. 1996, 388, 150–154. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-Y.; Ahn, H.-J.; Gu, J.-G.; Lee, K.-H.; Kim, J.-S.; Kang, H.-W.; Lee, J.-H. Molecular identification of Ca(2+)channels in human sperm. Exp. Mol. Med. 2003, 35, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Wiesner, B.; Weiner, J.; Middendorff, R.; Hagen, V.; Kaupp, U.B.; Weyand, I. Cyclic Nucleotide-gated Channels on the Flagellum Control Ca2+ Entry into Sperm. J. Cell Biol. 1998, 142, 473–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weyand, I.; Godde, M.; Frings, S.; Weiner, J.; Müller, F.; Altenhofen, W.; Hatt, H.; Kaupp, U.B.; Mueller, F. Cloning and functional expression of a cyclic-nucleotide-gated channel from mammalian sperm. Nat. Cell Biol. 1994, 368, 859–863. [Google Scholar] [CrossRef]
- Gerstner, A.; Zong, X.; Hofmann, F.; Biel, M. Molecular Cloning and Functional Characterization of a New Modulatory Cyclic Nucleotide-Gated Channel Subunit from Mouse Retina. J. Neurosci. 2000, 20, 1324–1332. [Google Scholar] [CrossRef]
- Ren, D.; Navarro, B.; Perez, G.I.; Jackson, A.C.; Hsu, S.; Shi, Q.; Tilly, J.L.; Clapham, D.E. A sperm ion channel required for sperm motility and male fertility. Nat. Cell Biol. 2001, 413, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Quill, T.A.; Ren, D.; Clapham, D.E.; Garbers, D.L. A voltage-gated ion channel expressed specifically in spermatozoa. Proc. Natl. Acad. Sci. USA 2001, 98, 12527–12531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zapata, O.; Ralston, J.; Beltraán, C.; Parys, J.B.; Chen, J.L.; Longo, F.J.; Darszon, A. Inositol triphosphate receptors in sea urchin sperm. Zygote 1997, 5, 355–364. [Google Scholar] [CrossRef]
- Treviño, C.L.; Felix, R.; Castellano, L.E.; Gutierrez, C.; Rodríguez, D.; Pacheco, J.; López-González, I.; Gomora, J.C.; Tsutsumi, V.; Hernández-Cruz, A.; et al. Expression and differential cell distribution of low-threshold Ca2+ channels in mammalian male germ cells and sperm. FEBS Lett. 2004, 563, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Blache, P.; Gros, L.; Salazar, G.; Bataille, D. Cloning and Tissue Distribution of a New Rat Olfactory Receptor-like (OL2). Biochem. Biophys. Res. Commun. 1998, 242, 669–672. [Google Scholar] [CrossRef] [PubMed]
- Drutel, G.; Arrang, J.M.; Diaz, J.; Wisnewsky, C.; Schwartz, K.; Schwartz, J.C. Cloning of OL1, a putative olfactory receptor and its expression in the developing rat heart. Recept. Channels 1995, 3, 33–40. [Google Scholar] [PubMed]
- Ngal, J. The family of genes encoding odorant receptors in the channel catfish. Cell 1993, 72, 657–666. [Google Scholar] [CrossRef]
- Freitag, J.; Krieger, J.; Strotmann, J.; Breer, H. Two classes of olfactory receptors in xenopus laevis. Neuron 1995, 15, 1383–1392. [Google Scholar] [CrossRef] [Green Version]
- Freitag, J.; Ludwig, G.; Andreini, I.; Rossler, P.; Breer, H. Olfactory receptors in aquatic and terrestrial vertebrates. J. Comp. Physiol. A 1998, 183, 635–650. [Google Scholar] [CrossRef]
- Glusman, G.; Yanai, I.; Rubin, I.; Lancet, D. The Complete Human Olfactory Subgenome. Genome Res. 2001, 11, 685–702. [Google Scholar] [CrossRef] [Green Version]
- Godfrey, P.A.; Malnic, B.; Buck, L.B. The mouse olfactory receptor gene family. Proc. Natl. Acad. Sci. USA 2004, 101, 2156–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malnic, B.; Godfrey, P.A.; Buck, L.B. The human olfactory receptor gene family. Proc. Natl. Acad. Sci. USA 2004, 101, 2584–2589. [Google Scholar] [CrossRef] [Green Version]
- Young, J.M.; Friedman, C.; Williams, E.M.; Ross, J.A.; Tonnes-Priddy, L.; Trask, B.J. Different evolutionary processes shaped the mouse and human olfactory receptor gene families. Hum. Mol. Genet. 2002, 11, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Zozulya, S.; Echeverri, F.; Nguyen, T. The human olfactory receptor repertoire. Genome Biol. 2001, 2. [Google Scholar] [CrossRef] [PubMed]
- Hoover, K.C. Evolution of Olfactory Receptors. Adv. Struct. Saf. Stud. 2013, 1003, 241–249. [Google Scholar]
- Zhang, X.; Firestein, S. The olfactory receptor gene superfamily of the mouse. Nat. Neurosci. 2002, 5, 124–133. [Google Scholar] [CrossRef]
- Niimura, Y.; Matsui, A.; Touhara, K. Extreme expansion of the olfactory receptor gene repertoire in African elephants and evolutionary dynamics of orthologous gene groups in 13 placental mammals. Genome Res. 2014, 24, 1485–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, E.; Hoon, M.A.; Mueller, K.L.; Chandrashekar, J.; Ryba, N.J.; Zuker, C.S. A Novel Family of Mammalian Taste Receptors. Cell 2000, 100, 693–702. [Google Scholar] [CrossRef] [Green Version]
- Roper, S.D.; Chaudhari, N. Taste buds: Cells, signals and synapses. Nat. Rev. Neurosci. 2017, 18, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Hofer, D.; Puschel, B.; Drenckhahn, D. Taste receptor-like cells in the rat gut identified by expression of alpha-gustducin. Proc. Natl. Acad. Sci. USA 1996, 93, 6631–6634. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-J.; Depoortere, I.; Hatt, H. Therapeutic potential of ectopic olfactory and taste receptors. Nat. Rev. Drug Discov. 2019, 18, 116–138. [Google Scholar] [CrossRef]
- Nelson, G.; Chandrashekar, J.; Hoon, M.A.; Feng, L.; Zhao, G.; Ryba, N.J.P.; Zuker, C.S. An amino-acid taste receptor. Nat. Cell Biol. 2002, 416, 199–202. [Google Scholar] [CrossRef]
- Niimura, Y. Olfactory Receptor Multigene Family in Vertebrates: From the Viewpoint of Evolutionary Genomics. Curr. Genom. 2012, 13, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Hayden, S.; Bekaert, M.; Crider, T.A.; Mariani, S.; Murphy, W.J.; Teeling, E.C. Ecological adaptation determines functional mammalian olfactory subgenomes. Genome Res. 2009, 20, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mombaerts, P. Genes and ligands for odorant, vomeronasal and taste receptors. Nat. Rev. Neurosci. 2004, 5, 263–278. [Google Scholar] [CrossRef] [PubMed]
- Niimura, Y.; Nei, M. Comparative evolutionary analysis of olfactory receptor gene clusters between humans and mice. Gene 2005, 346, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Axel, R. The Molecular Logic of Smell. Sci. Am. 1995, 273, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Mombaerts, P. Molecular biology of odorant receptors in vertebrates. Annu. Rev. Neurosci. 1999, 22, 487–509. [Google Scholar] [CrossRef] [PubMed]
- Matsui, A.; Go, Y.; Niimura, Y. Degeneration of Olfactory Receptor Gene Repertories in Primates: No Direct Link to Full Trichromatic Vision. Mol. Biol. Evol. 2010, 27, 1192–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niimura, Y.; Nei, M. Extensive Gains and Losses of Olfactory Receptor Genes in Mammalian Evolution. PLoS ONE 2007, 2, e708. [Google Scholar] [CrossRef]
- Nei, M.; Rooney, A.P. Concerted and Birth-and-Death Evolution of Multigene Families. Annu. Rev. Genet. 2005, 39, 121–152. [Google Scholar] [CrossRef] [Green Version]
- Bachmanov, A.A.; Beauchamp, G.K. Taste Receptor Genes. Annu. Rev. Nutr. 2007, 27, 389–414. [Google Scholar] [CrossRef] [Green Version]
- Conte, C.; Ebeling, M.; Marcuz, A.; Nef, P.; Andres-Barquin, P. Identification and characterization of human taste receptor genes belonging to the TAS2R family. Cytogenet. Genome Res. 2002, 98, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Go, Y.; Niimura, Y. Similar Numbers but Different Repertoires of Olfactory Receptor Genes in Humans and Chimpanzees. Mol. Biol. Evol. 2008, 25, 1897–1907. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, T.; Go, Y.; Udono, T.; Morimura, N.; Tomonaga, M.; Hirai, H.; Imai, H. Diversification of Bitter Taste Receptor Gene Family in Western Chimpanzees. Mol. Biol. Evol. 2010, 28, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Gilad, Y.; Man, O.; Pääbo, S. Evolution of Bitter Taste Receptors in Humans and Apes. Mol. Biol. Evol. 2004, 22, 432–436. [Google Scholar] [CrossRef] [Green Version]
- Shi, P.; Shi, P.; Zhang, J.; Zhang, J. Extraordinary Diversity of Chemosensory Receptor Gene Repertoires Among Vertebrates. Chem. Biol. Pteridine Folates 2009, 47, 57–75. [Google Scholar]
- Dong, N.; Jones, G.; Zhang, S. Dynamic evolution of bitter taste receptor genes in vertebrates. BMC Evol. Biol. 2009, 9, 12. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.V.; Chen, M.C.; Rozengurt, E. Genomic organization, expression, and function of bitter taste receptors (T2R) in mouse and rat. Physiol. Genom. 2005, 22, 139–149. [Google Scholar] [CrossRef]
- Li, D.; Zhang, J. Diet Shapes the Evolution of the Vertebrate Bitter Taste Receptor Gene Repertoire. Mol. Biol. Evol. 2014, 31, 303–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, D.T.; Lee, K.; Choi, H.; Choi, M.-K.; Le, M.T.; Song, N.; Kim, J.-H.; Seo, H.G.; Oh, J.-W.; Lee, K.; et al. The complete swine olfactory subgenome: Expansion of the olfactory gene repertoire in the pig genome. BMC Genom. 2012, 13, 584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; de la Cruz, O.; Pinto, J.M.; Nicolae, D.; Firestein, S.; Gilad, Y. Characterizing the expression of the human olfactory receptor gene family using a novel DNA microarray. Genome Biol. 2007, 8, R86. [Google Scholar] [CrossRef] [Green Version]
- Busse, D.; Kudella, P.; Grüning, N.-M.; Gisselmann, G.; Ständer, S.; Luger, T.; Jacobsen, F.; Steinsträßer, L.; Paus, R.; Gkogkolou, P.; et al. A Synthetic Sandalwood Odorant Induces Wound-Healing Processes in Human Keratinocytes via the Olfactory Receptor OR2AT4. J. Investig. Dermatol. 2014, 134, 2823–2832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manteniotis, S.; Wojcik, S.; Göthert, J.R.; Dürig, J.; Dührsen, U.; Gisselmann, G.; Hatt, H. Deorphanization and characterization of the ectopically expressed olfactory receptor OR51B5 in myelogenous leukemia cells. Cell Death Discov. 2016, 2, 16010. [Google Scholar] [CrossRef] [Green Version]
- Maßberg, D.; Simon, A.; Häussinger, D.; Keitel, V.; Gisselmann, G.; Conrad, H.; Hatt, H. Monoterpene (−)-citronellal affects hepatocarcinoma cell signaling via an olfactory receptor. Arch. Biochem. Biophys. 2015, 566, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.V.; Rozengurt, N.; Yang, M.; Young, S.H.; Sinnett-Smith, J.; Rozengurt, E. Expression of bitter taste receptors of the T2R family in the gastrointestinal tract and enteroendocrine STC-1 cells. Proc. Natl. Acad. Sci. USA 2002, 99, 2392–2397. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Zhou, L.; Terwilliger, R.; Newton, S.S.; de Araujo, I.E. Sweet taste signaling functions as a hypothalamic glucose sensor. Front. Integr. Neurosci. 2009, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.R.; Mayer, E.A. Gut-Brain Axis and Behavior. Issues Complem. Feed. 2017, 88, 45–53. [Google Scholar] [CrossRef]
- Liu, X.; Gu, F.; Jiang, L.; Chen, F.; Li, F.; Xin, L.; Li, J.; Fuxue, C. Expression of bitter taste receptor Tas2r105 in mouse kidney. Biochem. Biophys. Res. Commun. 2015, 458, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Hariri, B.M.; Payne, S.J.; Chen, B.; Mansfield, C.; Doghramji, L.J.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; Niv, M.Y.; Lee, R.J. In vitro effects of anthocyanidins on sinonasal epithelial nitric oxide production and bacterial physiology. Am. J. Rhinol. Allergy 2016, 30, 261–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, S.R.; Roura, E.; Thomas, W.G. Extrasensory perception: Odorant and taste receptors beyond the nose and mouth. Pharmacol. Ther. 2014, 142, 41–61. [Google Scholar] [CrossRef]
- Oh, D.Y.; Walenta, E. Omega-3 Fatty Acids and FFAR4; NCBI: Bethesda, MA, USA, 2014; Volume 5, p. 115. [Google Scholar]
- Ekoff, M.; Choi, J.-H.; James, A.; Dahlén, B.; Nilsson, G.; Dahlén, S.-E.; Immunology, C. Bitter taste receptor (TAS2R) agonists inhibit IgE-dependent mast cell activation. J. Allergy Clin. Immunol. 2014, 134, 475–478. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Curto, E.; Milligan, G. Metabolism meets immunity: The role of free fatty acid receptors in the immune system. Biochem. Pharmacol. 2016, 114, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wölfle, U.; Elsholz, F.A.; Kersten, A.; Haarhaus, B.; Müller, W.E.; Schempp, C.M. Expression and Functional Activity of the Bitter Taste Receptors TAS2R1 and TAS2R38 in Human Keratinocytes. Ski. Pharmacol. Physiol. 2015, 28, 137–146. [Google Scholar] [CrossRef]
- Clark, A.A.; Dotson, C.D.; Elson, A.E.T.; Voigt, A.; Boehm, U.; Meyerhof, W.; Steinle, N.I.; Munger, S.D. TAS2R bitter taste receptors regulate thyroid function. FASEB J. 2015, 29, 164–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deckmann, K.; Filipski, K.; Krasteva-Christ, G.; Fronius, M.; Althaus, M.; Rafiq, A.; Papadakis, T.; Renno, L.; Jurastow, I.; Wessels, L.; et al. Bitter triggers acetylcholine release from polymodal urethral chemosensory cells and bladder reflexes. Proc. Natl. Acad. Sci. USA 2014, 111, 8287–8292. [Google Scholar] [CrossRef] [Green Version]
- Mosinger, B.; Redding, K.M.; Parker, M.R.; Yevshayeva, V.; Yee, K.K.; Dyomina, K.; Li, Y.; Margolskee, R.F. Genetic loss or pharmacological blockade of testes-expressed taste genes causes male sterility. Proc. Natl. Acad. Sci. USA 2013, 110, 12319–12324. [Google Scholar] [CrossRef] [Green Version]
- Martin, L.T.P.; Nachtigal, M.W.; Selman, T.; Nguyen, E.; Salsman, J.; Dellaire, G.; Dupré, D.J. Bitter taste receptors are expressed in human epithelial ovarian and prostate cancers cells and noscapine stimulation impacts cell survival. Mol. Cell. Biochem. 2018, 454, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Simon, B.R.; Learman, B.S.; Parlee, S.D.; Scheller, E.L.; Mori, H.; Cawthorn, W.P.; Ning, X.; Krishnan, V.; Ma, Y.L.; Tyrberg, B.; et al. Sweet Taste Receptor Deficient Mice Have Decreased Adiposity and Increased Bone Mass. PLoS ONE 2014, 9, e86454. [Google Scholar] [CrossRef]
- Quesada-López, T.T.; Cereijo, R.R.; Turatsinze, J.-V.; Planavila, A.A.; Cairó, M.M.; Gavaldà-Navarro, A.A.; Peyrou, M.M.; Moure, R.R.; Iglesias, R.R.; Giralt, M.M.; et al. The lipid sensor GPR120 promotes brown fat activation and FGF21 release from adipocytes. Nat. Commun. 2016, 7, 13479. [Google Scholar] [CrossRef] [Green Version]
- Camilleri, M. Peripheral Mechanisms in Appetite Regulation. Gastroenterol. 2015, 148, 1219–1233. [Google Scholar] [CrossRef] [Green Version]
- Augustine, V.; Gokce, S.K.; Oka, Y. Peripheral and Central Nutrient Sensing Underlying Appetite Regulation. Trends Neurosci. 2018, 41, 526–539. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.M.J.T.A. Fenofibrate and Impaired Taste Perception in Type 2 Diabetes. Am. J. Case Rep. 2020, 21, e927647-1. [Google Scholar] [CrossRef]
- Nayak, A.P.; Shah, S.D.; Michael, J.V.; Deshpande, D.A. Bitter Taste Receptors for Asthma Therapeutics. Front. Physiol. 2019, 10, 884. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.J.; Cohen, N.A. Taste receptors in innate immunity. Cell. Mol. Life Sci. 2015, 72, 217–236. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.; Chijoff, E.; Veysey, M.; Keely, S.; Scarlett, C.J.; Lucock, M.; Beckett, E.L. Interactions between taste receptors and the gastrointestinal microbiome in inflammatory bowel disease. J. Nutr. Intermed. Metab. 2019, 18, 100106. [Google Scholar] [CrossRef]
- Calvo, S.S.-C.; Egan, J.M. The endocrinology of taste receptors. Nat. Rev. Endocrinol. 2015, 11, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Kok, B.P.; Galmozzi, A.; Littlejohn, N.K.; Albert, V.; Godio, C.; Kim, W.; Kim, S.M.; Bland, J.S.; Grayson, N.; Fang, M.; et al. Intestinal bitter taste receptor activation alters hormone secretion and imparts metabolic benefits. Mol. Metab. 2018, 16, 76–87. [Google Scholar] [CrossRef]
- Lim, J.; Pullicin, A.J. Oral carbohydrate sensing: Beyond sweet taste. Physiol. Behav. 2019, 202, 14–25. [Google Scholar] [CrossRef]
- Bachmanov, A.; Bosak, N.P.; Lin, C.; Matsumoto, I.; Ohmoto, M.; Reed, D.R.; Nelson, T.J.C.M. Genetics of taste receptors. Curr. Pharm. Des. 2014, 20, 2669–2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vegezzi, G.; Anselmi, L.; Huynh, J.; Barocelli, E.; Rozengurt, E.; Raybould, H.; Sternini, C. Diet-Induced Regulation of Bitter Taste Receptor Subtypes in the Mouse Gastrointestinal Tract. PLoS ONE 2014, 9, e107732. [Google Scholar] [CrossRef]
- Ichimura, A.; Hirasawa, A.; Poulain-Godefroy, O.; Bonnefond, A.; Hara, T.; Yengo, L.; Kimura, I.; Leloire, A.; Liu, N.; Iida, K.; et al. Dysfunction of lipid sensor GPR120 leads to obesity in both mouse and human. Nat. Cell Biol. 2012, 483, 350–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devillier, P.; Naline, E.; Grassin-Delyle, S. The pharmacology of bitter taste receptors and their role in human airways. Pharmacol. Ther. 2015, 155, 11–21. [Google Scholar] [CrossRef]
- Lee, R.J.; Xiong, G.; Kofonow, J.M.; Chen, B.; Lysenko, A.; Jiang, P.; Abraham, V.; Doghramji, L.; Adappa, N.D.; Palmer, J.N.; et al. T2R38 taste receptor polymorphisms underlie susceptibility to upper respiratory infection. J. Clin. Investig. 2012, 122, 4145–4159. [Google Scholar] [CrossRef] [Green Version]
- Linardopoulou, E.; Mefford, H.C.; Nguyen, O.; Friedman, C.; Engh, G.V.D.; Farwell, D.G.; Coltrera, M.; Trask, B.J. Transcriptional activity of multiple copies of a subtelomerically located olfactory receptor gene that is polymorphic in number and location. Hum. Mol. Genet. 2001, 10, 2373–2383. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.B.; Haines, S.L.; Akeson, R.A. Chemoreceptors expressed in taste, olfactory and male reproductive tissues. Gene 1996, 178, 1–5. [Google Scholar] [CrossRef]
- Vanderhaeghen, P.; Schurmans, S.; Vassart, G.; Parmentier, M. Olfactory receptors are displayed on dog mature sperm cells. J. Cell Biol. 1993, 123, 1441–1452. [Google Scholar] [CrossRef] [PubMed]
- Volz, A.; Ehlers, A.; Younger, R.; Forbes, S.; Trowsdale, J.; Schnorr, D.; Beck, S.; Ziegler, A. Complex Transcription and Splicing of Odorant Receptor Genes. J. Biol. Chem. 2003, 278, 19691–19701. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, A.; Dohr, G.; Uchanska-Ziegler, B. Possible Roles for Products of Polymorphic MHC and Linked Olfactory Receptor Genes during Selection Processes in Reproduction*. Am. J. Reprod. Immunol. 2002, 48, 34–42. [Google Scholar] [CrossRef]
- Feldmesser, E.; Olender, T.; Khen, M.; Yanai, I.; Ophir, R.; Lancet, D. Widespread ectopic expression of olfactory receptor genes. BMC Genom. 2006, 7, 121. [Google Scholar] [CrossRef] [Green Version]
- Ichimura, A.; Kadowaki, T.; Narukawa, K.; Togiya, K.; Hirasawa, A.; Tsujimoto, G. In silico approach to identify the expression of the undiscovered molecules from microarray public database: Identification of odorant receptors expressed in non-olfactory tissues. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2008, 377, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Branscomb, A.; Seger, J.; White, R.L. Evolution of odorant receptors expressed in mammalian testes. Genetics 2000, 156, 785–797. [Google Scholar] [PubMed]
- Zhang, X.; Rogers, M.; Tian, H.; Zou, D.-J.; Liu, J.; Ma, M.; Shepherd, G.M.; Firestein, S.J.; Zhang, X.; Zhang, X. High-throughput microarray detection of olfactory receptor gene expression in the mouse. Proc. Natl. Acad. Sci. USA 2004, 101, 14168–14173. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Bahat, A.; Gakamsky, A.; Girsh, E.; Katz, N.; Giojalas, L.C.; Tur-Kaspa, I.; Eisenbach, M. Human sperm chemotaxis: Both the oocyte and its surrounding cumulus cells secrete sperm chemoattractants. Hum. Reprod. 2005, 20, 761–767. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, B.S.; Tur-Kaspa, I.; Dor, J.; Mashiach, S.; Eisenbach, M. Human Sperm Chemotaxis: Is Progesterone a Chemoattractant?1. Biol. Reprod. 1999, 60, 1314–1319. [Google Scholar] [CrossRef] [Green Version]
- Villanueva-Diaz, C.; Vadillo-Ortega, F.; Kably-Ambe, A.; Diaz-Pérez, M.D.L.A.; Krivitzky, S.K. Evidence that human follicular fluid contains a chemoattractant for spermatozoa. Fertil. Steril. 1990, 54, 1180–1182. [Google Scholar] [CrossRef]
- Isobe, T.; Minoura, H.; Tanaka, K.; Shibahara, T.; Hayashi, N.; Toyoda, N. The effect of RANTES on human sperm chemotaxis. Hum. Reprod. 2002, 17, 1441–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Śliwa, L. Heparin as a Chemoattractant for Mouse Spermatozoa. Arch. Androl. 1993, 31, 149–152. [Google Scholar] [CrossRef]
- Śliwa, L. Effect of Heparin on Human Spermatozoa Migration in Vitro. Arch. Androl. 1993, 30, 177–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Śliwa, L.J.A. Substance and ß-Endorphin Act as Possible Chemoattractants of mouse sperm. Arch. Androl. 2001, 46, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Hoang, H.D.; Prasain, J.K.; Dorand, D.; Miller, M.A. A Heterogeneous Mixture of F-Series Prostaglandins Promotes Sperm Guidance in the Caenorhabditis elegans Reproductive Tract. PLoS Genet. 2013, 9, e1003271. [Google Scholar] [CrossRef] [Green Version]
- Edmonds, J.W.; Prasain, J.K.; Dorand, D.; Yang, Y.; Hoang, H.D.; Vibbert, J.; Kubagawa, H.M.; Miller, M.A. Insulin/FOXO Signaling Regulates Ovarian Prostaglandins Critical for Reproduction. Dev. Cell 2010, 19, 858–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado-Oliveira, G.; Lefièvre, L.; Ford, C.; Herrero, M.B.; Barratt, C.; Connolly, T.J.; Nash, K.; Morales-Garcia, A.; Kirkman-Brown, J.; Publicover, S. Mobilisation of Ca2+ stores and flagellar regulation in human sperm by S-nitrosylation: A role for NO synthesised in the female reproductive tract. Development 2008, 135, 3677–3686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimeno, S.; Alquezar-Baeta, C.; Peinado, J.; Nadal, P.; Casao, A.; Cebrián-Pérez, J.A.; Muiño-Blanco, T.; Pérez-Pé, R. Progesterone and estradiol influence on spermatic orientation in ram sperm. Zaragoza 2017, 2017, 395–397. [Google Scholar]
- Cohen-Dayag, A.; Tur-Kaspa, I.; Dor, J.; Mashiach, S.; Eisenbach, M. Sperm capacitation in humans is transient and correlates with chemotactic responsiveness to follicular factors. Proc. Natl. Acad. Sci. USA 1995, 92, 11039–11043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, T.; Salpekar, A.; Monk, M. Expression of a testis-specific member of the olfactory receptor gene family in human primordial germ cells. Mol. Hum. Reprod. 2001, 7, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Eflegel, C.; Evogel, F.; Hofreuter, A.E.; Schreiner, B.S.P.; Eosthold, S.; Eveitinger, S.; Ebecker, C.; Brockmeyer, N.H.; Emuschol, M.; Ewennemuth, G.; et al. Characterization of the Olfactory Receptors Expressed in Human Spermatozoa. Front. Mol. Biosci. 2016, 2, 73. [Google Scholar] [CrossRef] [Green Version]
- Yuan, T.T.-T.; Toy, P.; McClary, J.A.; Lin, R.J.; Miyamoto, N.G.; Kretschmer, P.J. Cloning and genetic characterization of an evolutionarily conserved human olfactory receptor that is differentially expressed across species. Gene 2001, 278, 41–51. [Google Scholar] [CrossRef]
- Veitinger, T.; Riffell, J.R.; Veitinger, S.; Nascimento, J.M.; Triller, A.; Chandsawangbhuwana, C.; Schwane, K.; Geerts, A.; Wunder, F.; Berns, M.W.; et al. Chemosensory Ca2+ Dynamics Correlate with Diverse Behavioral Phenotypes in Human Sperm. J. Biol. Chem. 2011, 286, 17311–17325. [Google Scholar] [CrossRef] [Green Version]
- Itakura, S.; Ohno, K.; Ueki, T.; Sato, K.; Kanayama, N. Expression of Golf in the rat placenta: Possible implication in olfactory receptor transduction. Placenta 2006, 27, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Asai, H.; Kasai, H.; Matsuda, Y.; Yamazaki, N.; Nagawa, F.; Sakano, H.; Tsuboi, A. Genomic Structure and Transcription of a Murine Odorant Receptor Gene: Differential Initiation of Transcription in the Olfactory and Testicular Cells. Biochem. Biophys. Res. Commun. 1996, 221, 240–247. [Google Scholar] [CrossRef]
- Fukuda, N.; Touhara, K. Developmental expression patterns of testicular olfactory receptor genes during mouse spermatogenesis. Genes Cells 2005, 11, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Zhang, X.; Sieli, P.T.; Falduto, M.T.; Torres, K.E.; Rosenfeld, C.S. Contrasting effects of different maternal diets on sexually dimorphic gene expression in the murine placenta. Proc. Natl. Acad. Sci. USA 2010, 107, 5557–5562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fellows, E.; Hazzard, T.; Kutzler, M. Gene Expression in Pre-Term, Pre-Labour and Parturient Canine Placenta. Reprod. Domest. Anim. 2012, 47, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Makeyeva, Y.; Nicol, C.; Ledger, W.L.; Ryugo, D.K. Immunocytochemical Localization of Olfactory-signaling Molecules in Human and Rat Spermatozoa. J. Histochem. Cytochem. 2020, 68, 491–513. [Google Scholar] [CrossRef]
- Walensky, L.D.; Ruat, M.; Bakin, R.E.; Blackshaw, S.; Ronnett, G.V.; Snyder, S.H. Two Novel Odorant Receptor Families Expressed in Spermatids Undergo 5′-Splicing. J. Biol. Chem. 1998, 273, 9378–9387. [Google Scholar] [CrossRef] [Green Version]
- Schultz, N.; Hamra, F.K.; Garbers, D.L. A multitude of genes expressed solely in meiotic or postmeiotic spermatogenic cells offers a myriad of contraceptive targets. Proc. Natl. Acad. Sci. USA 2003, 100, 12201–12206. [Google Scholar] [CrossRef] [Green Version]
- Leese, H.J.; Astley, N.R.; Lambert, D. Glucose and fructose utilization by rat spermatozoa within the uterine lumen. Reprod. 1981, 61, 435–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, S.E.; Gopichandran, N.; Picton, H.M.; Leese, H.J.; Orsi, N.M. Nutrient concentrations in murine follicular fluid and the female reproductive tract. Theriogenology 2005, 64, 992–1006. [Google Scholar] [CrossRef]
- Józwik, M.; Józwik, M.; Teng, C.; Battaglia, F.C.J.H.R. Amino acid, ammonia and urea concentrations in human pre-ovulatory ovarian follicular fluid. Hum. Reprod. 2006, 21, 2776–2782. [Google Scholar] [CrossRef]
- Jóźwik, M.; Jóźwik, M.; Milewska, A.J.; Battaglia, F.C.; Jóźwik, M.J.S. Competitive inhibition of amino acid transport in human preovulatory ovarian follicles. Biomed. Res. Int. 2017, 63, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Suarez, S.; Pacey, A.A. Sperm transport in the female reproductive tract. Hum. Reprod. Updat. 2005, 12, 23–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Blas, G.A.; Darszon, A.; Ocampo, A.Y.; Serrano, C.J.; Castellano, L.E.; Hernández-González, E.O.; Chirinos, M.; Larrea, F.; Beltran, C.; Treviño, C.L. TRPM8, a Versatile Channel in Human Sperm. PLoS ONE 2009, 4, e6095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, D.; Voigt, A.; Widmayer, P.; Borth, H.; Huebner, S.; Breit, A.; Marschall, S.; de Angelis, M.H.; Boehm, U.; Meyerhof, W.; et al. Expression of Tas1 Taste Receptors in Mammalian Spermatozoa: Functional Role of Tas1r1 in Regulating Basal Ca2+ and cAMP Concentrations in Spermatozoa. PLoS ONE 2012, 7, e32354. [Google Scholar] [CrossRef] [Green Version]
- Parks, J.E.; Arion, J.W.; Foote, R.H. Lipids of Plasma Membrane and Outer Acrosomal Membrane from Bovine Spermatozoa. Biol. Reprod. 1987, 37, 1249–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischof, J.C.; Wolkers, W.F.; Tsvetkova, N.M.; Oliver, A.E.; Crowe, J.H. Lipid and protein changes due to freezing in Dunning AT-1 cells. Cryobiology 2002, 45, 22–32. [Google Scholar] [CrossRef]
- Vadnais, M.L.; Althouse, G.C. Characterization of capacitation, cryoinjury, and the role of seminal plasma in porcine sperm. Theriogenology 2011, 76, 1508–1516. [Google Scholar] [CrossRef] [PubMed]
- Yeste, M. Sperm cryopreservation update: Cryodamage, markers, and factors affecting the sperm freezability in pigs. Theriogenology 2016, 85, 47–64. [Google Scholar] [CrossRef]
- Yeste, M.; Bonet, S.; Rodríguez-Gil, J.E. Artificial insemination with frozen-thawed boar sperm. Mol. Reprod. Dev. 2017, 84, 802–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehr, J.; Meyer, D.; Widmayer, P.; Borth, H.C.; Ackermann, F.; Wilhelm, B.; Gudermann, T.; Boekhoff, I. Expression of the G-protein α-subunit gustducin in mammalian spermatozoa. J. Comp. Physiol. A 2006, 193, 21–34. [Google Scholar] [CrossRef]
- Kiuchi, S.; Yamada, T.; Kiyokawa, N.; Saito, T.; Fujimoto, J.; Yasue, H. Genomic structure of swine taste receptor family 1 member 3, TAS1R3, and its expression in tissues. Cytogenet. Genome Res. 2006, 115, 51–61. [Google Scholar] [CrossRef]
- Gibson, A.D.; Garbers, D.L. Guanylyl Cyclases as a Family of Putative Odorant Receptors. Annu. Rev. Neurosci. 2000, 23, 417–439. [Google Scholar] [CrossRef]
- Luddi, A.; Governini, L.; Wilmskötter, D.; Gudermann, T.; Boekhoff, I.; Piomboni, P. Taste Receptors: New Players in Sperm Biology. Int. J. Mol. Sci. 2019, 20, 967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boekhoff, I.; Tareilus, E.; Strotmann, J.; Breer, H. Rapid activation of alternative second messenger pathways in olfactory cilia from rats by different odorants. EMBO J. 1990, 9, 2453–2458. [Google Scholar] [CrossRef]
- Huque, T.; Bruch, R.C. Odorant- and guanine nucleotide-stimulated phosphoinositide turnover in olfactory cilia. Biochem. Biophys. Res. Commun. 1986, 137, 36–42. [Google Scholar] [CrossRef]
- Ronnett, G.V.; Cho, H.; Hester, L.D.; Wood, S.F.; Snyder, S.H. Odorants differentially enhance phosphoinositide turnover and adenylyl cyclase in olfactory receptor neuronal cultures. J. Neurosci. 1993, 13, 1751–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sklar, P.B.; Anholt, R.R.; Snyder, S.H. The odorant-sensitive adenylate cyclase of olfactory receptor cells. Differential stimulation by distinct classes of odorants. J. Biol. Chem. 1986, 261, 15538–15543. [Google Scholar] [CrossRef]
- Spehr, M.; Wetzel, C.H.; Hatt, H.; Ache, B.W. 3-Phosphoinositides Modulate Cyclic Nucleotide Signaling in Olfactory Receptor Neurons. Neuron 2002, 33, 731–739. [Google Scholar] [CrossRef]
- Visconti, P.E.; Galantino-Homer, H.; Ning, X.; Moore, G.D.; Valenzuela, J.P.; Jorgez, C.J.; Alvarez, J.G.; Kopf, G.S.J.J. Cholesterol efflux-mediated signal transduction in mammalian sperm: β-cyclodextrins initiate transmembrane signaling leading to an increase in protein tyrosine phosphorylation and capacitation. J. Biol. Chem. 1999, 274, 3235–3242. [Google Scholar] [CrossRef] [Green Version]
- Visconti, P.E.; Moore, G.D.; Bailey, J.L.; Leclerc, P.; Connors, S.A.; Pan, D.; Olds-Clarke, P.; Kopf, G.S.J.D. Capacitation of mouse spermatozoa. II. Protein tyrosine phosphorylation and capacitation are regulated by a cAMP-dependent pathway. Development 1995, 121, 1139–1150. [Google Scholar] [PubMed]
- Lishko, P.V.; Botchkina, I.L.; Kirichok, Y. Progesterone activates the principal Ca2+ channel of human sperm. Nat. Cell Biol. 2011, 471, 387–391. [Google Scholar] [CrossRef]
- Strünker, T.; Goodwin, N.; Brenker, C.; Kashikar, N.D.; Weyand, I.; Seifert, R.; Kaupp, U.B. The CatSper channel mediates progesterone-induced Ca2+ influx in human sperm. Nat. Cell Biol. 2011, 471, 382–386. [Google Scholar] [CrossRef]
- Visconti, P.; Westbrook, V.; Chertihin, O.; Demarco, I.; Sleight, S.; Diekman, A. Novel signaling pathways involved in sperm acquisition of fertilizing capacity. J. Reprod. Immunol. 2002, 53, 133–150. [Google Scholar] [CrossRef]
- Signorelli, J.; Diaz, E.S.; Morales, P. Kinases, phosphatases and proteases during sperm capacitation. Cell Tissue Res. 2012, 349, 765–782. [Google Scholar] [CrossRef]
- Sagare-Patil, V.; Vernekar, M.; Galvankar, M.; Modi, D. Progesterone utilizes the PI3K-AKT pathway in human spermatozoa to regulate motility and hyperactivation but not acrosome reaction. Mol. Cell. Endocrinol. 2013, 374, 82–91. [Google Scholar] [CrossRef]
- Belluscio, L.; Gold, G.H.; Nemes, A.; Axel, R. Mice Deficient in Golf Are Anosmic. Neuron 1998, 20, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Brunet, L.J.; Gold, G.H.; Ngai, J. General Anosmia Caused by a Targeted Disruption of the Mouse Olfactory Cyclic Nucleotide–Gated Cation Channel. Neuron 1996, 17, 681–693. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.T.; Trinh, K.; Hacker, B.; Chan, G.C.; Lowe, G.; Gaggar, A.; Xia, Z.; Gold, G.H.; Storm, D.R. Disruption of the Type III Adenylyl Cyclase Gene Leads to Peripheral and Behavioral Anosmia in Transgenic Mice. Neuron 2000, 27, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Pérez, C.A.; Huang, L.; Rong, M.; Kozak, J.A.; Preuss, A.K.; Zhang, H.; Max, M.; Margolskee, R.F. A transient receptor potential channel expressed in taste receptor cells. Nat. Neurosci. 2002, 5, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Clapp, T.R.; Trubey, K.R.; Vandenbeuch, A.; Stone, L.M.; Margolskee, R.F.; Chaudhari, N.; Kinnamon, S.C. Tonic activity of Gα-gustducin regulates taste cell responsivity. FEBS Lett. 2008, 582, 3783–3787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taruno, A.; Vingtdeux, V.; Ohmoto, M.; Ma, Z.; Dvoryanchikov, G.; Li, A.; Adrien, L.; Zhao, H.; Leung, S.; Abernethy, M.; et al. CALHM1 ion channel mediates purinergic neurotransmission of sweet, bitter and umami tastes. Nat. Cell Biol. 2013, 495, 223–226. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Taruno, A.; Ohmoto, M.; Jyotaki, M.; Lim, J.C.; Miyazaki, H.; Niisato, N.; Marunaka, Y.; Lee, R.J.; Hoff, H.; et al. CALHM3 Is Essential for Rapid Ion Channel-Mediated Purinergic Neurotransmission of GPCR-Mediated Tastes. Neuron 2018, 98, 547–561.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, W. Fundamental aspects of sperm cryobiology: The importance of species and individual differences. Theriogenology 2000, 53, 47–58. [Google Scholar] [CrossRef]
- Medeiros, C.; Forell, F.; Oliveira, A.; Rodrigues, J. Current status of sperm cryopreservation: Why isn’t it better? Theriogenology 2002, 57, 327–344. [Google Scholar] [CrossRef]
- Susko-Parrish, J.L.; Uguz, C.; First, N.L. Differences in the Role of Cyclic Adenosine 3′,5′-Monophosphate during Capacitation of Bovine Sperm by Heparin or Oviduct Fluid1. Biol. Reprod. 1994, 51, 1099–1108. [Google Scholar] [CrossRef] [Green Version]
- Leemans, B.; Stout, T.A.E.; de Schauwer, C.; Heras, S.; Nelis, H.; Hoogewijs, M.; van Soom, A.; Gadella, B.M. Update on mammalian sperm capacitation: How much does the horse differ from other species? Reproduction 2019, 157, R181–R197. [Google Scholar] [CrossRef] [Green Version]
- Yamashiro, H.; Toyomizu, M.; Toyama, N.; Aono, N.; Sakurai, M.; Hiradate, Y.; Yokoo, M.; Moisyadi, S.; Sato, E. Extracellular ATP and Dibutyryl cAMP Enhance the Freezability of Rat Epididymal Sperm. J. Am. Assoc. Lab. Anim. Sci. 2010, 49, 167–172. [Google Scholar] [PubMed]
- Lee-Estevez, M.; Herrera, L.; Díaz, R.; Beltrán, J.; Figueroa, E.; Dumorné, K.; Ulloa-Rodríguez, P.; Short, S.; Risopatrón, J.; Valdebenito, I.; et al. Effects of cryopreservation on cAMP-dependent protein kinase and AMP-activated protein kinase in Atlantic salmon (Salmo salar) spermatozoa: Relation with post-thaw motility. Anim. Reprod. Sci. 2019, 209, 106133. [Google Scholar] [CrossRef]
- De Llera, A.H.; Martin-Hidalgo, D.; Gil, M.C.; Garcia-Marin, L.J.; Bragado, M.J. AMP-Activated Kinase AMPK Is Expressed in Boar Spermatozoa and Regulates Motility. PLoS ONE 2012, 7, e38840. [Google Scholar] [CrossRef] [Green Version]
- Fraser, L.R. The “switching on” of mammalian spermatozoa: Molecular events involved in promotion and regulation of capacitation. Mol. Reprod. Dev. 2009, 77, 197–208. [Google Scholar] [CrossRef]
- Wang, Q.; Liszt, K.I.; Depoortere, I. Extra-oral bitter taste receptors: New targets against obesity? Peptide 2020, 127, 170284. [Google Scholar] [CrossRef]
- Spehr, M.; Schwane, K.; Riffell, J.A.; Zimmer, R.K.; Hatt, H. Odorant receptors and olfactory-like signaling mechanisms in mammalian sperm. Mol. Cell. Endocrinol. 2006, 250, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Ran, M.-X.; Li, Y.; Zhang, Y.; Liang, K.; Ren, Y.-N.; Zhang, M.; Zhou, G.-B.; Zhou, Y.-M.; Wu, K.; Wang, C.-D.; et al. Transcriptome Sequencing Reveals the Differentially Expressed lncRNAs and mRNAs Involved in Cryoinjuries in Frozen-Thawed Giant Panda (Ailuropoda melanoleuca) Sperm. Int. J. Mol. Sci. 2018, 19, 3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewcock, J.W.; Reed, R.R. A feedback mechanism regulates monoallelic odorant receptor expression. Proc. Natl. Acad. Sci. USA 2004, 101, 1069–1074. [Google Scholar] [CrossRef] [Green Version]
- Serizawa, S.; Miyamichi, K.; Sakano, H. Negative Feedback Regulation Ensures the One Neuron-One Receptor Rule in the Mouse Olfactory System. Chem. Senses 2005, 30, i99–i100. [Google Scholar] [CrossRef] [PubMed]
- Carlson, A.E.; Westenbroek, R.E.; Quill, T.; Ren, D.; Clapham, D.E.; Hille, B.; Garbers, D.L.; Babcock, D.F. CatSper1 required for evoked Ca2+ entry and control of flagellar function in sperm. Proc. Natl. Acad. Sci. USA 2003, 100, 14864–14868. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Location | Specie | ||||
|---|---|---|---|---|---|
| Human | Bovine | Dog | Mouse | Rat | |
| Head | Cav2.1c, Cav3.1d, Cav3.3d, TRPC1a, TRPC3a, TRPC4a, TRPC6a, IP3R j | IP3R j | IP3R j | Cav1.2b, Cav2.1c, Cav2.2d, Cav2.3d, Cav3.1d, Cav3.2d, TRPC2a, IP3R j | IP3R j |
| Mid-piece | Cav1.2b, Cav2.3d, Cav3.1d, Cav3.2d, Cav3.3d, TRPC3a, TRPC4a, TRPC6a | CNGA3e | Catsper2h, Cav2.1c, Cav2.3d, Cav3.1d, Cav3.3d, TRPC1a, TRPC3a, TRPC6a | ||
| Upper tail | Cav1.2b, Cav3.1d, Cav3.2d, TRPC1a, TRPC4a, TRPC6a | CNGA3e, CNGB1f | Catsper1g, Catsper2h, Cav1.2b, Cav2.2d, Cav2.3d, Cav3.1k, Cav3.2d, Cav3.3d, TRPC1a, TRPC3a | ||
| Lower tail | Cav2.3d | CNGA3e | Cav3.2d, TRPC3a | ||
| Specie | Olfactory Genes Distribution | Reference | Taste Genes distribution | Reference | ||
|---|---|---|---|---|---|---|
| Intact genes | Pseudogenes | Intact genes | Pseudogenes | |||
| Human | 396 | 425 | [99] | 38 (T2R) | 16 (T2R) | [102,103] |
| Chimpanzee | 380 | 414 | [104] | 28 (T2R) | 10 (T2R) | [105,106] |
| Cow | 1186 | 1057 | [87] | 12 (T2R) | 15 (T2R) | [107] |
| Dog | 811 | 278 | [100] | 15 (T2R) | 5 (T2R) | [107] |
| Horse | 1066 | 1569 | [87] | 19 (T2R) | 36 (T2R) | [108] |
| Mouse | 1130 | 236 | [87] | 35 (T2R) | 5 (T2R) | [102] |
| Rat | 1207 | 508 | [100] | 36 (T2R) | 7 (T2R) | [109] |
| Rabbit | 768 | 256 | [87] | 28 (T2R) | 13 (T2R) | [110] |
| Pig | 1113 | 188 | [111] | 15 (T2R) | 7 (T2R) | [110] |
| Species | Organ | Receptors | Analysis Method | Reference |
|---|---|---|---|---|
| Human | Sperm | OR7E24, OR4S1, OR4C13, OR1I1, HT2, OR1D4, OR51E1, OR51E2, OR6B2, OR10J1, OR2H1/2, OR2W3 | Confocal microscopy, WB, ICS | [10,167,168] |
| Prostate | OR51E2, o1r59, Olfr78, | RT-PCR, WB, NB | [8,169] | |
| Testis | OR4C13, OR7A5, OR4D1, OR1D2, OR1D2, OR4D1, OR1D2, OR1E1 hOR17-4, hOR17-2, Olfr16, OR4N4, OR3A2, OR10J1 | RT-PCR, q-PCR, Ca-Imaging, RNA-seq | [6,9,10,170] | |
| Placenta | hOR-17 | RT-PCR | [171] | |
| Mouse | Sperm | MOR23 | RT-PCR, in situ hybridization | [11] |
| Prostate | N/A | N/A | N/A | |
| Testis | MOR23, MOR244-3, MOR139-3, MOR248-11, MOR267-13, MOR283-1, MOR8-1, MOR31-2, OR10J5, | RT-PCR, NB | [11,12,172,173] | |
| Placenta | Olfr154, Olfr520, Olfr433, O1fr381 | Microarray | [174] | |
| Dog | Sperm | DTMT | RNase protection assay, WB | [44,148] |
| Prostate | N/A | N/A | N/A | |
| Testis | DTPCR64, HGMP07, DTMT, OR1E2, | RT-PCR | [5,12] | |
| Placenta | cOR2AV3 | Microarray, RT-PCR | [175] | |
| Rat | Sperm | Putative olfactory Proteins GRK3, beta-arrestin2, Golf | WB, ICC, IH | [13,176] |
| Prostate | N/A | N/A | N/A | |
| Testis | OD1, OD2, Olr825, Olr1696 | RT-PCR, WB, IH, ISH | [13,177] | |
| Placenta | O1r1767, Olr1513, Olr1687, Olr1571 | RT-PCR | [171] |
| Specie | Location | Receptors/Protein | Analysis Method | Reference |
|---|---|---|---|---|
| Human | Sperm | Tas1r1, Tas1r3, α-gustducin, α-transducin | RT-PCR, WB | [14,185] |
| Testis | Tas1r14 | Droplets digital PCR | [14] | |
| Mouse | Testis | Tas2r102, Tas2r 105, Tas2r 106, Tas2r 113, Tas2r 114, Tas2r 116, Tas2r 124, Tas2r 125, Tas2r 134, Tas2r 135, Tas2r 136 | RT-PCR | [15] |
| Boar | Sperm | α-gustducin, α-transducin | WB, immunohistochemistry | [16] |
| Testis | T1R3 | RT-PCR, in situ hybridization | [192] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, M.A.; Wang, Y.; Qin, Z.; Yuan, X.; Zhang, Y.; Zeng, C. Odorant and Taste Receptors in Sperm Chemotaxis and Cryopreservation: Roles and Implications in Sperm Capacitation, Motility and Fertility. Genes 2021, 12, 488. https://doi.org/10.3390/genes12040488
Ali MA, Wang Y, Qin Z, Yuan X, Zhang Y, Zeng C. Odorant and Taste Receptors in Sperm Chemotaxis and Cryopreservation: Roles and Implications in Sperm Capacitation, Motility and Fertility. Genes. 2021; 12(4):488. https://doi.org/10.3390/genes12040488
Chicago/Turabian StyleAli, Malik Ahsan, Yihan Wang, Ziyue Qin, Xiang Yuan, Yan Zhang, and Changjun Zeng. 2021. "Odorant and Taste Receptors in Sperm Chemotaxis and Cryopreservation: Roles and Implications in Sperm Capacitation, Motility and Fertility" Genes 12, no. 4: 488. https://doi.org/10.3390/genes12040488