
The Role of Knockout Olfactory Receptor Genes in Odor Discrimination

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Results
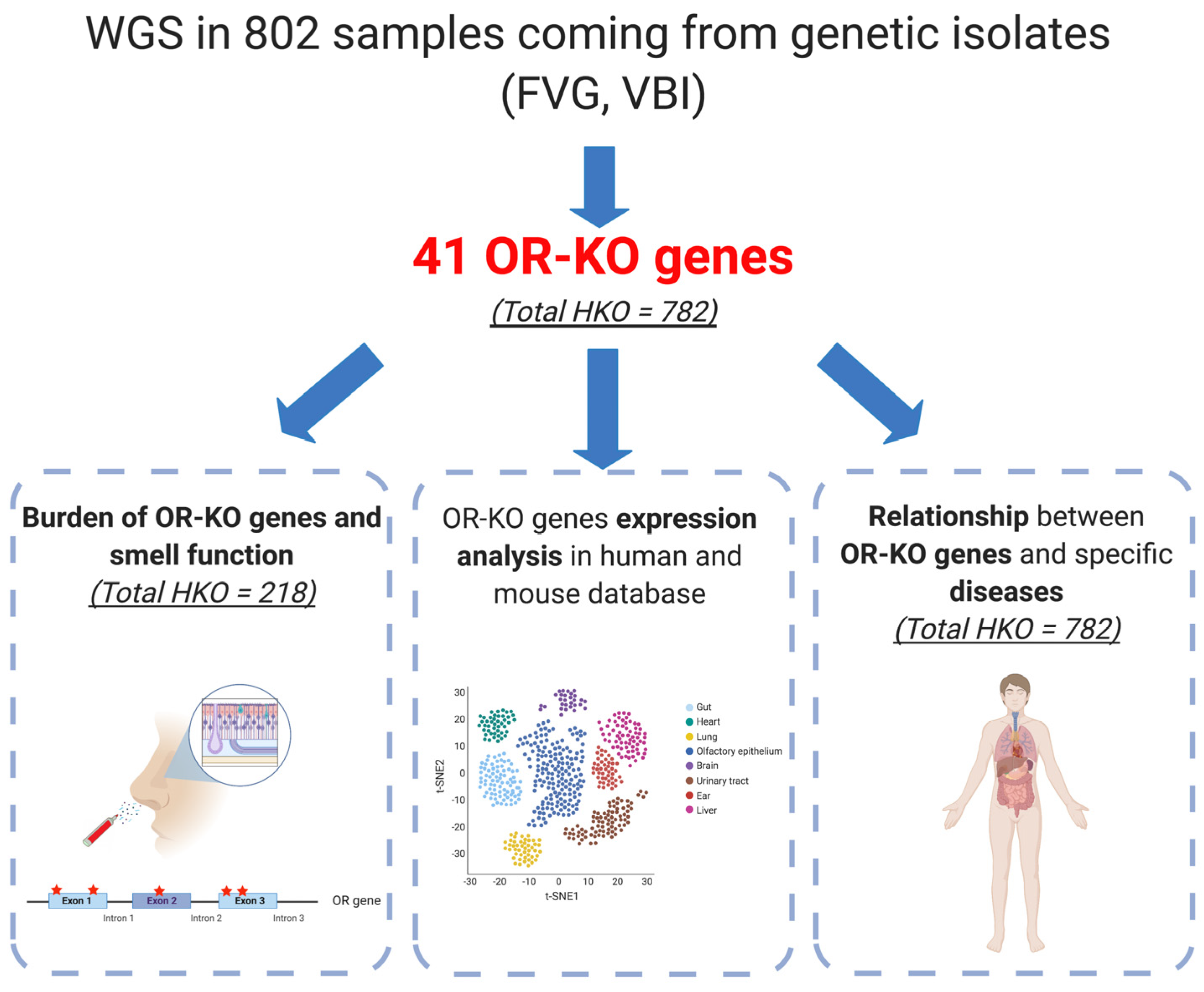
2.1. Dataset Overview and Characterization of Olfactory Receptor Knockout (OR-KO) Variants
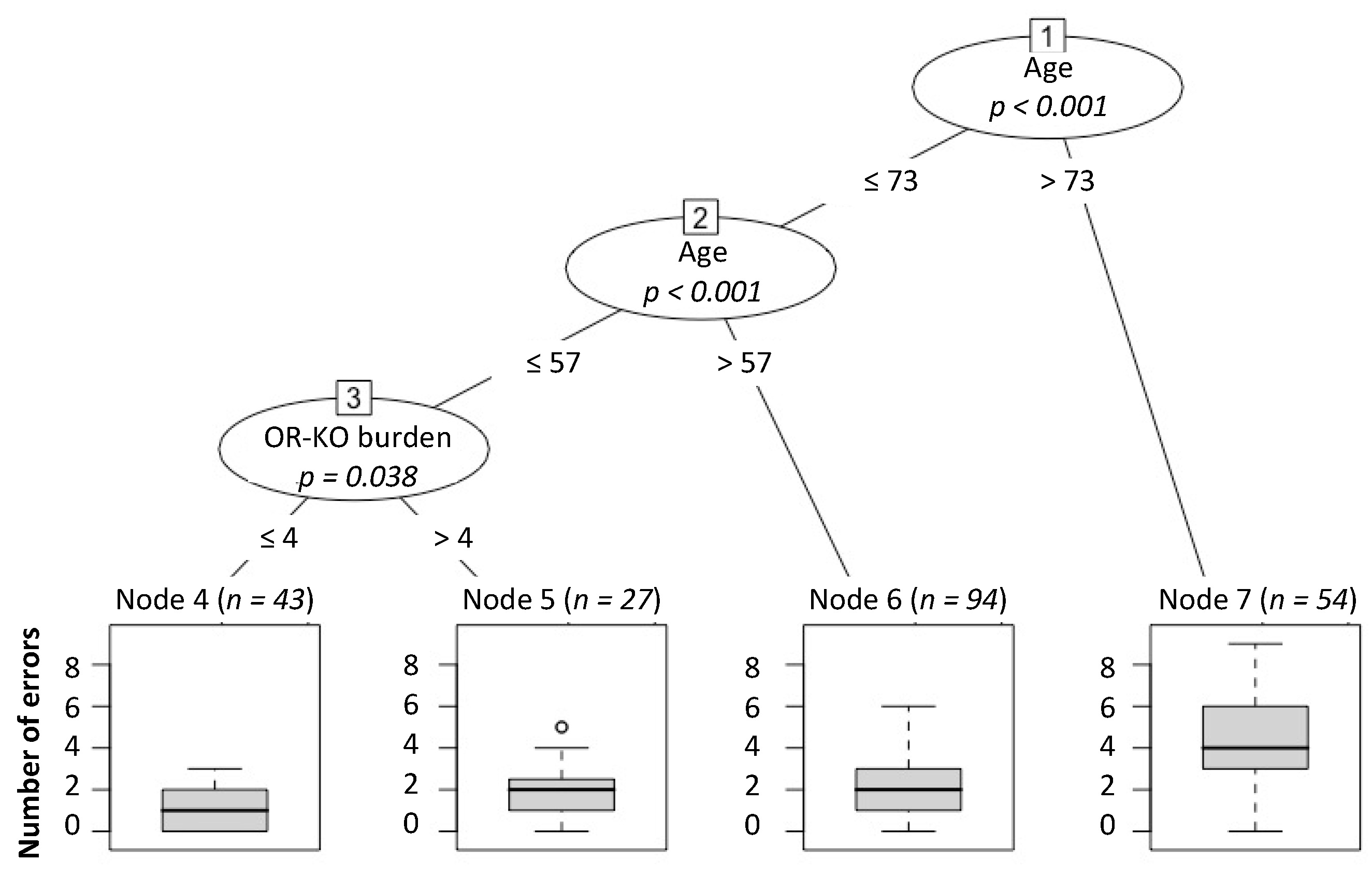
2.2. Relationship between OR-KO Genes’ Burden and Smell Performance
2.3. Expression Patterns of OR-KO Genes
2.4. Relationship with Pathologies
3. Discussion
4. Materials and Methods
4.1. Identification of OR-KO Genes and Comparison with External Databases
4.2. Clinical Evaluation
4.3. Smell Functionality Assessment
4.4. Relationship between Smell Performance and the Burden of OR-KO Genes
4.5. Expression of ORs in Human and Mouse
4.6. Relationship with Pathologies
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buck, L.B. Unraveling the Sense of Smell (Nobel Lecture). Angew. Chem. Int. Ed. 2005, 44, 6128–6140. [Google Scholar] [CrossRef]
- Roberts, S.C.; Havlíček, J.; Schaal, B. Human olfactory communication: Current challenges and future prospects. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20190258. [Google Scholar] [CrossRef]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Bushdid, C.; Magnasco, M.O.; Vosshall, L.B.; Keller, A. Humans Can Discriminate More than 1 Trillion Olfactory Stimuli. Science 2014, 343, 1370–1372. [Google Scholar] [CrossRef] [Green Version]
- Child, K.M.; Herrick, D.B.; Schwob, J.E.; Holbrook, E.H.; Jang, W. The Neuroregenerative Capacity of Olfactory Stem Cells Is Not Limitless: Implications for Aging. J. Neurosci. 2018, 38, 6806–6824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malnic, B.; Hirono, J.; Sato, T.; Buck, L.B. Combinatorial Receptor Codes for Odors. Cell 1999, 96, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Glusman, G.; Yanai, I.; Rubin, I.; Lancet, D. The Complete Human Olfactory Subgenome. Genome Res. 2001, 11, 685–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olender, T.; Feldmesser, E.; Atarot, T.; Eisenstein, M.; Lancet, D. The olfactory receptor universe—From whole genome analysis to structure and evolution. Genet. Mol. Res. 2004, 3, 545–553. [Google Scholar]
- Niimura, Y.; Nei, M. Extensive Gains and Losses of Olfactory Receptor Genes in Mammalian Evolution. PLoS ONE 2007, 2, e708. [Google Scholar] [CrossRef] [PubMed]
- Olender, T.; Fuchs, T.; Linhart, C.; Shamir, R.; Adams, M.; Kalush, F.; Khen, M.; Lancet, D. The canine olfactory subgenome. Genomics 2004, 83, 361–372. [Google Scholar] [CrossRef]
- Jenkins, E.K.; DeChant, M.T.; Perry, E.B. When the Nose Doesn’t Know: Canine Olfactory Function Associated with Health, Management, and Potential Links to Microbiota. Front. Vet. Sci. 2018, 5, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouquier, S.; Blancher, A.; Giorgi, D. The olfactory receptor gene repertoire in primates and mouse: Evidence for reduction of the functional fraction in primates. Proc. Natl. Acad. Sci. USA 2000, 97, 2870–2874. [Google Scholar] [CrossRef] [Green Version]
- Verbeurgt, C.; Wilkin, F.; Tarabichi, M.; Gregoire, F.; Dumont, J.E.; Chatelain, P. Profiling of olfactory receptor gene ex-pression in whole human olfactory mucosa. PLoS ONE 2014, 9, e96333. [Google Scholar] [CrossRef] [PubMed]
- Malnic, B.; Godfrey, P.A.; Buck, L.B. The human olfactory receptor gene family. Proc. Natl. Acad. Sci. USA 2004, 101, 2584–2589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.J. System-Wide Expression and Function of Olfactory Receptors in Mammals. Genom. Inform. 2018, 16, 2–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasin-Brumshtein, Y.; Lancet, D.; Olender, T. Human olfaction: From genomic variation to phenotypic diversity. Trends Genet. 2009, 25, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Attems, J.; Walker, L.; Jellinger, K.A. Olfaction and Aging: A Mini-Review. Gerontology 2015, 61, 485–490. [Google Scholar] [CrossRef]
- Marin, C.; Vilas, D.; Langdon, C.; Alobid, I.; López-Chacón, M.; Haehner, A.; Hummel, T.; Mullol, J. Olfactory Dysfunction in Neu-rodegenerative Diseases. Curr. Allergy Asthma Rep. 2018, 18, 42. [Google Scholar] [CrossRef]
- Howell, J.; Costanzo, R.M.; Reiter, E.R. Head trauma and olfactory function. World J. Otorhinolaryngol. Head Neck Surg. 2018, 4, 39–45. [Google Scholar] [CrossRef]
- Kebir, S.; Hattingen, E.; Niessen, M.; Rauschenbach, L.; Fimmers, R.; Hummel, T.; Schäfer, N.; Lazaridis, L.; Kleinschnitz, C.; Herrlinger, U.; et al. Olfactory function as an independent prognostic factor in glioblastoma. Neurology 2020, 94, e529–e537. [Google Scholar] [CrossRef]
- Kawabata, T.; Takeuchi, K.; Nagata, Y.; Ishikawa, T.; Choo, J.; Sato, Y.; Tambara, M.; Teranishi, M.; Wakabayashi, T. Preservation of Olfactory Function Following Endoscopic Single-Nostril Transseptal Transsphenoidal Surgery. World Neurosurg. 2019, 132, e665–e669. [Google Scholar] [CrossRef] [PubMed]
- Skuja, S.; Zieda, A.; Ravina, K.; Chapenko, S.; Roga, S.; Teteris, O.; Groma, V.; Murovska, M. Structural and Ultrastructural Alterations in Human Olfactory Pathways and Possible Associations with Herpesvirus 6 Infection. PLoS ONE 2017, 12, e0170071. [Google Scholar] [CrossRef] [Green Version]
- Keller, A.; Zhuang, H.; Chi, Q.; Vosshall, L.B.; Matsunami, H. Genetic variation in a human odorant receptor alters odour perception. Nat. Cell Biol. 2007, 449, 468–472. [Google Scholar] [CrossRef]
- Gisladottir, R.S.; Ivarsdottir, E.V.; Helgason, A.; Jonsson, L.; Hannesdottir, N.K.; Rutsdottir, G.; Arnadottir, G.A.; Skuladottir, A.; Jonsson, B.A.; Norddahl, G.L.; et al. Sequence Variants in TAAR5 and Other Loci Affect Human Odor Perception and Naming. Curr. Biol. 2020, 30, 4643–4653.e3. [Google Scholar] [CrossRef] [PubMed]
- Trimmer, C.; Keller, A.; Murphy, N.R.; Snyder, L.L.; Willer, J.R.; Nagai, M.H.; Katsanis, N.; Vosshall, L.B.; Matsunami, H.; Mainland, J.D. Genetic variation across the human olfactory receptor repertoire alters odor perception. Proc. Natl. Acad. Sci. USA 2019, 116, 9475–9480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McRae, J.F.; Jaeger, S.R.; Bava, C.M.; Beresford, M.K.; Hunter, D.; Jia, Y.; Chheang, S.L.; Jin, D.; Peng, M.; Gamble, J.C.; et al. Identification of Regions Associated with Variation in Sensitivity to Food-Related Odors in the Human Genome. Curr. Biol. 2013, 23, 1596–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knaapila, A.; Zhu, G.; Medland, S.E.; Wysocki, C.J.; Montgomery, G.W.; Martin, N.G.; Wright, M.J.; Reed, D.R. A Genome-Wide Study on the Perception of the Odorants Androstenone and Galaxolide. Chem. Senses 2012, 37, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, N.; MacPherson, J.M.; Tung, J.Y.; Hon, L.S.; Naughton, B.; Saxonov, S.; Avey, L.; Wojcicki, A.; Pe’Er, I.; Mountain, J. Web-Based, Participant-Driven Studies Yield Novel Genetic Associations for Common Traits. PLoS Genet. 2010, 6, e1000993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaeger, S.R.; McRae, J.F.; Bava, C.M.; Beresford, M.K.; Hunter, D.; Jia, Y.; Chheang, S.L.; Jin, D.; Peng, M.; Gamble, J.C.; et al. A Mendelian Trait for Olfactory Sensitivity Affects Odor Experience and Food Selection. Curr. Biol. 2013, 23, 1601–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mainland, J.D.; Keller, A.; Li, Y.R.; Zhou, T.; Trimmer, C.; Snyder, L.L.; Moberly, A.H.; Adipietro, K.A.; Liu, W.L.L.; Zhuang, H.; et al. The missense of smell: Functional variability in the human odorant receptor repertoire. Nat. Neurosci. 2014, 17, 114–120. [Google Scholar] [CrossRef] [Green Version]
- McRae, J.F.; Mainland, J.D.; Jaeger, S.R.; Adipietro, K.A.; Matsunami, H.; Newcomb, R.D. Genetic Variation in the Odorant Receptor OR2J3 Is Associated with the Ability to Detect the “Grassy” Smelling Odor, cis-3-hexen-1-ol. Chem. Senses 2012, 37, 585–593. [Google Scholar] [CrossRef] [Green Version]
- Cocca, M.; Barbieri, C.; Concas, M.P.; Robino, A.; Brumat, M.; Gandin, I.; Trudu, M.; Sala, C.F.; Vuckovic, D.; Giorgia, G.; et al. A bird’s-eye view of Italian ge-nomic variation through whole-genome sequencing. Eur. J. Hum. Genet. 2020, 28, 435–444. [Google Scholar] [CrossRef]
- Van der Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; Del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From FastQ data to high confidence variant calls: The Genome Analysis Toolkit best practices pipeline. Curr. Protoc. Bioinform. 2013, 43, 11.10.1–11.10.33. [Google Scholar]
- The 1000 Genomes Project Consortium. A global reference for human genetic variation. Nature 2015, 526, 68–74.
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alföldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, R.C.; Casajuana-Martin, N.; García-Recio, A.; Alcántara, L.; Pardo, L.; Campillo, M.; Gonzalez, A. The mutational landscape of human olfactory G protein-coupled receptors. BMC Biol. 2021, 19, 21. [Google Scholar] [CrossRef]
- Saraiva, L.R.; Riveros-McKay, F.; Mezzavilla, M.; Abou-Moussa, E.H.; Arayata, C.J.; Makhlouf, M.; Trimmer, C.; Ibarra-Soria, X.; Khan, M.; Van Gerven, L.; et al. A tran-scriptomic atlas of mammalian olfactory mucosae reveals an evolutionary influence on food odor detection in humans. Sci. Adv. 2019, 5, eaax0396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dintica, C.S.; Marseglia, A.; Rizzuto, D.; Wang, R.; Seubert, J.; Arfanakis, K.; Bennett, D.A.; Xu, W. Impaired olfaction is associated with cognitive decline and neurodegeneration in the brain. Neurology 2019, 92, e700–e709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doty, R.L.; Hawkes, C.H. Chemosensory dysfunction in neurodegenerative diseases. Handb. Clin. Neurol. 2019, 164, 325–360. [Google Scholar] [CrossRef] [PubMed]
- Robak, L.A.; Jansen, I.E.; van Rooij, J.; Uitterlinden, A.G.; Kraaij, R.; Jankovic, J.; Heutink, P.; Shulman, J.M. International Parkinson’s Disease Genomics Consortium (IPDGC). Excessive burden of lysosomal storage disorder gene variants in Parkinson’s disease. Brain 2017, 140, 3191–3203. [Google Scholar] [CrossRef]
- Parenti, I.; Rabaneda, L.G.; Schoen, H.; Novarino, G. Neurodevelopmental Disorders: From Genetics to Functional Pathways. Trends Neurosci. 2020, 43, 608–621. [Google Scholar] [CrossRef]
- Hummel, T.; Sekinger, B.; Wolf, S.R.; Pauli, E.; Kobal, G. ‘Sniffin’ Sticks’: Olfactory performance assessed by the combined testing of odor identification, odor discrimination and olfactory threshold. Chem. Senses 1997, 22, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.S.; Tajudeen, B.A.; Kennedy, D.W. Diseases of the nasal cavity. Handb. Clin. Neurol. 2019, 164, 285–302. [Google Scholar] [CrossRef] [PubMed]
- Prediger, R.D.; Schamne, M.G.; Sampaio, T.B.; Moreira, E.L.; Rial, D. Animal models of olfactory dysfunction in neurodegenerative diseases. Handb. Clin. Neurol. 2019, 164, 431–452. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: https://www.r-project.org/ (accessed on 7 June 2020).
- Hothorn, T.; Hornik, K.; Zeileis, A. Unbiased Recursive Partitioning: A Conditional Inference Framework. J. Comput. Graph. Stat. 2006, 15, 651–674. [Google Scholar] [CrossRef] [Green Version]
- Strasser, H.; Weber, C. On the asymptotic theory of permutation statistics. Math. Methods Stat. 1999, 8, 220–250. [Google Scholar]
- Thul, P.J.; Lindskog, C. The human protein atlas: A spatial map of the human proteome. Protein Sci. 2018, 27, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Law, M.; Shaw, D.R. Mouse Genome Informatics (MGI) Is the International Resource for Information on the Laboratory Mouse. In Eukaryotic Genomic Databases; Kollmar, M., Ed.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2018; Volume 1757. [Google Scholar]

{kind=link}
{kind=link}
| Gene | Isoform | cDNA Change | Protein Change | Chr | Pos | Ref | Alt | rsID | Freq FVG/VBI | KO FVG/VBI | N Smell FVG/VBI |
|---|---|---|---|---|---|---|---|---|---|---|---|
| OR10J1 | NM_012351.3 | c.759T > A | p.(Cys253*) | 1 | 159410340 | T | A | rs12409540 | 0.0675/0.1144 | 1/6 | 0/1 |
| OR2W3 | NM_001001957.2 | c.893dup | p.(Ala300Glyfs*?) | 1 | 248059779 | G | GA | rs80255919 | 0.1614/0.0873 | 13/2 | 1/0 |
| OR2T4 | NM_001004696.1 | c.757del | p.(Ile253Serfs*8) | 1 | 248525638 | CA | C | rs34079073 | 0.5106/0.5224 | 110/120 | 40/20 |
| OR5K4 | NM_001005517.1 | c.901del | p.(Ile301Leufs*2) | 3 | 98073591 | TA | T | rs11288615 | 0.6177/0.5224 | 141/121 | 57/31 |
| OR5K3 | NM_001005516.1 | c.904dup | p.(Ile302Asnfs*?) | 3 | 98110406 | G | GA | rs79045298 | 0.6098/0.5377 | 138/129 | 55/33 |
| OR5K2 | NM_001004737.1 | c.654T > A | p.(Tyr218*) | 3 | 98217178 | T | A | rs55639376 | 0.1204/0.1392 | 3/11 | 2/1 |
| OR2V2 | NM_206880.1 | c.320_323del | p.(Cys107Leufs*30) | 5 | 180582256 | TTGTC | T | rs140598308 | 0.0582/0.1014 | 2/3 | 0/0 |
| OR13C5 | NM_001004482.1 | c.926del | p.(His309Profs*3) | 9 | 107360768 | GT | G | rs11314210 | 0.1706/0.1321 | 13/3 | 5/1 |
| OR1J1 | NM_001004451.1 | c.705C > A | p.(Cys235*) | 9 | 125239501 | G | T | rs45579335 | 0.0066/0.0224 | 0/1 | 0/0 |
| OR1J2 | NM_054107.1 | c.312dup | p.(Ile105Tyrfs*5) | 9 | 125273385 | A | AT | rs145911830 | 0.1204/0.1439 | 3/8 | 2/2 |
| OR13A1 | NM_001004297.3 | c.805dup | p.(Tyr269Leufs*66) | 10 | 45799065 | T | TA | rs35302355 | 0.0608/0.0377 | 5/1 | 3/1 |
| OR51T1 | NM_001004759.2 | c.551_552insCACCACCC | p.(Glu185Thrfs*5) | 11 | 4903673 | T | TACCACCCC | rs564566592 | 0.004/0.013 | 0/1 | 0/0 |
| OR52J3 | NM_001001916.2 | c.907C > T | p.(Arg303*) | 11 | 5068662 | C | T | rs57026471 | 0.1098/0.1285 | 5/7 | 0/1 |
| OR52E2 | NM_001005164.2 | c.551del | p.(Met184Argfs*25) | 11 | 5080307 | AT | A | - | (null)/0.0106 | (null)/1 | 0/0 |
| OR52A1 | NM_012375.2 | c.804dup | p.(Ser269Valfs*13) | 11 | 5172795 | A | AC | rs112098990 | 0.2447/0.3113 | 27/44 | 6/17 |
| OR51B5 | NM_001005567.3 | c.197_213del | p.(Ala66Glyfs*48) | 11 | 5364541 | CCAGCCCCAGGTCTGTGG | C | rs147062602 | (null)/0.0377 | (null)/1 | 0/0 |
| OR51J1 | NM_001348224.1 | c.567_570dup | p.(Cys191Ilefs*8) | 11 | 5424387 | T | TTATC | rs113047337 | 0.1005/0.092 | 7/6 | 1/3 |
| OR51Q1 | NM_001004757.2 | c.706C > T | p.(Arg236*) | 11 | 5444136 | C | T | rs2647574 | 0.3532/0.4021 | 55/79 | 20/19 |
| OR51I1 | NM_001005288.2 | c.43C > T | p.(Gln15*) | 11 | 5462702 | G | A | rs16930998 | 0.0172/0.0212 | 1/1 | 1/0 |
| OR51I2 | NM_001004754.2 | c.714_715dup | p.(Asn239Thrfs*18) | 11 | 5475431 | T | TCA | rs35301588 | 0.3704/0.309 | 54/46 | 15/12 |
| OR52D1 | NM_001005163.2 | c.605_608dup | p.(Thr204Alafs*33) | 11 | 5510540 | G | GGGCT | rs576495879 | 0.1442/0.1568 | 17/11 | 5/5 |
| OR52N4 | NM_001005175.3 | c.514A > T | p.(Arg172*) | 11 | 5776484 | A | T | rs4910844 | 0.2341/0.3208 | 23/42 | 8/9 |
| OR4X1 | NM_001004726.1 | c.819T > A | p.(Tyr273*) | 11 | 48286231 | T | A | rs10838851 | 0.7659/0.6344 | 223/165 | 76/38 |
| OR4C11 | NM_001004700.2 | c.469C > T | p.(Gln157*) | 11 | 55371381 | G | A | rs75423534 | 0.0754/0.0955 | 14/18 | 7/5 |
| OR4P4 | NM_001004124.2 | c.189C > G | p.(Tyr63*) | 11 | 55406022 | C | G | rs76160133 | 0.1296/0.1922 | 21/45 | 7/4 |
| OR8I2 | NM_001003750.1 | c.867C > G | p.(Tyr289*) | 11 | 55861650 | C | G | rs61887097 | 0.1124/0.0849 | 7/5 | 3/1 |
| OR5M11 | NM_001005245.1 | c.378T > A | p.(Tyr126*) | 11 | 56310356 | A | T | rs17547284 | 0.119/0.0896 | 10/4 | 4/1 |
| OR5M10 | NM_001004741.1 | c.347_354del | p.(Ala116Glyfs*37) | 11 | 56344843 | CCATTGAAG | C | rs148438199 | 0.119/0.0873 | 10/4 | 4/1 |
| OR5M1 | NM_001004740.1 | c.429_432del | p.(Cys143Trpfs*19) | 11 | 56380546 | CCAGA | C | rs71931749 | 0.2302/0.263 | 21/24 | 13/7 |
| OR6Q1 | NM_001005186.2 | c.685del | p.(Leu229Cysfs*21) | 11 | 57799108 | AC | A | rs34846253 | 0.2487/0.2229 | 25/20 | 8/5 |
| OR10D3 | NM_001355213.1 | c.756T > G | p.(Tyr252*) | 11 | 124056732 | T | G | rs2512227 | 0.4987/0.5696 | 90/135 | 27/33 |
| OR8B3 | NM_001005467.1 | c.550dup | p.(Leu184Profs*23) | 11 | 124266697 | A | AG | rs201661436 | 0.0635/0.0649 | 0/3 | 0/2 |
| OR10AD1 | NM_001004134.1 | c.199_200insG | p.(Leu67Argfs*56) | 12 | 48596875 | C | CA | rs79650217 | 0.2063/0.2406 | 19/28 | 6/8 |
| OR9K2 | NM_001005243.1 | c.38del | p.(Leu13Cysfs*22) | 12 | 55523586 | AT | A | rs58036029 | 0.3823/0.3278 | 59/43 | 21/13 |
| OR6C74 | NM_001005490.1 | c.184C > T | p.(Arg62*) | 12 | 55641255 | C | T | rs4522268 | 0.377/0.3243 | 56/42 | 21/13 |
| OR6C1 | NM_001005182.1 | c.24dup | p.(Glu9Argfs*10) | 12 | 55714406 | C | CA | rs5798345 | 0.4405/0.3892 | 79/64 | 26/10 |
| OR6C76 | NM_001005183.1 | c.933del | p.(Lys311Asnfs*?) | 12 | 55820958 | CA | C | rs57387180 | 0.1772/0.2205 | 12/26 | 6/6 |
| OR4L1 | NM_001004717.1 | c.248_266del | p.(Ile83Thrfs*10) | 14 | 20528448 | TCATAGATTTGCTCACTGAC | T | rs33965693 | 0.3981/0.3561 | 64/54 | 25/13 |
| OR11G2 | NM_001005503.1 | c.687_688dup | p.(Gly230Lysfs*4) | 14 | 20666175 | C | CA | rs55781225 | 0.668/0.6568 | 170/183 | 71/32 |
| OR2C1 | NM_012368.3 | c.818del | p.(Phe273Serfs*13) | 16 | 3406756 | GT | G | rs142397376 | 0.0886/0.0837 | 1/1 | 0/0 |
| OR7G3 | NM_001001958.1 | c.928_929insACTAT | p.(Ser310Tyrfs*?) | 19 | 9236698 | G | GATGGT | rs111867493 | 0.2791/0.2925 | 27/40 | 10/11 |
| OR7G3 | NM_001001958.1 | c.710del | p.(Ala237Valfs*9) | 19 | 9236916 | AG | A | rs75266995 | 0.0304/0.0519 | 0/3 | 0/0 |
| N (males %) | 218 (43.6%) |
| Age (y), mean (SD) | 61.9 (15.3) |
| Number of errors in Sniffin’ Sticks test, median (IQR) | 2.0 (1.0–4.0) |
| Normosmic *, % | 34.9 |
| Hyposmic, % | 50.9 |
| Anosmic, % | 14.2 |
| Number of individuals for each OR-KO gene **, [range], median (IQR) | [0–114], 8.0 (1.0–28.75) |
| Number of OR-KO genes per individual, [range], median (IQR) | [0–11], 4.0 (3.0–5.0) |
| Human Gene Symbol | Expression Human OE (Saraiva et al., 2019) | Expression Human Tissues (Human Protein Atlas) | Mouse Gene Symbol (Mouse Genome Informatics) | Expression Mouse OE (Saraiva et al., 2019) | Expressed Mouse OS (Mouse Genome Informatics) | Expression Mouse Tissues (Mouse Genome Informatics) |
|---|---|---|---|---|---|---|
| OR10J1 | 4.37 | Testis, granulocytes | Olfr418 (1) | NA | Yes | Alimentary system |
| OR2W3 | 4.7 | Bone marrow, thyroid gland, cerebral cortex, hypothalamus, basal ganglia | Olfr322 | NA | Yes | NA |
| Olfr317 | 24.29 | Yes | Nervous system, reproductive system | |||
| OR2T4 | 2.22 | Prostate, cervix uterine, cerebral cortex | Olfr331 | 0.28 | No | NA |
| Olfr224 | 303.89 | Yes | Hemolymphoid system, reproductive system | |||
| Olfr325 | 245.05 | Yes | Embryo ectoderm, auditory system, reproductive system | |||
| Olfr328 | 287.48 | Yes | Reproductive system | |||
| Olfr329 (2) | 395.85 | Yes | Early conceptus, endocrine system, hemolymphoid system, reproductive system | |||
| Olfr330 | 341.8 | Yes | Alimentary system, auditory system, endocrine system, reproductive system | |||
| OR5K4 | 0 | 0 | Olfr180 | 188.79 | Yes | Reproductive system |
| OR5K3 | 0 | 0 | Olfr175 (3) | NA | Yes | Liver and biliary system |
| Olfr195 | 506.03 | Yes | Urinary system | |||
| OR5K2 | 7.52 | Skeletal muscle, cerebellum, skin, lung, colon | Olfr177 | 142.22 | Yes | NA |
| OR2V2 | 38.24 | Granulocytes, bone marrow, fallopian tube | Olfr1396 | 203.03 | Yes | Cardiovascular system, connective tissue, hemolymphoid system, integumental system, limbs, liver and biliary system, musculoskeletal system, urinary system |
| OR13C5 | 2.07 | 0 | Olfr452 | 78.19 | Yes | Auditory system |
| OR1J1 | 2.9 | Salivary gland, testis, bone marrow, granulocytes | Olfr3 | 34.88 | Yes | Auditory system, reproductive system |
| OR1J2 | 4.89 | Urinary bladder, epididymis, testis | Olfr348 | 22.23 | Yes | NA |
| OR13A1 | 43.39 | Urinary bladder, spleen, lymph node, tonsil, B-cells | Olfr211 | 404.96 | Yes | NA |
| OR51T1 | 1.04 | Prostate | Olfr574 | 0 | Yes | Endocrine system, nervous system |
| OR52J3 | 1.66 | 0 | Olfr592 | 36.36 | Yes | Auditory system |
| OR52E2 | 0.71 | Testis | Olfr589 | 1.33 | Yes | Auditory system |
| Olfr594 | 63.59 | Yes | Nervous system | |||
| OR52A1 | 50.01 | Granulocytes, testis, B-cells, skeletal muscle, cerebellum | Olfr68 | 18.3 | Yes | Liver and biliary system |
| OR51B5 | 1.71 | Epididymis, T-cells | NA | NA | NA | NA |
| OR51J1 | 0.71 | NA | NA | NA | NA | NA |
| OR51Q1 | 0.94 | Epididymis, cerebellum | Olfr635 | 36.72 | Yes | NA |
| Olfr638 | 11.5 | Yes | NA | |||
| OR51I1 | 3.73 | Epididymis, testis | Olfr639 | 46.82 | Yes | Reproductive system |
| Olfr640 | 136.96 | Yes | NA | |||
| OR51I2 | 0 | Granulocytes | Olfr641 | 15.13 | Yes | Branchial arches, nervous system |
| OR52D1 | 1.04 | Testis | Olfr646 | 25 | Yes | NA |
| Olfr691 | 79.3 | Yes | Auditory system, nervous system, reproductive system | |||
| OR52N4 | 11.26 | Spleen, small intestine, ovary, epididymis, T-Cells | Olfr503 | 0.66 | NA | NA |
| Olfr658 | 14.75 | Yes | Nervous system, visual system | |||
| OR4X1 | 0 | 0 | NA | NA | NA | NA |
| OR4C11 | 3.39 | 0 | Olfr1201 | 27.04 | Yes | NA |
| Olfr1205 | 158.61 | Yes | NA | |||
| Olfr1206 | 206.79 | Yes | NA | |||
| OR4P4 | 8.26 | Bone marrow, granulocytes, skin, natural killer (NK) cells | NA | NA | NA | NA |
| OR8I2 | 0 | 0 | Olfr1104 | 81.01 | Yes | Early conceptus, reproductive system |
| OR5M11 | 0 | Urinary bladder, testis | Olfr1028 | 23.72 | Yes | NA |
| Olfr1029 | 18.05 | Yes | Liver and biliary system, nervous system, reproductive system | |||
| OR5M10 | 17.86 | Salivary gland | Olfr1022 | 2.42 | Yes | Nervous system |
| Olfr1023 | 12.88 | Yes | Nervous system | |||
| OR5M1 | 16.73 | 0 | Olfr1023 | 12.88 | Yes | Nervous system |
| OR6Q1 | 0 | 0 | NA | NA | NA | NA |
| OR10D3 | 7.09 | Testis | Olfr958 | 42.05 | Yes | Branchial arches, nervous system |
| OR8B3 | 22.86 | Testis | Olfr147 | 52.98 | Yes | Early conceptus |
| OR10AD1 | 2.28 | Pituitary gland, adrenal gland, testis, cerebellum, appendix | Olfr286 | NA | Yes | Embryo ectoderm, hemolymphoid system, nervous system, reproductive system |
| Olfr287 | NA | Yes | Early conceptus, hemolymphoid system, nervous system, reproductive system | |||
| Olfr288 | 80.43 | Yes | Early conceptus, alimentary system, musculoskeletal system, reproductive system, urinary system | |||
| OR9K2 | 9.33 | 0 | Olfr825 | 41.63 | Yes | NA |
| Olfr826 | 32.78 | Yes | NA | |||
| OR6C74 | 0.35 | 0 | Olfr821 | 95.42 | Yes | NA |
| OR6C1 | 21.11 | 0 | Olfr786 | 119.92 | Yes | NA |
| Olfr802 | 30.95 | Yes | NA | |||
| OR6C76 | 5.05 | Epididymis, fallopian tube | Olfr792 | 23.76 | Yes | Nervous system, reproductive system |
| Olfr798 | 60.66 | Yes | Embryo ectoderm | |||
| Olfr809 | 41.56 | Yes | NA | |||
| Olfr813 | 56.92 | Yes | Auditory system | |||
| OR4L1 | 0.88 | 0 | Olfr723 | 91.15 | Yes | NA |
| Olfr724 | 33.25 | Yes | NA | |||
| OR11G2 | 160.36 | Bone marrow | Olfr744 | 39.01 | Yes | Hemolymphoid system |
| OR2C1 | 1.65 | Fallopian tube, T-cells, spinal cord, parathyroid gland, B-cells | Olfr15 | 785.44 | Yes | Auditory system |
| OR7G3 | 0 | Fallopian tube | Olfr832 | 9.79 | Yes | Reproductive system |
| Olfr834 | 0 | Yes | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Concas, M.P.; Cocca, M.; Francescatto, M.; Battistuzzi, T.; Spedicati, B.; Feresin, A.; Morgan, A.; Gasparini, P.; Girotto, G. The Role of Knockout Olfactory Receptor Genes in Odor Discrimination. Genes 2021, 12, 631. https://doi.org/10.3390/genes12050631
Concas MP, Cocca M, Francescatto M, Battistuzzi T, Spedicati B, Feresin A, Morgan A, Gasparini P, Girotto G. The Role of Knockout Olfactory Receptor Genes in Odor Discrimination. Genes. 2021; 12(5):631. https://doi.org/10.3390/genes12050631
Chicago/Turabian StyleConcas, Maria Pina, Massimiliano Cocca, Margherita Francescatto, Thomas Battistuzzi, Beatrice Spedicati, Agnese Feresin, Anna Morgan, Paolo Gasparini, and Giorgia Girotto. 2021. "The Role of Knockout Olfactory Receptor Genes in Odor Discrimination" Genes 12, no. 5: 631. https://doi.org/10.3390/genes12050631
APA StyleConcas, M. P., Cocca, M., Francescatto, M., Battistuzzi, T., Spedicati, B., Feresin, A., Morgan, A., Gasparini, P., & Girotto, G. (2021). The Role of Knockout Olfactory Receptor Genes in Odor Discrimination. Genes, 12(5), 631. https://doi.org/10.3390/genes12050631