
Comprehensive Transcriptome Analysis of Rare Carpinus putoensis Plants under NO2 stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. NO2 Fumigation
2.3. Determination of Total Peroxidase (POD) Activity
2.4. Transmission Electron Microscopy (TEM)
2.5. RNA Isolation, cDNA Library Construction, and Illumina Sequencing
2.6. Data Analysis for RNA-seq Experiments
2.7. Identification, Annotation, and Enrichment Analysis of Differentially Expressed Genes
2.8. Validation by RT-qPCR
3. Results
3.1. Morphology and Cell Structure of C. putoensis Leaves
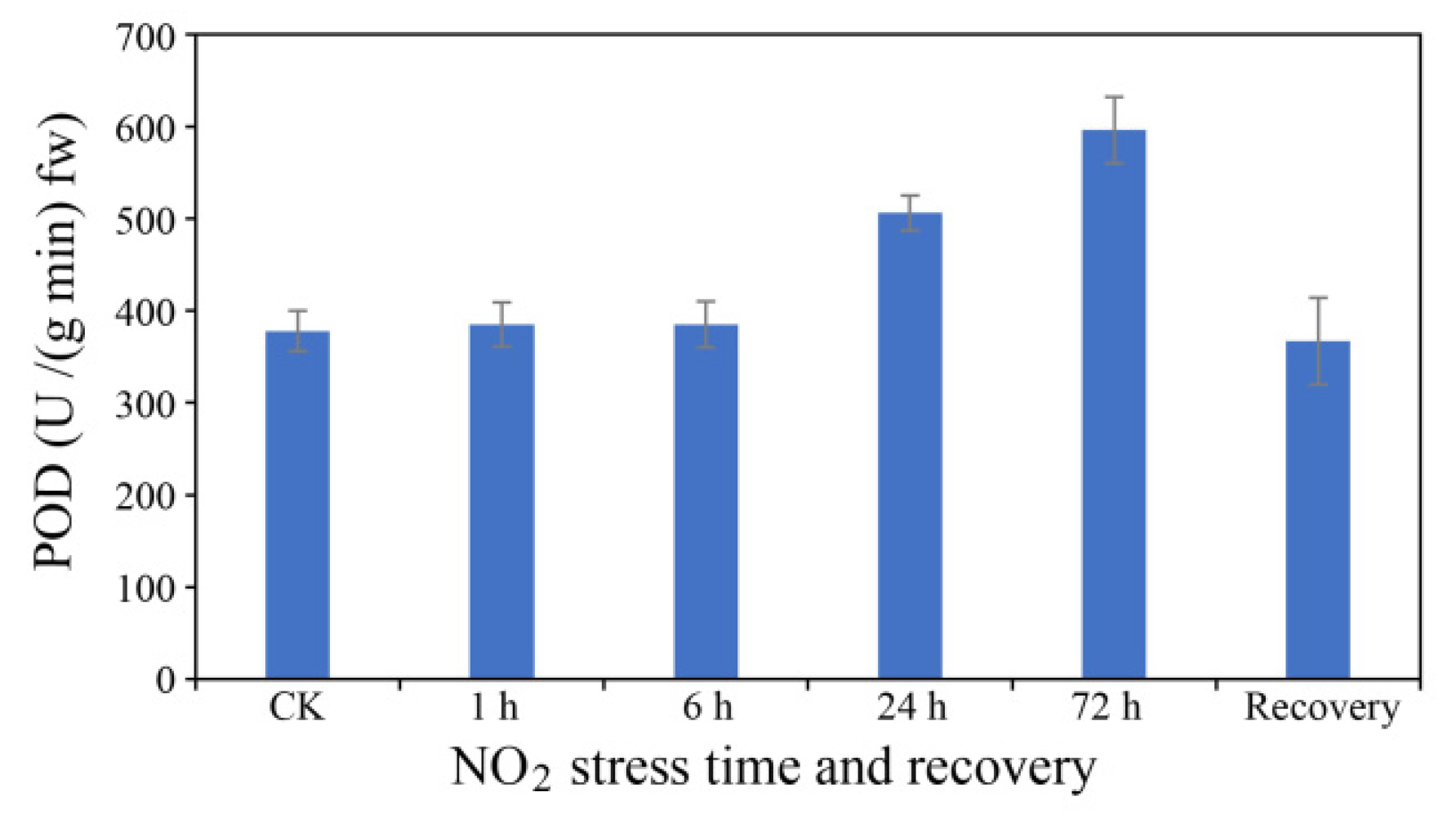
3.2. Changes in POD Activity
3.3. RNA-seq Analysis of Clean Data from C. putoensis
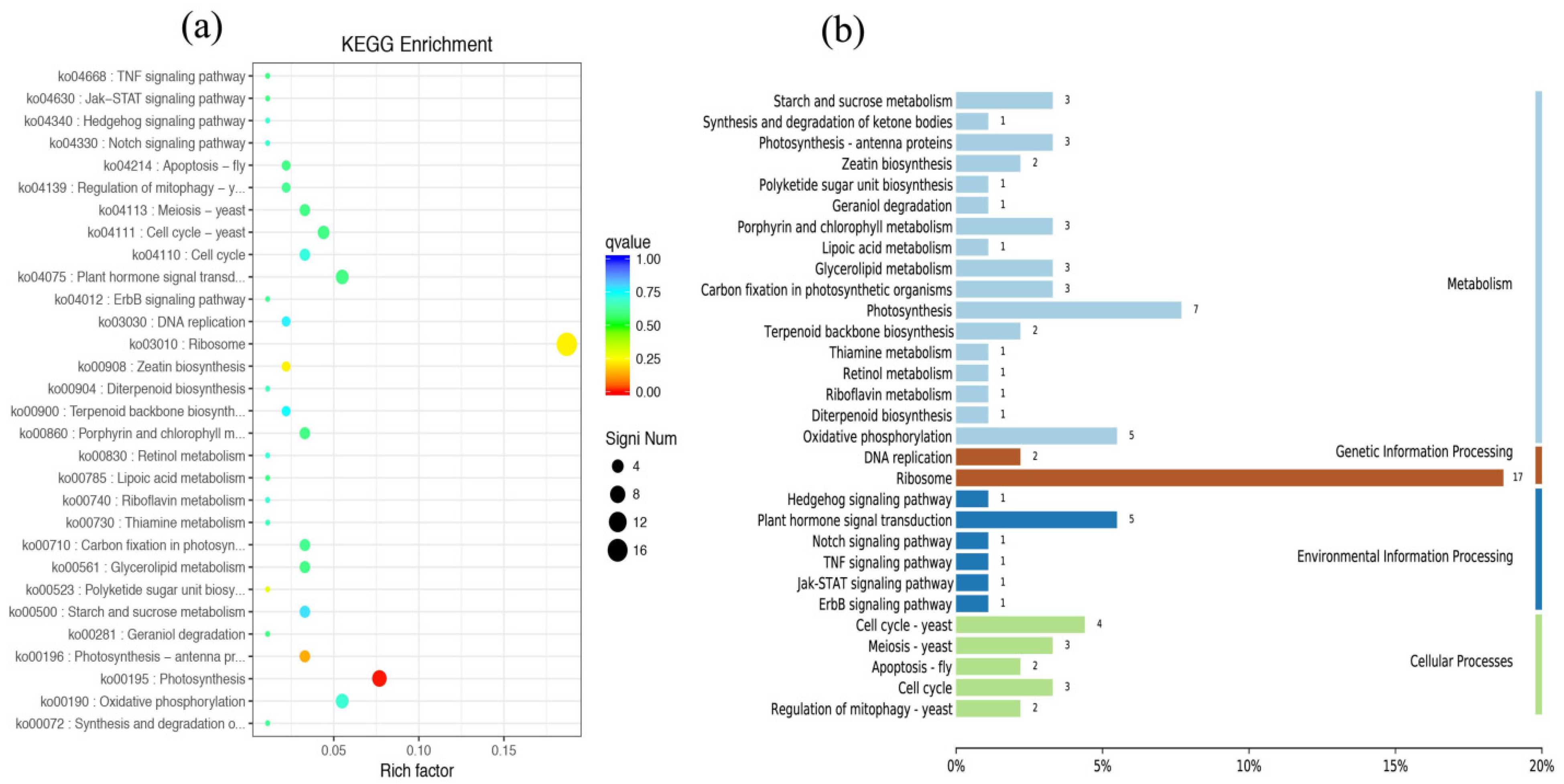
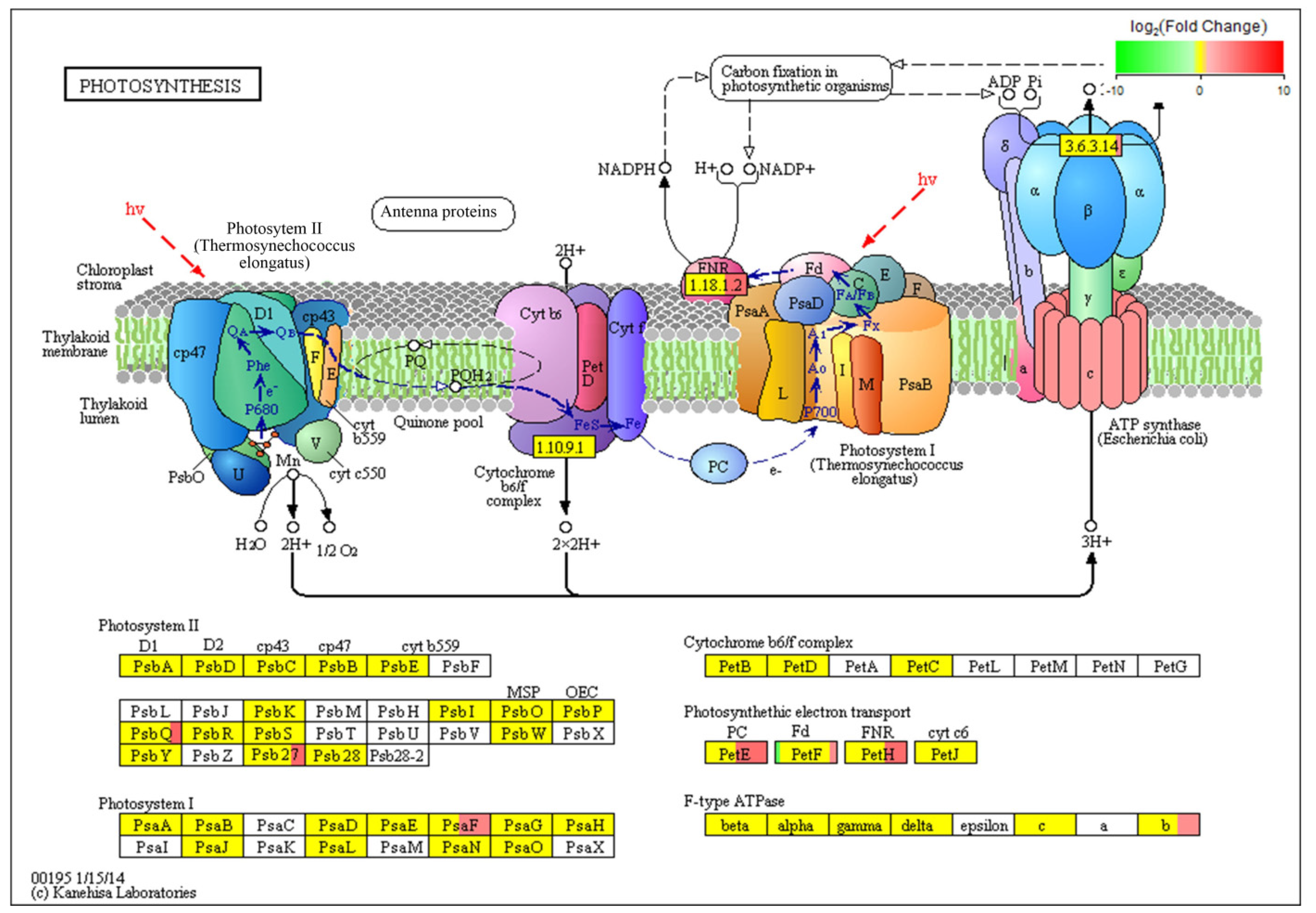
3.4. Identification and Analysis of DEGs in C. putoensis Leaves under NO2 Stress
3.5. RT-qPCR Analysis of NO2 Stress-related Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rahmat, M.; Maulina, W.; Rustami, E.; Azis, M.; Budiarti, D.; Seminar, K.; Yuwono, A.; Alatas, H. Performance in real condition of photonic crystal sensor based NO2 gas monitoring system. Atmos. Environ. 2013, 79, 480–485. [Google Scholar] [CrossRef]
- Sasakawa, H.; Yoneyama, T. Transformation of atmospheric NO2 absorbed in spinach leaves. Plant Cell Physiol. 1979, 20, 263–266. [Google Scholar] [CrossRef]
- Bermejo-Orduna, R.; McBride, J.; Shiraishi, K.; Elustondo, D.; Lasheras, E.; Santamaría, J. Biomonitoring of traffic-related nitrogen pollution using Letharia vulpina (L.) Hue in the Sierra Nevada, California. Sci. Total Environ. 2014, 490, 205–212. [Google Scholar] [CrossRef]
- Lu, M.; Li, Y.J.; Lu, J.P. The study of greening trees on the atmospheric pollutant absorption ability. J. Urban. Environ. Urban. Ecol. 2002, 15, 7–9. [Google Scholar]
- Stulen, I.; Pérez-Soba, M.; de Kok, L.J.; van der Eerden, L. Impact of gaseous nitrogen deposition on plant functioning. New Phytol. 1998, 139, 61–70. [Google Scholar] [CrossRef]
- Morikawa, H.; Takahashi, M.; Sakamoto, A.; Matsubara, T.; Arimura, G.-I.; Kawamura, Y.; Fukunaga, K.; Fujita, K.; Sakurai, N.; Hirata, T.; et al. Formation of unidentified nitrogen in plants: An implication for a novel nitrogen metabolism. Planta 2004, 219, 14–22. [Google Scholar] [CrossRef]
- Nussbaum, S.; von Ballmoos, P.; Gfeller, H.; Schlunegger, U.P.; Fuhrer, J.; Rhodes, D.; Brunold, C.; Ballmoos, P. Incorporation of atmospheric 15NO2-nitrogen into free amino acids by Norway spruce Picea abies (L.) Karst. Oecologia 1993, 94, 408–414. [Google Scholar] [CrossRef]
- Weber, P.; Nussbaum, S.; Fuhrer, J.; Gfeller, H.; Schlunegger, U.P.; Brunold, C.; Rennenberg, H. Uptake of atmospheric 15NO2 and its incorporation into free amino acids in wheat (Triticum aestivum L.). Physiol. Plant. 2006, 94, 71–77. [Google Scholar] [CrossRef]
- Beneoict, H.M.; Breen, W.H. The use of weeds as a means of evaluating vegetation damage caused by air pollution. Proc. Nat. Ah. Polhtt. Syrup 1995, 59, 177–190. [Google Scholar]
- Middleton, J.T.; Darley, E.F.; Brewer, R.F. Damage to Vegetation from Polluted Atmospheres. J. Air Pollut. Control. Assoc. 1958, 8, 9–15. [Google Scholar] [CrossRef]
- Mayer, D.; Mithofer, A. Short-term exposure to nitrogen dioxide provides Bbasal pathogen resistance. Plant. Physiol. 2018, 178, 468–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morikawa, H.; Higaki, A.; Nohno, M.; Takahashi, M.; Kamada, M.; Nakata, M.; Toyohara, G.; Okamura, Y.; Matsui, K.; Kitani, S.; et al. More than a 600-fold variation in nitrogen dioxide assimilation among 217 plant taxa. Plant. Cell Environ. 1998, 21, 180–190. [Google Scholar] [CrossRef]
- Yang, J.; Caia, L.; Liua, D.; Chen, G.; Gratzfeld, J. China’s conservation program on Plant Species with Extremely Small Pop-ulations (PSESP): Progress and perspectives. Biol. Conserv. 2020, 244, 108535. [Google Scholar] [CrossRef]
- Ma, Q.F. The Adjustment of Endogenous Salicylic Acid on NO2 Stress in Arabidopsis; Shenyang Normal University: Shenyang, China, 2016. [Google Scholar]
- Zhong, T.L.; Li, G.Y.; Shi, B.L. Comparison of gas exchange and chlorophyll fluorescence parameters in three endangered species of Zhejiang Province. J. Shanghai Jiaotong Univ. Agric. Sci. 2009, 27, 149–152. [Google Scholar]
- Sheng, Q.Q.; Zhu, Z.L. Photosynthetic Capacity, Stomatal Behavior and Chloroplast Ultrastructure in Leaves of the Endan-gered Plant Carpinus putoensis W.C.Cheng during Gaseous NO2 Exposure and after Recovery. Forests 2018, 9, 561. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.G.; Jensen, M.E.; Wright, J.L.; Burman, R.D. Operational estimates of evapotranspiration. Agron. J. 1989, 81, 650–662. [Google Scholar] [CrossRef]
- Asada, K. Production and Action Oxygen in Photosynthetic Tissue; CRC Press: Boca Raton FL, USA, 1994; pp. 77–104. [Google Scholar]
- Wu, Y.X.; Andreas, V.T. Impact of fungicides on active oxygen species and antioxidant enzymes in spring barley (Hordeum vulgare L.) exposed to ozone. Environ. Pollut. 2002, 116, 37–47. [Google Scholar] [CrossRef]
- Shimazaki, K.-I.; Yu, S.-W.; Sakaki, T.; Tanaka, K. Differences between Spinach and Kidney Bean Plants in Terms of Sensitivity to Fumigation with NO2. Plant Cell Physiol. 1992, 33, 267. [Google Scholar] [CrossRef]
- Wang, L.; Stegemann, J.P. Extraction of high quality RNA from polysaccharide matrices using cetlytrimethylammonium bromide. Biomaterials 2010, 31, 1612–1618. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, S.; Li, W. RSeQC: Quality control of RNA-seq experiments. Bioinformatics 2012, 28, 2184–2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okonechnikov, K.; Conesa, A.; García-Alcalde, F. Qualimap 2: Advanced multi-sample quality control for high-throughput sequencing data. Bioinformatics 2016, 32, 292–294. [Google Scholar] [CrossRef] [PubMed]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [Green Version]
- Florea, L.; Song, L.; Salzberg, S.L. Thousands of exon skipping events differentiate among splicing patterns in sixteen human tissues. Research 2013, 2, 188. [Google Scholar]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein–protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Yates, A.; Akanni, W.; Amode, M.R. Ensembl 2016 Nucleic Acids Research. Oxford Acad. 2016, 44, D710–D716. [Google Scholar]
- Oulhen, N.; Foster, S.; Wray, G.; Wessel, G. Identifying Gene Expression from Single Cells to Single Genes. Methods Cell Biol. 2019, 151, 127–158. [Google Scholar]
- Schiffgens-Gruber, A.; Lütz, C. Ultrastructure of mesophyll cell chloroplasts of spruce needles exposed to O3, SO2 and NO2 alone and in combination. Environ. Exp. Bot. 1992, 32, 243–254. [Google Scholar] [CrossRef]
- Wu, G.; Shortt, B.J.; Lawrence, E.B.; Levine, E.B.; Fitzsimmons, K.C.; Shah, D.M. Disease resistance conferred by expression of a gene encoding H2O2-generating glucose oxidase in transgenic potato plants. Plant. Cell 1995, 7, 1357–1368. [Google Scholar] [CrossRef] [PubMed]
- Shalata, A.; Neumann, P.M. Exogenous ascorbic acid (vitamin C) increases resistance to salt stress and reduces lipid peroxi-dation. J. Experiment. Bot. 2001, 52, 2207–2211. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Bellaloui, N.; Sun, G.; Tigabu, M.; Wang, J. Exogenous sodium sulfide improves morphological and physiological responses of a hybrid Populus species to nitrogen dioxide. J. Plant. Physiol. 2014, 171, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.P.; Machado, E.C.; Silva, J.A.B.; Lagoa, A.M.M.A.; Silveira, J.A.G. Photosynthetic gas exchange, chlorophyll fluo-rescence and some associated metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery. Environ. Experiment. Bot. 2004, 51, 45–56. [Google Scholar] [CrossRef]
- Liu, B.; Kang, C.; Wang, X.; Bao, G. Physiological and morphological responses of Leymus chinensis to saline-alkali stress. Grassl. Sci. 2016, 61, 217–226. [Google Scholar] [CrossRef]
- Scalet, M.; Federice, R.; Guido, M.C.; Manes, F. Peroxidase activity and polyamine changes in response to ozone and simulated acid rain in Aleppo pine needles. Environ. Experiment. Bot. 1995, 35, 417–425. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, W.; He, J.; Zhang, L.; Wei, Y.; Yang, M. Nitric oxide alleviates salt stress in seed germination and early seedling growth of pakchoi (Brassica chinensis L.) by enhancing physiological and biochemical parameters. Ecotoxicol. Environ. Saf. 2020, 187, 109785. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Hou, S.; Guo, X.; Jia, J.; Yang, W.; Liu, Z.; Chen, S.; Li, X.; Qi, D.; Liu, G.; et al. A MYB-related transcription factor from sheepgrass, LcMYB2, promotes seed germination and root growth under drought stress. BMC Plant. Biol. 2019, 19, 564. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.; Xie, X.Y.; Wang, M.C.; Yang, Y.Z. Characterization of the complete chloroplast genome of Carpinus putoensis. Conserv. Genet. Resour. 2017, 9, 127–129. [Google Scholar] [CrossRef]
- Licausi, F.; Kosmacz, M.; Weits, D.; Giuntoli, B.; Giorgi, F.M.; Voesenek, L.A.C.J.; Perata, P.; Van Dongen, J.T. Oxygen sensing in plants is mediated by an N-end rule pathway for protein destabilization. Nat. Cell Biol. 2011, 479, 419–422. [Google Scholar] [CrossRef]
- Golldack, D.; Lüking, I.; Yang, O. Plant tolerance to drought and salinity: Stress regulating transcription factors and their functional significance in the cellular transcriptional network. Plant. Cell Rep. 2011, 30, 1383–1391. [Google Scholar] [CrossRef]
- Sakuma, Y.; Maruyama, K.; Qin, F.; Osakabe, Y.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Dual function of an Arabidopsis transcription factor DREB2A in water-stress-responsive and heat-stress-responsive gene expression. Proc. Natl. Acad. Sci. USA 2006, 103, 18822–18827. [Google Scholar] [CrossRef] [Green Version]
- Hinz, M.; Wilson, I.W.; Yang, J.; Buerstenbinder, K.; Llewellyn, D.; Dennis, E.S.; Sauter, M.; Dolferus, R. Arabidopsis RAP2.2: An Ethylene Response Transcription Factor That Is Important for Hypoxia Survival. Plant. Physiol. 2010, 153, 757–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abogadallah, G.M.; Nada, R.M.; Malinowski, R.; Quick, P. Overexpression of HARDY, an AP2/ERF gene from Arabidopsis, improves drought and salt tolerance by reducing transpiration and sodium uptake in transgenic Trifolium alexandrinum L. Planta 2011, 233, 1265–1276. [Google Scholar] [CrossRef]
- Zhu, Q.; Zhang, J.; Gao, X.; Tong, J.; Xiao, L.; Li, W.; Zhang, H. The Arabidopsis AP2/ERF transcription factor RAP2.6 participates in ABA, salt and osmotic stress responses. Gene 2010, 457, 1–12. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant. Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-L.; Qu, L.; Guo, D.; Wang, Y.; Zhu, J.-H.; Peng, S.-Q. Histone deacetylase interacts with a WRKY transcription factor to regulate the expression of the small rubber particle protein gene from Hevea brasiliensis. Ind. Crop. Prod. 2020, 145, 111989. [Google Scholar] [CrossRef]
- Burner, J.; West, B.; Mayer, P.M. What do we expect from the dissociation of ionized nitro-substituted polycyclic aromatic hydrocarbons in the interstellar medium? Int. J. Mass Spectrom. 2019, 434, 81–86. [Google Scholar] [CrossRef]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Skriver, K. NAC transcription factors: Structurally distinct, functionally diverse. Trends Plant. Sci. 2005, 10, 79–87. [Google Scholar] [CrossRef]
- Wang, M.; Zou, Z.; Li, Q.; Sun, K.; Chen, X. The CsHSP17.2 molecular chaperone is essential for thermos tolerance in Camellia sinensis. Sci. Rep. 2017, 7, 1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, J.M.; Bukhari, S.A.H.; Zeng, J.B.; Quan, X.Y.; Ali, E.; Muhammad, N.; Zhang, G.-P. Nitrogen (N) metabolism related enzyme activities, cell ul-trastructure and nutrient contents as affected by N level and barley genotype. J. Integrat. Agric. 2017, 16, 190–198. [Google Scholar] [CrossRef]
- Mukherjee, A.; Agrawal, S.B.; Agrawal, M. Responses of tropical tree species to urban air pollutants: ROS/RNS formation and scavenging. Sci. Total Environ. 2020, 710, 136363. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Kargul, J.; Barber, J. Photosynthetic acclimation: Structural reorganization of light harvesting antenna–role of redox-dependent phosphorylation of major and minor chlorophyll a/b binding proteins. FEBS J. 2008, 275, 1056–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahl, A.; Kahl, G. Air pollutant stress changes the steady-state transcript levels of three photosynthesis genes. Environ. Pollut. 1995, 88, 57–65. [Google Scholar] [CrossRef]
- Xu, Q.; Zhou, B.; Ma, C.; Xu, X.; Xu, J.; Jiang, Y.; Liu, C.; Li, G.; Herbert, S.J.; Hao, L. Salicylic Acid-Altering Arabidopsis Mutants Response to NO2 Exposure. Bull. Environ. Contam. Toxicol. 2010, 84, 106–111. [Google Scholar] [CrossRef]
- Khare, T.; Kumar, V.; Kishor, P.B.K. Na+ and Cl− ions show additive effects under NaCl stress on induction of oxidative stress and the responsive anti-oxidative defense in rice. Protoplasma 2015, 252, 1149–1165. [Google Scholar] [CrossRef] [PubMed]
- An, J.P.; Li, R.; Qu, F.J.; You, C.X.; Wang, X.F. Ectopic expression of an apple cytochrome P450 gene MdCYPM1 negatively regulates plant photo morphogenesis and stress response in Arabidopsis. Biochem. Biophys. Res. Commun. 2017, 483, 1–9. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | No. | ≥500 bp | ≥1000 bp | N50 | N90 | Maximum Length | Minimum Length | Total Length | Average Length | CG Content |
|---|---|---|---|---|---|---|---|---|---|---|
| Transcript | 529,540 | 124,713 | 29,088 | 470 | 231 | 7490 | 201 | 225,567,341 | 425.97 | 40–50% |
| Unigene | 250,200 | 41,790 | 9609 | 381 | 221 | 7490 | 201 | 94,258,132 | 376.73 | 60–70% |
| Database | Number of Genes | Percentage (%) |
|---|---|---|
| CDD | 79,760 | 31.88 |
| KOG | 64,226 | 25.67 |
| NR | 94,267 | 37.68 |
| NT | 77,874 | 31.12 |
| PFAM | 51,696 | 20.66 |
| Swiss-prot | 103,389 | 41.32 |
| TrEMBL | 93,882 | 37.52 |
| GO | 110,530 | 44.18 |
| KEGG | 9284 | 3.71 |
| At least one database | 136,276 | 54.47 |
| All database | 4595 | 1.84 |
| DEGs | DEG Number | CDD | KOG | GO | KEGG | NR | NT |
|---|---|---|---|---|---|---|---|
| Upregulated genes | 1119 | 1119 | 330 | 690 | 91 | 740 | 597 |
| Downregulated genes | 1240 | 1240 | 547 | 996 | 186 | 1090 | 760 |
| Total | 2359 | 2359 | 877 | 1686 | 277 | 1830 | 1357 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheng, Q.; Liu, C.; Song, M.; Xu, J.; Zhu, Z. Comprehensive Transcriptome Analysis of Rare Carpinus putoensis Plants under NO2 stress. Genes 2021, 12, 754. https://doi.org/10.3390/genes12050754
Sheng Q, Liu C, Song M, Xu J, Zhu Z. Comprehensive Transcriptome Analysis of Rare Carpinus putoensis Plants under NO2 stress. Genes. 2021; 12(5):754. https://doi.org/10.3390/genes12050754
Chicago/Turabian StyleSheng, Qianqian, Congzhe Liu, Min Song, Jingyuan Xu, and Zunling Zhu. 2021. "Comprehensive Transcriptome Analysis of Rare Carpinus putoensis Plants under NO2 stress" Genes 12, no. 5: 754. https://doi.org/10.3390/genes12050754