
Assessment of the Genetic Potential of the Peregrine Falcon (Falco peregrinus peregrinus) Population Used in the Reintroduction Program in Poland


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and DNA Extraction
2.2. Microsatellite Genotyping
2.3. Statistical Analyses
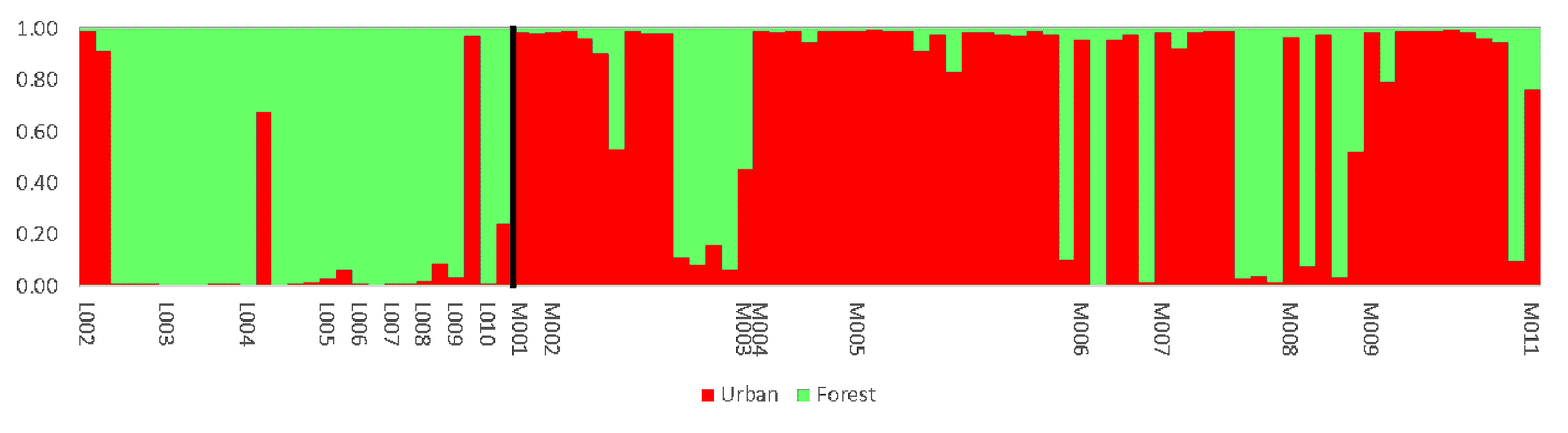
3. Result
4. Discussions
4.1. Population Variability
4.2. Bird Release Point
4.3. Stock of Falcons for Reintroduction
4.4. Implications for Conservation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bryndová, M.; Putnová, L.; Bartoňová, P.; Kaplanová, K.; Urban, T. Genetic differences between wild and captive populations of the peregrine falcon (Falco peregrinus) and the saker falcon (Falco cherrung) living in the Czech Republic. J. Agric. Sci. Technol. B 2012, 2, 642–651. [Google Scholar]
- Wegner, P.; Kleinstäuber, G.; Baum, F.; Schilling, F. Long-term investigation of the degree of exposure of German peregrine falcons (Falco peregrinus) to damaging chemicals from the environment. J. Ornithol. 2005, 146, 34–54. [Google Scholar] [CrossRef]
- Brown, J.W.; Van Coeverden De Groot, P.J.; Birt, T.P.; Seutin, G.; Boag, P.T.; Friesen, V.L. Appraisal of the consequences of the DDT-induced bottleneck on the level and geographic distribution of neutral genetic variation in Canadian peregrine falcons, Falco peregrinus. Mol. Ecol. 2007, 16, 327–343. [Google Scholar] [CrossRef]
- Crick, H.Q.P.; Ratcliffe, D.A. The peregrine Falco peregrinus breeding population of the United Kingdom in 1991. Bird Study 1995, 42, 1–19. [Google Scholar] [CrossRef]
- Olsen, P.; Emison, B.; Mooney, N.; Brothers, N. DDT and dieldrin: Effects on resident peregrine falcon populations in south-eastern Australia. Ecotoxicology 1992, 1, 89–100. [Google Scholar] [CrossRef]
- Mizera, T.; Sielicki, J. The peregrine falcon Falco peregrinus in Poland—Its situation and perspectives for reinstatment. Acta Ornithol. 1995, 30, 47–52. [Google Scholar]
- Sielicki, S.; Sielicki, J. Restoration of peregrine falcon in Poland 1989–2007. In Peregrine Falcon Populations—Status and Perspectives in 21th Century, 1st ed.; Sielicki, J., Mizera, B., Eds.; Turul publishing and Poznań University of Life Sciences Press: Warsaw-Poznań, Poland, 2009; Volume 1, pp. 699–722. [Google Scholar]
- Cichocki, W. Some species of breeding birds in Tatra National Park. Parki Narodowe i Rezerwaty Przyrody 1986, 7, 57–62. (In Polish) [Google Scholar]
- Ivanovsky, V.; Vintchevski, A. Status of the peregrine falcon in Belarus. In Peregrine Falcon Populations—Status and Perspectives in 21th Century, 1st ed.; Sielicki, J., Mizera, B., Eds.; Turul publishing and Poznań University of Life Sciences Press: Warsaw-Poznań, Poland, 2009; Volume 1, pp. 117–120. [Google Scholar]
- Brambilla, M.; Rubolini, D.; Guidali, F. Factors affecting breeding habitat selection in a cliff-nesting peregrine Falco peregrinus population. J. Ornithol. 2006, 147, 428–435. [Google Scholar] [CrossRef]
- Kirmse, W. Tree-nesting Peregrines Falco p. peregrinus in Europe did not recover. In Raptors Worldwide, 1st ed.; Chancellor, R.D., Meyburg, B.U., Eds.; WWGBP: Berlin, Germany, 2004; Volume 1, pp. 271–277. [Google Scholar]
- Sielicki, S.; Sielicki, J. Falconry and the restoration of the peregrine falcon in Poland in 1990–2015. In Falconary—Its Influence on Biodiversity and Cultural Heritage, 1st ed.; Szymak, U., Sianko, P., Eds.; Muzeum Podlaskie w Białymstoku: Białystok, Poland, 2016; Volume 1, pp. 41–47. [Google Scholar]
- Ławicki, Ł.; Sielicki, S. Restoration of the tree-nesting population of the peregrine falcon Falco peregrinus in Pomerania. Ornis Pol. 2019, 60, 75–84. [Google Scholar]
- Banks, A.N.; Crick, H.Q.P.; Coombes, R.; Benn, S.; Ratcliffe, D.A.; Humphreys, E.M. The breeding status of peregrine falcons Falco peregrinus in the UK and Isle of Man in 2002. Bird Study 2010, 57. [Google Scholar] [CrossRef] [Green Version]
- Verdejo, J.; López-López, P. Long-term monitoring of a peregrine falcon population: Size, breeding performance and nest-site characteristics. Ardeola 2008, 55, 87–96. [Google Scholar]
- Wisniewski, G. Programme for the reinstatement of the peregrine falcon Falco peregrinus in Poland. Acta Ornithol. 1995, 30, 73–78. [Google Scholar]
- Nesje, M.; Røed, K.H.; Bell, D.A.; Lindberg, P.; Lifjeld, J.T. Microsatellite analysis of population structure and genetic variability in peregrine falcons (Falco peregrinus). Anim. Conserv. 2000, 3, 267–275. [Google Scholar] [CrossRef]
- Nittinger, F.; Gamauf, A.; Pinsker, W.; Wink, M.; Haring, E. Phylogeography and population structure of the saker falcon (Falco cherrug) and the influence of hybridization: Mitochondrial and microsatellite data. Mol. Ecol. 2007, 16, 1497–1517. [Google Scholar] [CrossRef]
- Jacobsen, F.; Nesje, M.; Bachmann, L.; Lifjeld, J.T. Significant genetic admixture after reintroduction of peregrine falcon (Falco peregrinus) in Southern Scandinavia. Conserv. Genet. 2008, 9, 581–591. [Google Scholar] [CrossRef]
- Bell, D.A.; Griffiths, C.S.; Caballero, I.C.; Hartley, R.R.; Lawson, R.H. Genetic Evidence for Global Dispersal in the peregrine falcon (Falco peregrinus) and Affinity with the taita falcon (Falco fasciinucha). J. Raptor Res. 2014, 48, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Rutkowski, R.; Rejt, Ł.; Tereba, A.; Gryczyńska-Siemiatkowska, A.; Janic, B. Population genetic structure of the European kestrel Falco tinnunculus in Central Poland. Eur. J. Wildl. Res. 2010, 56, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Groombridge, J.J.; Dawson, D.A.; Burke, T.; Prys-Jones, R.; Brooke, M.; de L. Brooke, M.; Shah, N. Evaluating the demographic history of the Seychelles kestrel (Falco araea): Genetic evidence for recovery from a population bottleneck following minimal conservation management. Biol. Conserv. 2009, 142, 2250–2257. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. GenAlEx 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Marshall, T.C.; Slate, J.; Kruuk, L.; Pemberton, J.M. Statistical confidence for likelihood-based paternity inference in natural populations. Mol. Ecol. 1998, 7, 639–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Sneath, P.H.A.; Sokal, R.R. Numerical Taxonomy; Freeman: San Francisco, CA, USA, 1973. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked Loci and Correlated Allele Frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Talbot, S.L.; Palmer, A.G.; Sage, G.K.; Sonsthagen, S.A.; Swem, T.; Brimm, D.J.; White, C.M. Lack of genetic polymorphism among peregrine falcons Falco peregrinus of Fiji. J. Avian Biol. 2011, 42, 415–428. [Google Scholar] [CrossRef]
- White, C.M.; Cade, T.J.; Enderson, J.H. Peregrine Falcons of the World; Lynx Edicions: Barcelona, Spain, 2013. [Google Scholar]
- Minias, P.; Włodarczyk, R.; Minias, A.; Dziadek, J. How birds colonize cities: Genetic evidence from a common waterbird, the Eurasian coot. J. Avian Biol. 2017, 48, 1095–1103. [Google Scholar] [CrossRef]
- Johnson, J.A.; Talbot, S.L.; Sage, G.K.; Burnham, K.K.; Brown, J.W.; Maechtle, T.L.; Seegar, W.S.; Yates, M.A.; Anderson, B.; Mindell, D.P. The use of genetics for the management of a recovering population: Temporal assessment of migratory peregrine falcons in North America. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilcox, J.J.S.; Boissinot, S.; Idaghdour, Y. Falcon genomics in the context of conservation, speciation, and human culture. Ecol. Evol. 2019, 9, 14523–14537. [Google Scholar] [CrossRef]
- Fleming, L.V.; Douse, A.F.; Williams, N.P. Captive breeding of peregrine and other falcons in Great Britain and implications for conservation of wild populations. Endanger. Species Res. 2011, 14, 243–257. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Allele Ranges | n | Na | Ne | Ho | He | FST |
|---|---|---|---|---|---|---|---|
| NVHfp107 | 114–208 | 350 | 6 | 1.950 | 0.454 | 0.487 | 0.042 |
| NVHfp13 | 93–103 | 350 | 9 | 4.714 | 0.554 | 0.788 | 0.030 |
| NVHfp46_1 | 117–122 | 350 | 6 | 2.925 | 0.540 | 0.658 | 0.055 |
| NVHfp5 | 104–108 | 352 | 3 | 1.489 | 0.057 | 0.328 | 0.159 |
| NVHfp54 | 104–116 | 348 | 7 | 2.395 | 0.382 | 0.528 | 0.049 |
| NVHfp79_4 | 104–142 | 352 | 3 | 1.982 | 0.000 | 0.495 | 0.029 |
| NVHfp82_2 | 134–140 | 352 | 4 | 1.143 | 0.085 | 0.125 | 0.060 |
| NVHfp86_2 | 140–145 | 343 | 5 | 2.146 | 0.370 | 0.534 | 0.045 |
| NVHfp89 | 116–132 | 347 | 9 | 4.467 | 0.617 | 0.776 | 0.052 |
| NVHfp92_1 | 110–130 | 349 | 9 | 2.617 | 0.298 | 0.618 | 0.075 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puchała, K.O.; Nowak-Życzyńska, Z.; Sielicki, S.; Olech, W. Assessment of the Genetic Potential of the Peregrine Falcon (Falco peregrinus peregrinus) Population Used in the Reintroduction Program in Poland. Genes 2021, 12, 666. https://doi.org/10.3390/genes12050666
Puchała KO, Nowak-Życzyńska Z, Sielicki S, Olech W. Assessment of the Genetic Potential of the Peregrine Falcon (Falco peregrinus peregrinus) Population Used in the Reintroduction Program in Poland. Genes. 2021; 12(5):666. https://doi.org/10.3390/genes12050666
Chicago/Turabian StylePuchała, Karol O., Zuzanna Nowak-Życzyńska, Sławomir Sielicki, and Wanda Olech. 2021. "Assessment of the Genetic Potential of the Peregrine Falcon (Falco peregrinus peregrinus) Population Used in the Reintroduction Program in Poland" Genes 12, no. 5: 666. https://doi.org/10.3390/genes12050666