1. Introduction
Signaling pathways Wnt and TGF-beta are involved in the formation of axes in multicellular animals: they provide axial patterning in developing embryos and support it in adult animals. In addition to development, both signaling cascades are involved in the regeneration of multicellular animals of different levels of complexity—from cnidarians to vertebrates. Wnt and TGF-beta pathways regulate cell proliferation, migration, differentiation, and apoptosis during various regenerative processes [
1,
2,
3,
4,
5].
Sponges (Porifera) occupy a key position at the base of the phylogenetic tree of animals. With this in mind, they are essential for understanding the origin and evolution of multicellular body patterning. The axes in the body of sponges are clearly different from those in other multicellular animals because most sponges have a polarity axis but lack an axis of symmetry [
6]. Interestingly, sponges have genes of the Wnt and TGF-beta pathways, and these genes are even involved in establishing axes in embryo and adult bodies. Ligands of both pathways have dynamic expression patterns during embryonic development and in the larva of
Amphimedon queenslandica and
Sycon ciliatum. In addition, in the adult
S. ciliatum, the expression of several Wnt and TGF-beta is confined to the osculum and the entrances of the radial choanocyte chambers, i.e., axial structures [
7,
8,
9].
However, the possible involvement of Wnt and TGF-beta in regeneration after injury in sponges has not been studied. Experiments with reparative regeneration are convenient only for sponges with a clear morphological polarity because, in this case, we can assume what place an excised area occupies with respect to the axes of the body. In massive multioscular leuconoid sponges, it is difficult to understand what position on the axis an excised body fragment occupied. That complicates the interpretation of the experiment results. In this case, another type of regeneration, whole-body regeneration (WBR) after tissue dissociation, i.e., cell reaggregation, provides a convenient model for studying polarity formation.
The original anatomical structure and patterning of the sponge body are completely disrupted during tissue dissociation. During the process of cell reaggregation, separated cells coalesce, forming structureless primary multicellular aggregates. Subsequently, these aggregates undergo a transition to radially symmetric, developing primmorphs with an aquiferous system, and then to a reconstructed functional sponge with a polarity axis formed de novo. This model has been thoroughly described at the morphological level in some sponge species (reviewed in [
10,
11]) but studied from the molecular point of view only in one species of calcareous sponges [
12].
We began to study the participation of Wnt and TGF-beta signaling pathways in the cell reaggregation in demosponge Halisarca dujardinii with identification of pathways’ components in the sponge transcriptome, subsequent domain and phylogenetic analysis of orthologous proteins, and evaluation of gene expression levels at different stages of reaggregation using RNA-seq and whole-mount in situ hybridization (WMISH). Our preliminary results, including the first WMISH data on primmorphs, show differential dynamic expression of Wnt and TGF-beta components at different stages of reaggregation, as well as a polar localization for several transcripts.
3. Results
3.1. Wnt Pathway Components in Halisarca dujardinii
We have previously identified Wnt ligands in the
H. dujardinii transcriptome and described their phylogenetic position as well as expression in the adult sponge and larva [
13]. We found 10 Wnt ligands containing conserved cysteine residues and an RWNC motif. Phylogenetic analysis showed that only one of them could be close to the Wnt (namely AquWntC) of another demosponge,
Amphimedon queenslandica, and thus the molecules were named alphabetically, starting with HduWntC. Here, we searched the transcriptome for the main participants in the intracellular signal transduction in the Wnt cascade: Frizzled receptors and LRP coreceptors, a secondary messenger Dishevelled, transcriptional cofactors (beta-catenin, TCF, Groucho), a protein complex responsible for beta-catenin degradation, and enzymes responsible for Wnt ligand maturation (porcupine and Wntless).
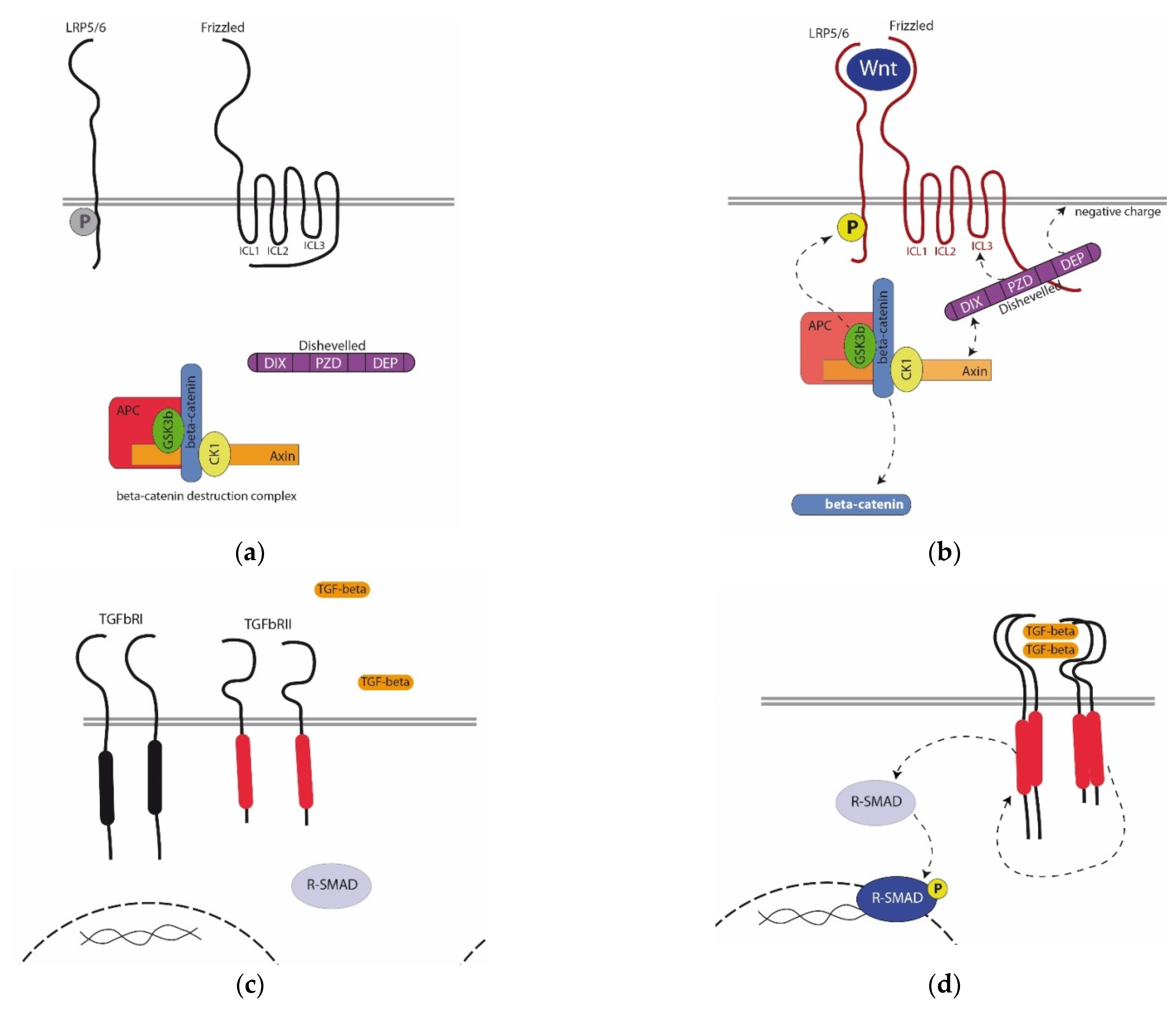
Many models for the regulation of the activity of the Wnt cascade have been described. The simplest ON/OFF model is as follows. In the absence of a signal (OFF state), the pool of the beta-catenin is constantly degraded due to the beta-catenin destruction complex (bCDC) in the cytoplasm of a competent (responding to a signal) cell (
Figure 1a). bCDC, consisting of the core proteins Axin and APC, glycogen synthase kinase-3 beta (GSK3b), and casein kinase 1 (CK1), phosphorylates cytoplasmic beta-catenin, which leads to its ubiquitin-dependent degradation in proteasomes. In the instructing (generating a signal) cell, prior to secretion, the Wnt ligand is transported and acylated in the Golgi apparatus by Wntless protein and palmitoleoyltransferase porcupine, respectively. Acylation is absolutely necessary for the binding of the Wnt ligand to the receptor. Blocking these enzymes by knockout or drugs makes signal transmission impossible. On the surface of a competent cell, the ligand binds to the Frizzled receptor and LRP5/6 coreceptor (
Figure 1b). Frizzled is a seven-pass transmembrane protein, and LRP is a single-pass one. The cytoplasmic regions of both proteins contain sequences responsible for interaction with cytoplasmic signaling participants: the third intracytoplasmic loop (ICL3) and C-end of Frizzled responsible for interaction with Dishevelled protein, whereas the cytoplasmic end of LRP5/6 contains sites phosphorylated by GSK3b from bCDC. The binding of the ligand to the receptor and coreceptor leads to conformational changes in the cytoplasmic domains of the Frizzled and recruitment of Dishevelled from the cytoplasm. Dishevelled, in turn, recruits bCDC, resulting in its break into individual proteins and loss of the ability to phosphorylate beta-catenin. beta-catenin is preserved from degradation and migrates into a nucleus, regulating the expression of target genes as a transcriptional cofactor [
21].
Using reciprocal BLAST, we identified six Frizzled paralogs, four SFRP paralogs, and one protein for each of LRP5/6, porcupine, Wntless, Dishevelled, APC, TCF, Groucho, GSK3b, CK1, and beta-catenin. No Axin, one of the core components of the bCDC, acting as a kernel in the assembly of the complex, was found in the H. dujardinii transcriptome.
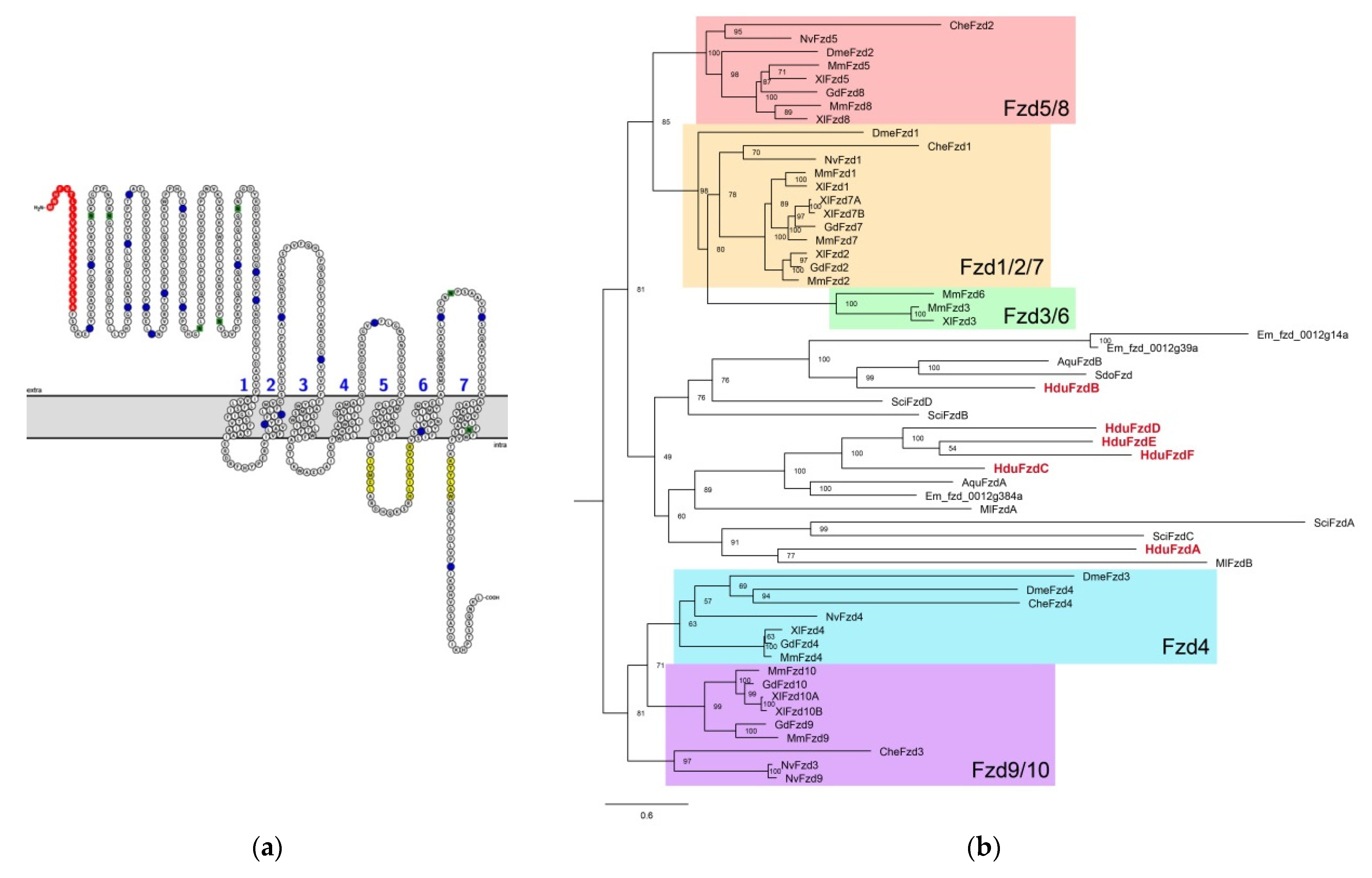
The N-end of the Frizzled receptor facing the intercellular space contains an extended cysteine-rich domain (CRD) responsible for the binding of the Wnt ligand (
Figure 2a).
All six
H. dujardinii Frizzled have a secondary structure typical for this receptor: an extracellular CRD domain, seven transmembrane domains, conservatively located cysteines in loops between transmembrane domains, and motifs in intracellular regions responsible for interaction with Dishevelled (
Figure 2a).
Phylogenetic analysis of the evolutionary relationships of the identified sponge
Frizzled genes showed a situation similar to the position of Wnt ligands: we cannot assign them to any of the receptor families described in higher metazoans (
Figure 2b). In this regard, by analogy with ligands, we named receptors in alphabetical order.
In addition to available
Sycon ciliatum,
A. queenslandica, and
Suberites domuncula sequences from sponges, and
Mnemiopsis leidyi from Ctenophora, we identified
Frizzled sequences in the recently sequenced demosponge
Ephydatia muelleri genome [
22]. Three sequences were obtained by tblastn with consequent HMMSCAN against Pfam database and TMHMM analysis. These three Frizzleds represent seven-pass transmembrane receptors with a CRD domain in the extracellular part of the peptide.
The branching order of the different families is conserved between the different trees, and family groupings are generally well supported. There appear to be five bilaterian families: Fzd1/2/7, Fzd3/6, Fzd5/8, Fzd4, and Fzd9/10. The nomenclature of some analyzed nonbilaterian genes is misleading:
CheFzd2 and
DmeFzd2 fall into the Fzd5/8 branch,
DmeFzd3 fall into the Fzd4 family, and
CheFzd3 and
NveFzd3 fall into the Fzd9/10 family. The last two look like mistakes of the phylogenetical classification of
Clytia hemisphaerica,
Nematostella vectensis, and
Drosophila melanogaster sequences because the Fzd3/6 family seems to be a chordate innovation [
8].
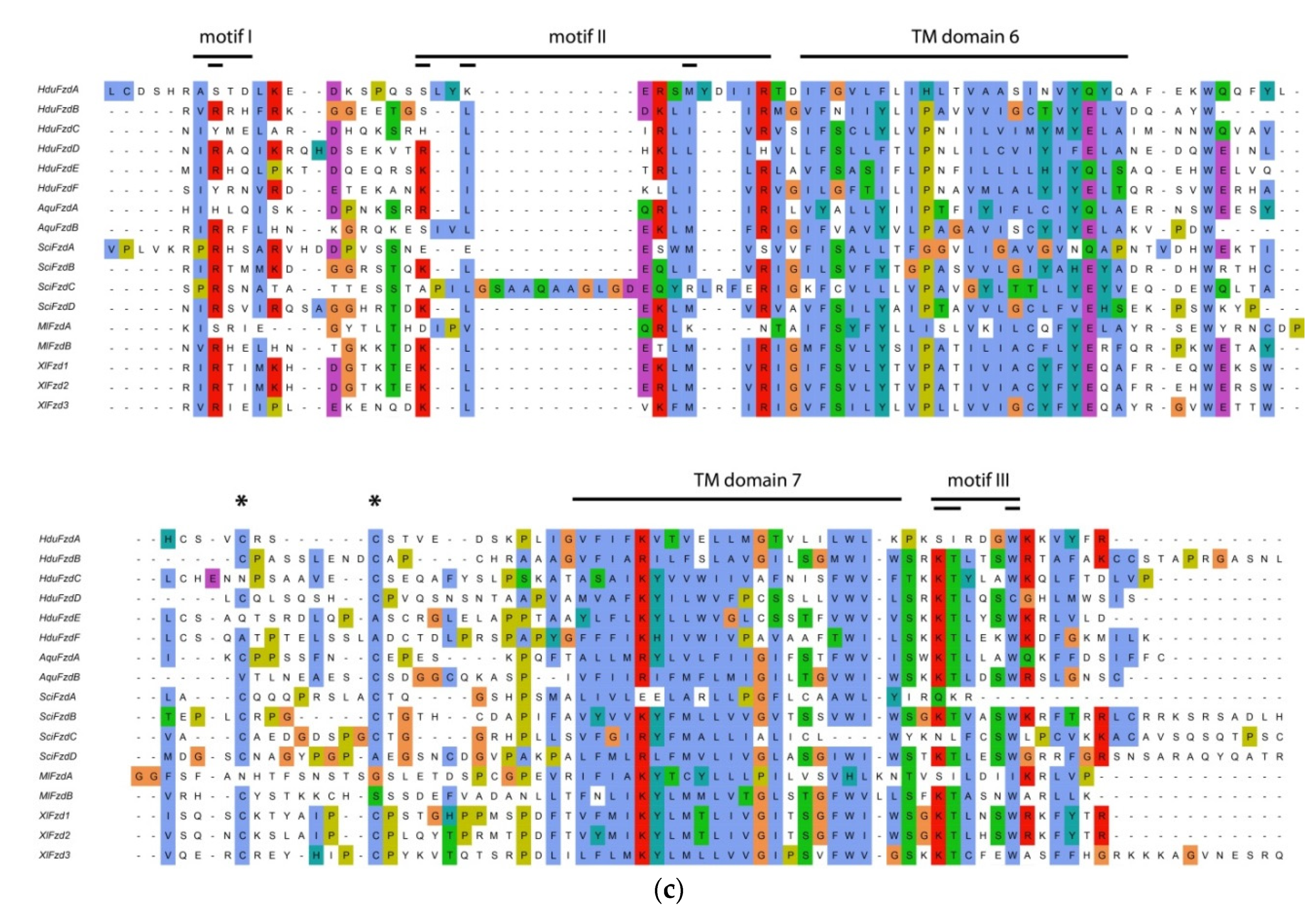
Frizzled contains three motifs for interaction with the PZD domain of the Dishevelled protein. ICL3 of Frizzled contains two of these motifs—motif 1 (IRxV) and motif 2 (KLEKLMVR,
Figure 2a,c). Most of the invariant amino acids in these motifs occupy positions in the alignments that are conservative with respect to vertebrates. There are two conserved cysteine residues between TM domains 6 and 7; the first of them is replaced with non-similar amino acid residue only in
HduFzdC and
HduFzdE (
Figure 2c). The same substitution is observed in
FzdA of the ctenophore
M. leidyi [
23]. The third motif (KTxxxW) is located in the free cytoplasmic C-end of Frizzled. Here invariant amino acid residues are substituted in
HduFzdA, as well as in
MleFzdA and
SciFzdA. This Frizzled-specific motif has been shown to directly interact with the PDZ domain of Dishevelled [
24]. Thus, in
H. dujardinii, the majority of Frizzled proteins demonstrate conservatism at the level of functionally essential protein sequences ensuring signal transduction into the cytoplasm.
HduFzdA seems to be an exception, as it contains several non-analogous substitutions in the motifs.
There are also secreted Frizzled-related proteins (SFRPs) that have a CRD but lack transmembrane domains. They represent soluble forms of the receptor secreted by a cell and act as ligand traps because they bind diffusing Wnt and prevent it from binding to functional receptors. We identified four such proteins that have a CRD with a high level of similarity to Frizzled. However, the number of CRD-containing proteins is large, and their targets are heterogeneous because interaction occurs between two cysteine-rich regions of the polypeptide chain. Probably other CRD-containing proteins can act as functional analogs of SFRPs, among the described ones (for example, Dickkopf) [
25]. The regulation of signaling activity at the level of ligand–receptor binding is a rather broad topic, and thus we leave the analysis of SFRPs for further research.
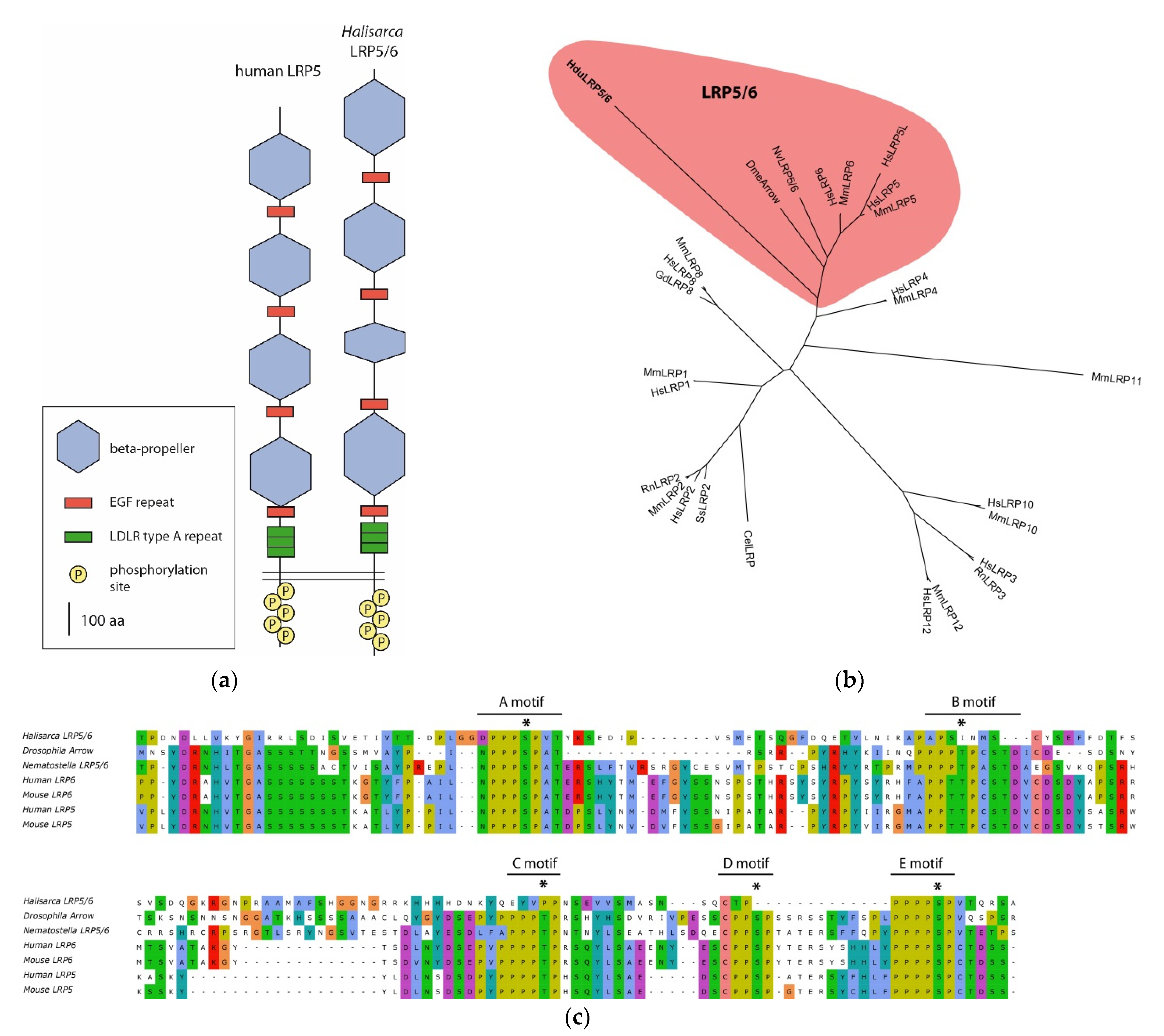
The Wnt coreceptor belongs to the family of low-density lipoprotein receptors (LDLRs), only the 5 and 6 classes (LRP5/6) of which are involved in the Wnt cascade. LRP5/6 are characterized by a conservative set and arrangement of domains. We found several LDLRs and focused on them because LRP5/6 is responsible for recruiting cytoplasmic protein Axin—a core component of the bCDC. Due to the absence of Axin in
H. dujardinii, it seemed important to understand whether it has a structurally and functionally conservative LRP5/6. Only one of the found LDLRs possessed the complete set of distinctive features of LRP5/6 (
Figure 3a): the extracellular part of the receptor contains four tandems of a beta-propeller and EGF repeats. A beta-propeller is a domain that has a tertiary structure resembling a six-bladed propeller, with each blade consisting of four parallel beta-sheets. Each beta-propeller consists of five LDLR class B1 repeats. We were not able to identify all character repeats of class B1 with HMMSCAN, and therefore additionally we predicted the secondary structure of peptide with JPred4 [
26]. The position and number of beta-sheet sets coincided with the expected position of beta-propellers in the peptide, and their predicted tertiary organization corresponds to a beta-propeller (
Supplementary File S7). In the juxtamembrane region of the receptor, there are conserved three repeats of LDLR class A. The receptor’s cytoplasmic domain contains five PPPSPxS motifs, which are phosphorylated by GSK3b and CK1 upon activation of the cascade. These motifs mediate interaction with Axin [
27,
28,
29,
30,
31]. Alignment with proteins of mammals,
D. melanogaster, and
N. vectensis shows that conservative phosphorylated amino acid residues are present in four out of five motifs (
Figure 3c). In the phylodendrogram, the LRP5/6 of the
H. dujardinii falls in the clade with the Arrow protein of
D. melanogaster, LRP5/6 of the
N. vectensis, and LRP classes 5 and 6 of mammals.
3.2. TGF-Beta Pathway Components in Halisarca dujardinii
The TGF-beta superfamily includes more than 30 structurally related proteins that are morphogens similarly to Wnt. Several families are distinguished in this superfamily: TGF-beta itself, several BMP families, Lefty, Activin/Inhibin, Vg1, a number of Gdf, and others. All ligands from TGF-beta superfamily synthesized as a precursor protein. This precursor contains three domains—the N-terminal signal peptide, propeptide (or latency-associated peptide), and the C-terminal mature peptide. During processing, the mature peptide is cleaved by Furin, a convertase, at the RxxR conserved site [
32]. The mature peptide forms a dimer, which binds to the specific TGF-beta receptor (
Figure 1c,d; [
33]). The TGF-beta receptor is a single-pass membrane protein containing a cytoplasmic kinase domain with both serine/threonine kinase activity and tyrosine kinase activity [
34]. There are two types of the TGF-beta receptors—Type II receptor, constitutively active, and Type I receptor. When the dimeric ligand binds, a complex is formed of two Type I receptor molecules and two Type II receptor molecules. Constitutively active Type II receptor then phosphorylates a TGF-b Type I receptor. Phosphorylated Type I receptors phosphorylate and thereby activate receptor-associated SMAD proteins (R-SMADs), including SMAD1/5 and SMAD2/3 [
35,
36]. R-SMAD proteins are composed of two main functional domains: the Mad-homology domains 1 and 2 (MH1 and MH2). Activated R-SMADs interact with the common-mediator SMAD (Co-SMAD, SMAD4) and become translocated to the nucleus. This SMAD protein complex then regulates the expression of TGF-beta target genes by interacting with transcription factors. Inhibitory SMADs (I-SMADs, SMAD-6) are able to compete with Co-SMADs in interaction with R-SMADs, thus inhibiting the signaling pathway.
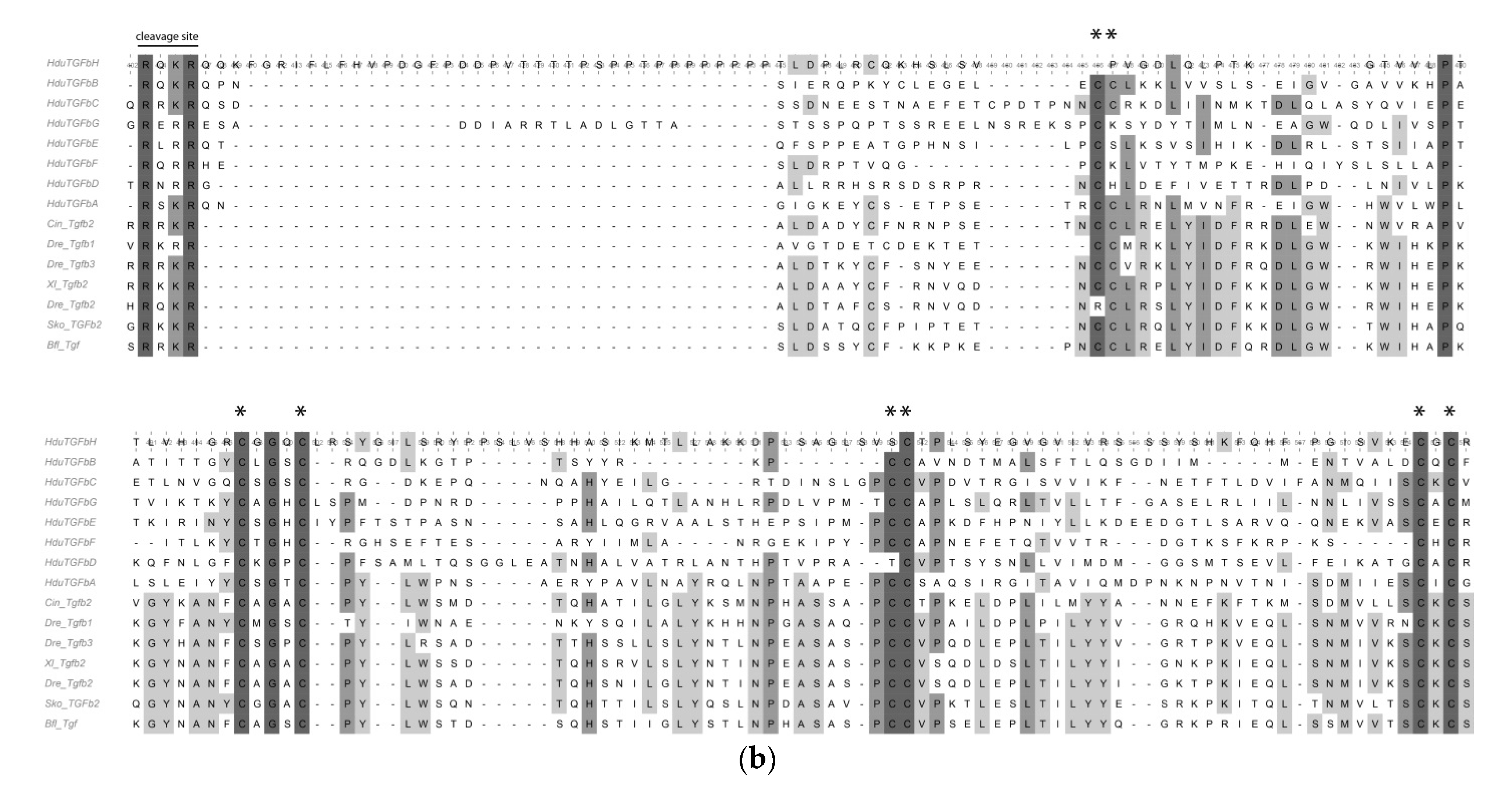
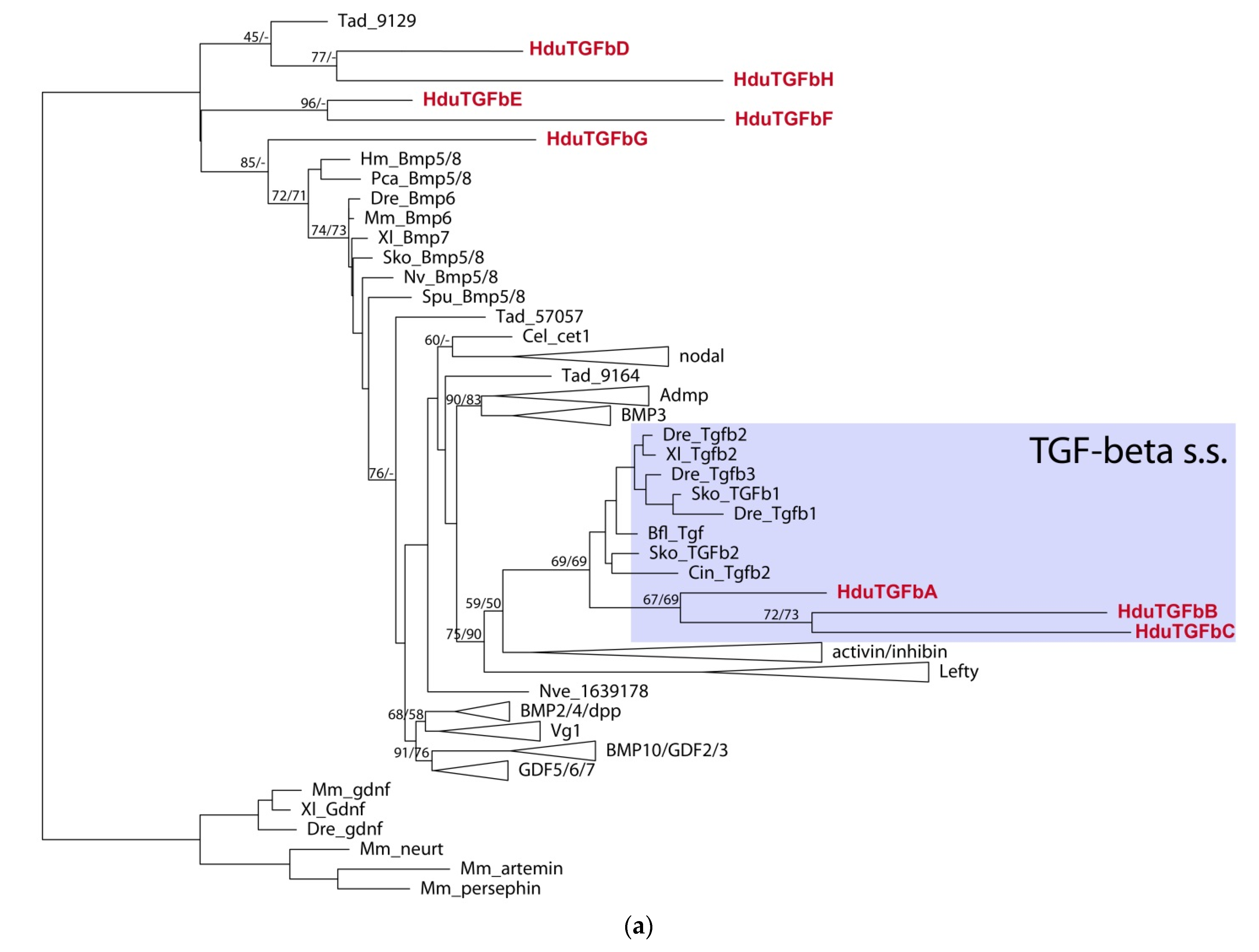
We were able to identify and isolate eight putative TGF-beta ligands, six receptors, and six SMADs. Due to the relatively high divergence of the sponge sequences, only three ligands,
HduTGFbA–
HduTGFbC, could be ascribed to TGF-beta sensu stricto, while the rest five sequences (
HduTGFbD–
HduTGFbH) group as sister to the other TGF-beta families, with the closest TGF member being one of
Trichoplax adhaerens ligands (
Figure 4a).
HduTGFbA,
HduTGFbB, and
HduTGFbC have eight cysteine residues, conserved in gene families of the TGF-beta-related clade (
Figure 4b).
HduTGFbG,
HduTGFbE, and
HduTGFbF have seven conserved cysteine residues, while
HduTGFbD has only six, and
HduTGFbH missed first, second, and fifth cysteine residues and have only five of them. Interestingly, when we ran an analysis on a full TGF-beta precursor with both the propeptide and the peptide domain, only
HduTGFbA fell into the TGF-beta s.s. clade and the rest of the ligand groups at the base of the TGF-beta tree.
Three pairs of genes appear to be relatively recent tandem duplications since they group closely together and are located adjacent to each other on the same contig of the genome draft: HduTGFbB/HduTGFbC, HduTGFbD/HduTGFbH, and HduTGFbE/HduTGFbF. The rest two ligands, HduTGFbA and HduTGFbG, are located on separate contigs (data not shown).
Sequence analyses using SignalP and HMMSCAN predicted signal peptides and TGF-beta peptides for HduTGFbA–HduTGFbD and HduTGFbG–HduTGFbH. We suggest that the propeptides are missing in these cases or are highly divergent and not detected by homology searches. For HduTGFbE and HduTGFbF, a propeptide and TGF-b peptide are predicted, but the signal sequence is not. The cleavage site for furin protease (RxxR) at the border of the mature peptide is clearly present for all ligands.
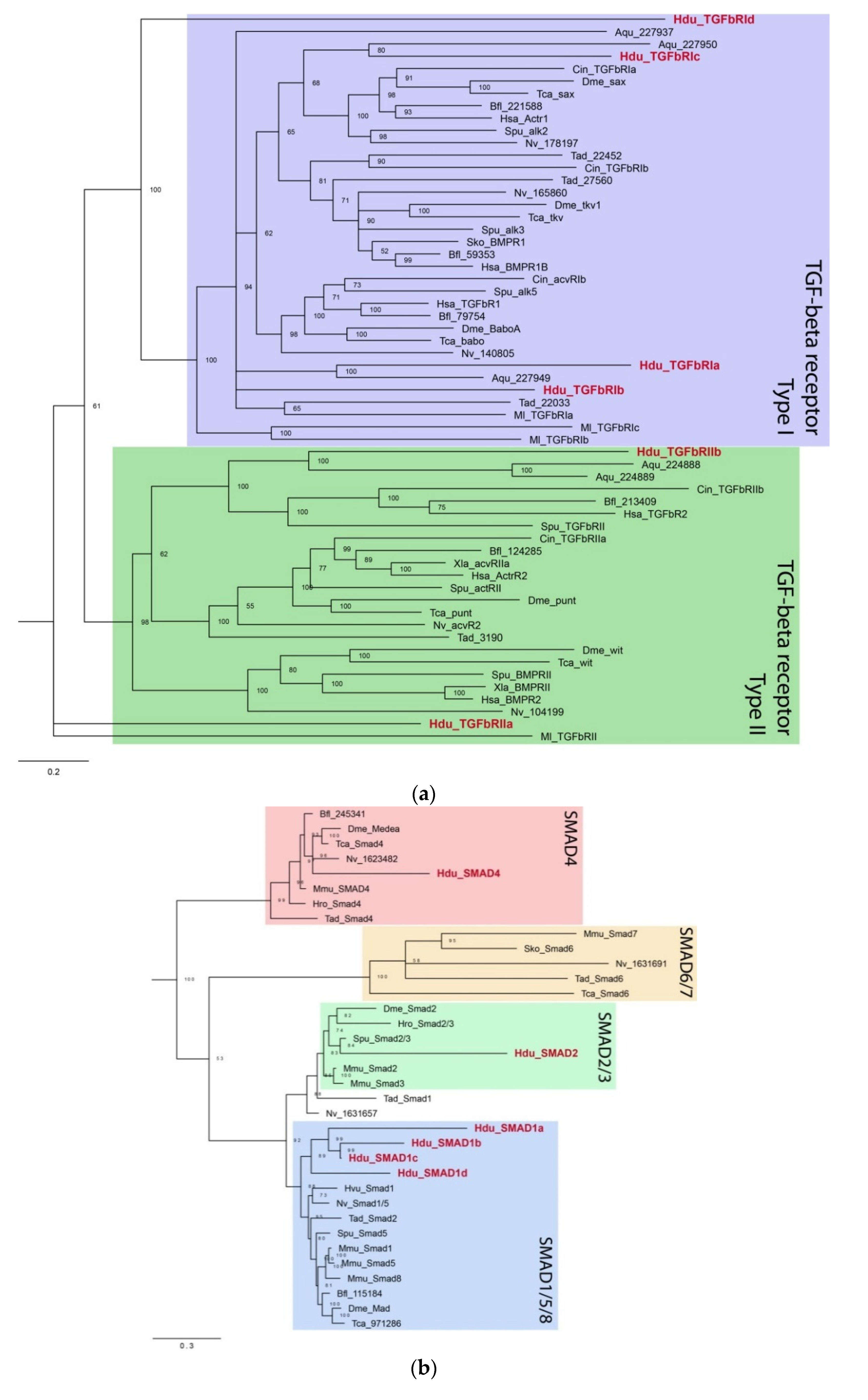
There are two Type II receptors (
HduTGFbRIIa and
HduTGFbRIIb) and four Type I receptors (
HduTGFbRIa–
HduTGFbRId). All contain the extracellular receptor domain, single-pass transmembrane domain, and intracellular kinase domain. Additionally, all
H. dujardinii Type I receptors possess the glycine–serine repeat (GS region) adjacent to the kinase domain, an arrangement that is characteristic of metazoan Type I receptors. Phylogenetic analyses included sequences of TGF-beta receptors from representative metazoans and showed that
H. dujardinii receptors are not supported in individual receptor subclasses (
Figure 5a). Instead,
HduTGFbRIIa appears sister to all other Type II receptors and
HduTGFbRId as a sister to all Type I subclasses.
HduTGFbRIc together with
A. queenslandica sequence form a weakly supported group with activating receptors Type I (sax/Actr1/alk2).
HduTGFbRIa and
HduTGFbRIb occupy the base of the Type I receptors subtree together with
A. queenslandica and
M. leidyi sequences.
HduTGFbRIIb forms a well-supported clade with two
A. queenslandica sequences, lancelet, human, sea urchin, and sea squirt Type II receptors.
We were also able to detect six SMAD proteins (
Figure 5b). There are five receptor SMADs, one belonging to the SMAD2/3 family (
HduSMAD2), and four SMADs forming a sister group to the SMAD1/5/8 clade (
HduSMAD1a–
HduSMAD1d). There is also a single Co-Smad (
HduSMAD4). No inhibitory SMAD belonging to SMAD6 was observed. All SMADs have the predicted MH1 and MH2 domains, characteristic of SMAD proteins.
3.3. Stages of Cell Reaggregation in Halisarca dujardinii
Cell reaggregation in H. dujardinii represents a development process leading from a suspension of single cells to a fully functional sponge. This process occurs through several highly stereotypic stages. However, cell reaggregation shows high plasticity in some aspects, leading to the heterogeneity of the aggregates. Firstly, the rate of cell reaggregation usually demonstrates variations, especially at later stages of the process. The rate of cell reaggregation varies not only in cell cultures obtained from different individuals but also within a single culture. That means that a culture will always represent an intermix of aggregates at slightly different developmental stages at a given time point. Another source of the aggregate heterogeneity is their size, as a culture usually includes aggregates varying from hundreds of micrometers to several millimeters in diameter.
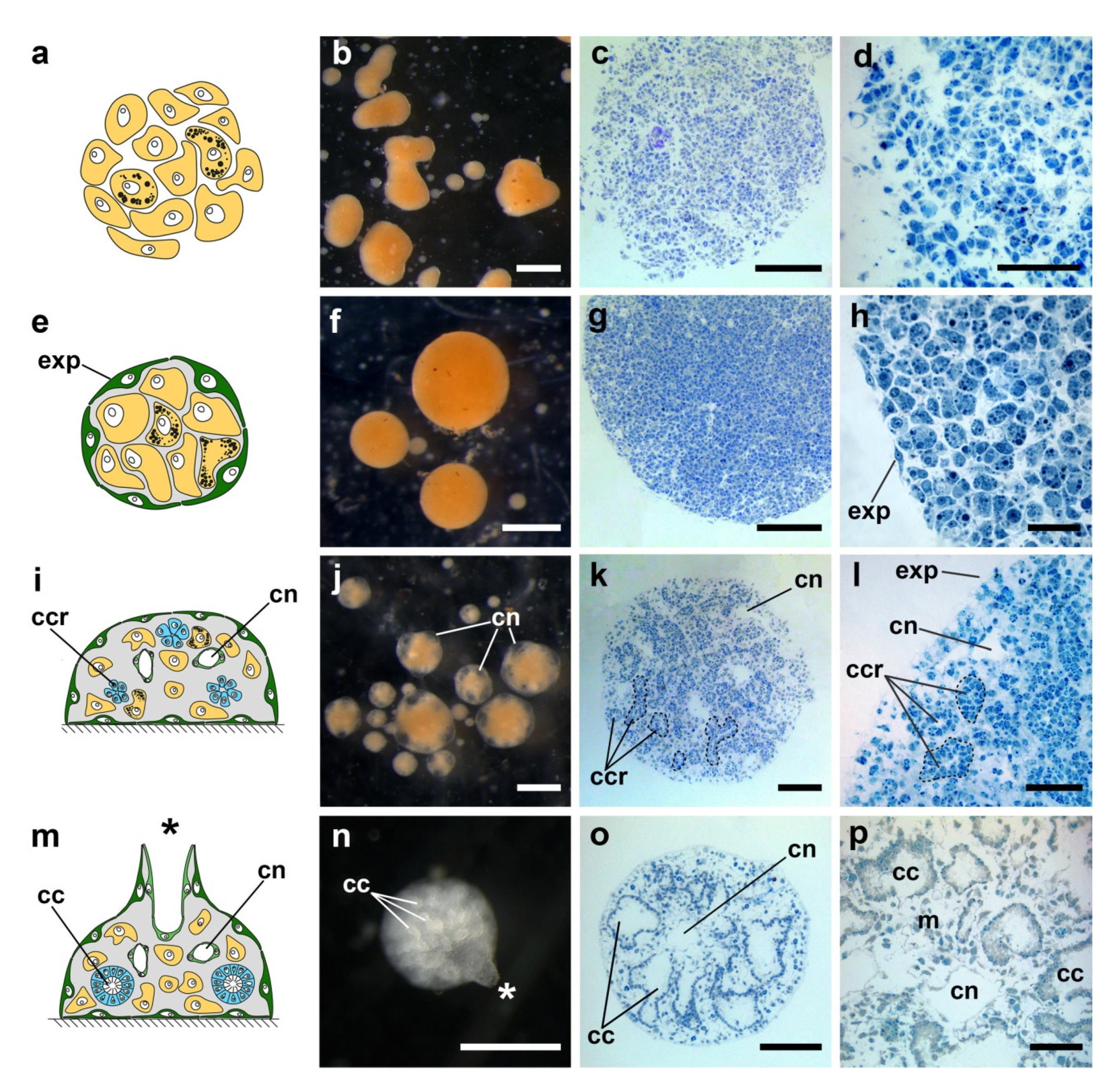
Nevertheless, the accurate in vivo studies supplemented with histological and ultrastructural investigation allow us to reliably identify several key and stereotypic stages of the cell reaggregation in
H. dujardinii (
Figure 6):
(1) Primary multicellular aggregates (PMAs), 0–24 h post-dissociation (hpd). The first small PMAs appear in a culture after 20–30 min of cultivation due to random contacts between single cells. During subsequent 24 h, they grow incorporating new cells or merging with each other. The distinguishable features of PMAs are an irregular shape, rough surface, and absence of any signs of internal structure, as PMAs represent a simple intermix of different cells.
(2) Early-stage primmorphs (ESPs) and true primmorphs (TPs), 1–3 days post-dissociation (dpd). Starting from 1 dpd, PMAs begin their transformation into ESPs and TPs. During the transformation, the aggregate surface is gradually covered by exopinacoderm, resulting in the isolation of the aggregate’s internal milieu. ESPs are aggregates with surface partially covered by exopinacoderm, while TPs are characterized by the complete exopinacoderm. As the rate of the exopinacoderm formation varies between aggregates, a culture after 1 dpd represents an intermix of aggregates at various stages of epithelization, including both ESPs and TPs. After 3 dpd, the epithelization in the vast majority of aggregates is complete, and culture predominantly contains TPs.
(3) Developing primmorphs (DPs), 5 + dpd. After a lag phase of various lengths, TPs start their progressive development to reconstruct functional sponges. This process begins with the formation of numerous small cavities in the internal parts of a primmorph. These cavities are the first sign of aquiferous system formation. Further, they give rise to canals of the aquiferous system.
(4) Progressed developing primmorphs (PDPs), 7 + dpd. As progressive development of primmorphs proceeds, rudiments of choanocyte chambers appear inside PDPs. These rudiments are dense spherical groups of cells, which will differentiate into choanocytes later on. Simultaneously, with the formation of choanocyte chamber rudiments, cavities gradually grow in size and obtain an endopinacocyte lining characteristic for mature canals of the aquiferous system.
(5) Reconstructed functional sponges (FSs), 10 + dpd. At this stage, cell reaggregation ends, and each aggregate transforms into a functional sponge with a complete aquiferous system: numerous ostia in exopinacoderm, elaborate net of canals and choanocyte chambers, and one or several oscular tubes.
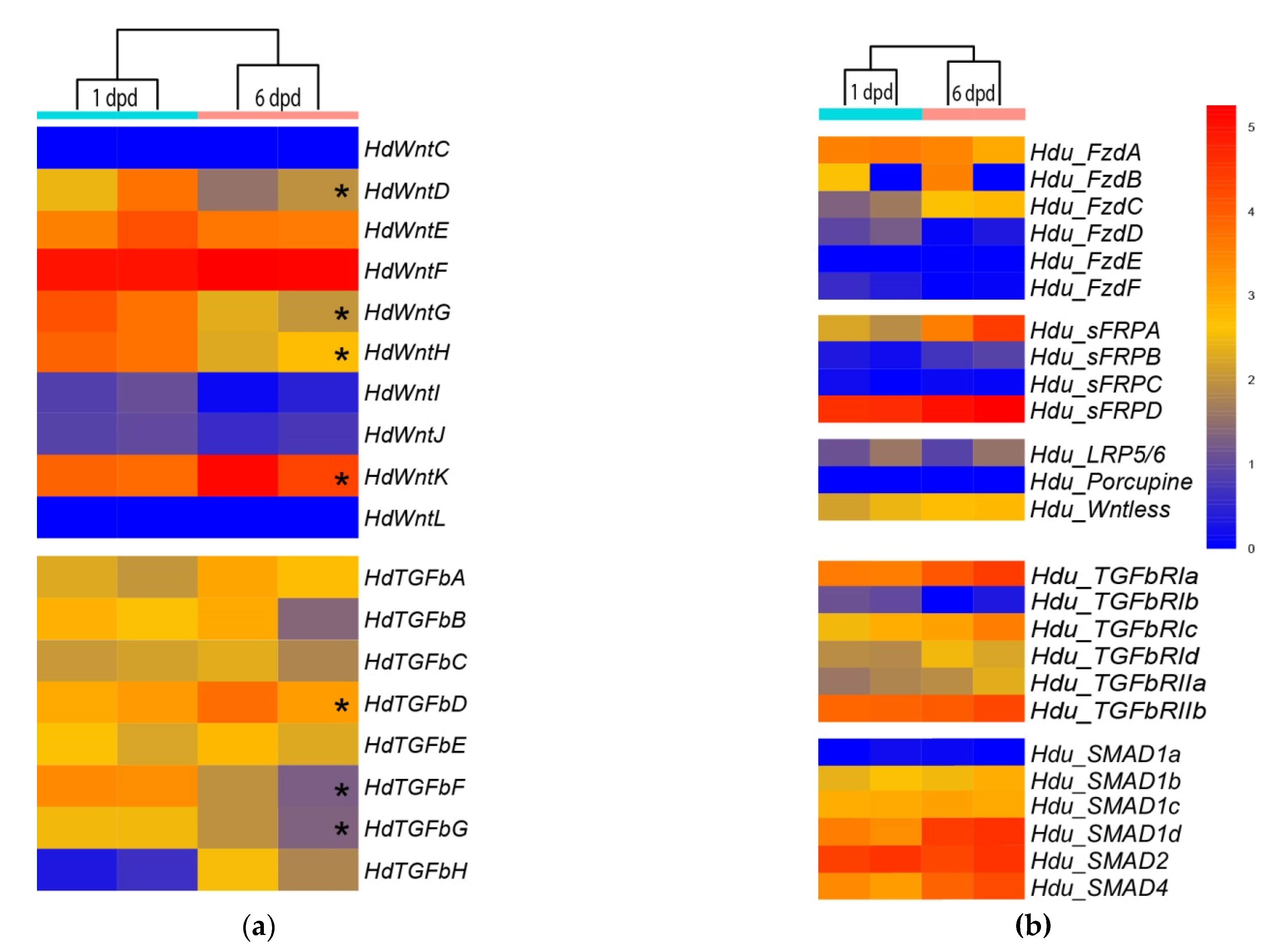
3.4. RNA-Seq Analysis of Wnt and TGF-Beta Pathway Gene Expression
We studied the expression of the identified genes at two time points during cell reaggregation of H. dujardinii, at stages of ESP/TP, 1 day post dissociation (dpd), and DP, 6 dpd, using RNA-seq technology. These time points were chosen as the key stages of the cell reaggregation: the isolation of their internal milieu through the formation of the exopinacoderm occurs at 1 dpd; the development of an aquiferous system and other anatomical structures, which are probably underlain by re-establishment of positional information and determination of new organism polarity, occurs at 6 dpd. The bulk of aggregates from three independent cultures was taken for stages 1 dpd and 7 dpd. After sequencing, transcript quantification, and principal component analysis, one sample from each stage was omitted as highly diverged, and only two samples for each stage were processed for differential expression analysis.
To visualize the expression of the identified
Wnt and
TGF-beta ligands, receptors,
LRP5/6,
SMADs, and some involved proteins during primmorph development, we generated a heatmap representation of expression. We were particularly interested in whether the elevation of
Wnt and
TGF-beta expression (including local) correlates with the interruption of radial symmetry during primmorph development. The majority of
Wnts (8 out of 10) show no differences in expression between ESP/TP and DP stages (
Figure 7a). Five genes (
HduWntD, HduWntE, HduWntI, HduWntJ, HduWntK) are expressed at the medium level, two genes (
HduWntC,
HduWntL) at a low level, and one gene (
HduWntF) at a high level at both stages (
Figure 7a). In contrast,
HduWntG and
HduWntH show significant differences between studied stages, being represented in RNA of ESP/TP samples.
TGF-beta does not show such dramatic differences for any transcript. Among them, two molecules are present at a higher level at the ESP/TP stage and another one at the DP stage.
Some differences in expression levels are also observed between paralogs of Frizzled, SFRP, TGF-beta receptors, and SMADs:
HduFzdA,
HduSFRPD,
HduTGFbRIa,
HduTGFbRIIb, and
HduSMAD2 are actively expressed at both analyzed stages, while
HduFzdD,
HduFzdE,
HduFzdF,
HduSFRPC, and
HduSMAD1a are presented at minimal levels (
Figure 7b). Intriguingly, the expression levels of
porcupine and
Wntless, proteins necessary for the Wnt secretion, differ by two orders of magnitude between each other but do not differ between the stages.
Dishevelled,
TCF, and
APC are expressed at baseline levels, while
Groucho and
GSK3b are expressed at a high level. All of them show no differences between the developmental stages. Thus we demonstrate that some Wnt and TGF-beta molecules express during whole-body regeneration. Although their expression levels demonstrate wide variations even between replications, evidently expression level for the same signaling molecules change over time. Pay attention to the bulk nature of samples from primmorph culture, heterogeneity, and WMISH results (next paragraph) that we consider to carefully interpret quantitative data about
Wnt and
TGF-beta genes expression.
3.5. Wnt and TGF-Beta Expression during Primmorph Development in Halisarca dujardinii
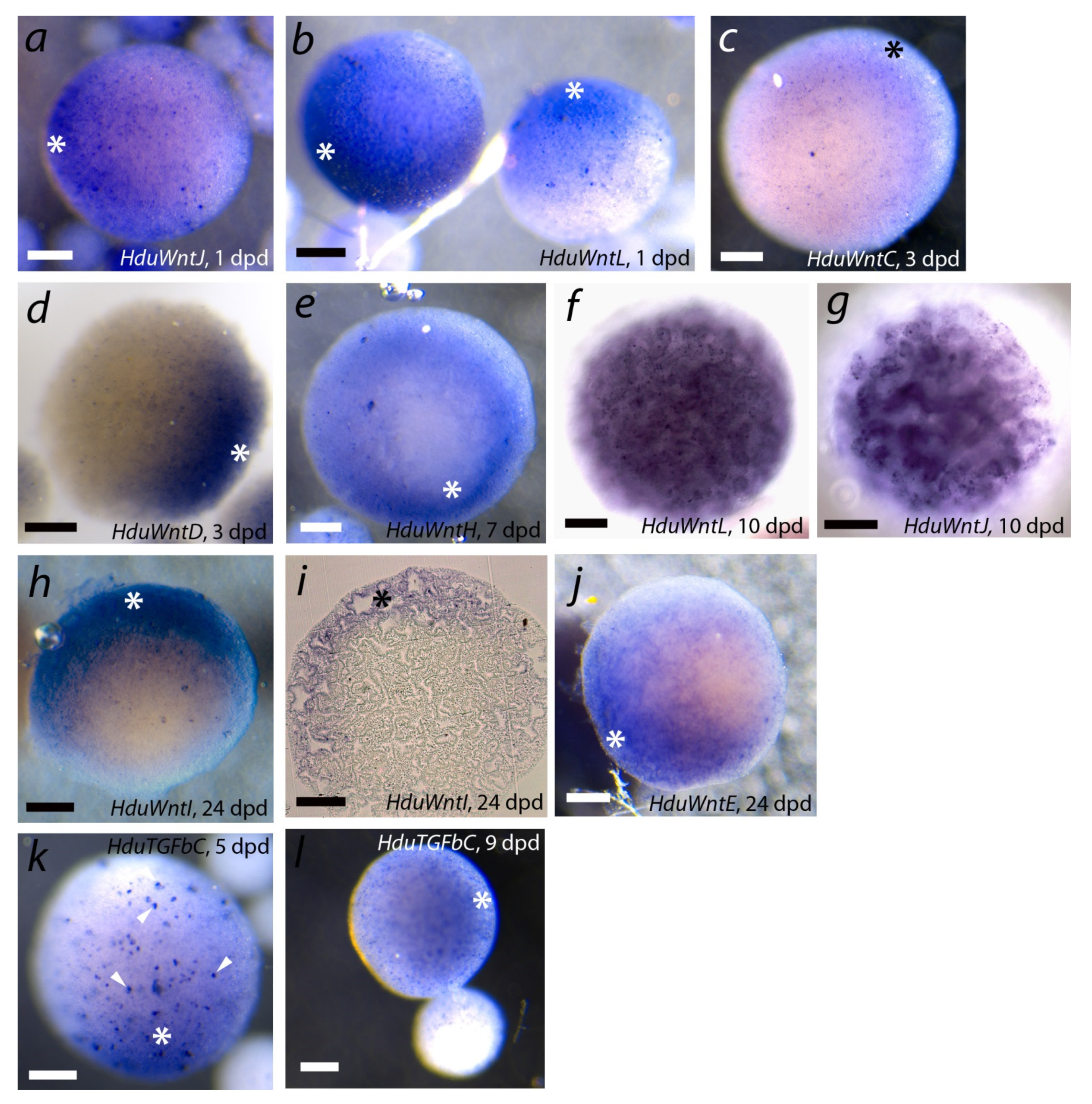
To determine the role of Wnt and TGF-beta ligands in the development of
H. dujardinii aggregates, we described the spatial patterns of their expression at different stages using WMISH. It was the first attempt to study spatial expression organization in primmorphs, and thus we focused on several key stages rather than checking all Wnt and TGF-beta genes at all stages of primmorph development. Stages 1, 3, 7, 10, and 24 dpd were investigated for
Wnt, 5 and 9 dpd, for
TGF-beta expression. Generally,
Wnt, in contrast to
TGF-beta, demonstrated a wide variety of expression patterns. However, we noted the heterogeneity of aggregates in the levels of Wnt and TGF-beta ligands’ expression. Cell aggregates of
H. dujardinii show considerable size variations from the earliest stages of cell reaggregation [
10]. We were able to show that large (700–800 µm) aggregates had diffuse or localized expression of most of the studied genes, while small ones (50–100 µm) most often did not show any signal. Therefore, further descriptions of expression patterns are given only for large aggregates.
Figure 8 represents some expression patterns demonstrating polarity in transcript localization. Diffuse patterns or absence of detectable expression are not shown for studied stages. We found asymmetric expression at different reaggregation stages for studied genes. Four genes (
HduWntC,
HduWntD,
HduWntJ, and
HduWntL) show asymmetric expression already during exopinacoderm formation at the ESP/TP stage (1–3 dpd) (
Figure 8a–d). ESPs and TPs show the expression along the gradient with a maximum at one pole of the aggregate. These genes lose the polar expression at later stages: at the PDP stage (at 10 dpd),
HduWntD is expressed diffusely, while
HduWntJ and
HduWntL are expressed in choanocytes of the emerging aquiferous system (
Figure 8f,g).
For
HduWntH and
HduWntI, expression was studied only at later stages of cell reaggregation: in PDPs 7 dpd and 24 dpd. For both genes at both studied stages, an asymmetric expression with a maximum at one pole of the aggregate was shown (
Figure 8e,h,i).
HduWntE has clearly asymmetric expression in the majority of studied PDPs at 24 dpd. This gene was expressed along most of the aggregate surface, except for one pole free of the transcript (
Figure 8j). Intriguingly, at the earlier DP stage (6 dpd), the
HduWntE transcript had uniformly high expression. Combined WMISH and RNA-seq data show that expression of
HduWntE is gradually restricted during aggregate development:
HduWntE show a high level of the expression in ESPs/TPs (1 dpd), then its expression level decreases in DPs (6 dpd), and, finally,
HduWntE become vanished from one pole of DPDs (24 dpd).
Seven out of the eight studied
TGF-beta demonstrated diffuse expression of varying intensity at all studied stages (1, 3, 6, 9, 12, and 20 dpd;
Supplementary File S8). In contrast,
HduTGFbC demonstrated a slight increase in expression at one pole of a primmorph at the DP (5 dpd) and PDP (9 dpd) stages.
Thus, we can state a dynamic pattern of asymmetric expression of individual Wnt and TGF-beta at different stages of H. dujardinii cell reaggregation.
4. Discussion
Both signaling mechanisms, Wnt and TGF-beta, are involved in the axis specification in a broad range of metazoan species. Most metazoans have at least one clearly defined axis in adulthood. The majority of sponges demonstrate a well-established morphological axis only at the stage of the swimming larva, and the involvement of the Wnt and TGF-beta ligands in embryonic development and larval patterning has been described only in
Sycon ciliatum and
Amphimedon queenslandica [
7,
8,
9]. In contrast, the adult sponge has no axis in the usual sense—as the imaginary line joining two ends/sides of the body. However, at least one polar structure could be distinguished in any adult sponge—the osculum, exhalant aperture of the aquiferous system. It differs from other tissues both in structure and by molecular profile. It was shown that the osculum, due to the Wnt cascade activity, organizes the aquiferous system, i.e., there is a molecular mechanism that supports the animal’s body plan [
13,
37,
38]. Moreover, cells constituting the osculum express
Wnt and
TGF-beta [
9,
13].
Sponges are capable of whole-body regeneration from dissociated cells into a functional adult animal in the course of cell reaggregation [
11]. The molecular interactions that support the body plan should be lost during the tissue dissociation procedure and then be reestablished during subsequent aggregate development. This assumption is supported by observed morphological transformations of developing aggregates, which, in particular, change their symmetry: asymmetric primary multicellular aggregates develop into radially symmetric primmorphs and then into polarized reconstructed sponges, bearing osculum [
10,
39]. Differential expression of Wnt and TGF-beta was described during reaggregation of
S. ciliatum cells by RNA-seq [
12]. Considering these facts, we assumed that Wnt and TGF-beta pathways’ activity might take place in the polarization (appearance of asymmetry) of the developing primmorphs in
Halisarca dujardinii.
The participants of Wnt and TGF-beta pathways in developmental processes have been previously described in another member of the class Demospongiae,
A. queenslandica, and the calcareous sponge
S. ciliatum. In the
Sycon, 21 Wnt and 22 TGF-beta proteins were found, which dispelled the idea of only two homologs of Wnt ligand in the last common ancestor of multicellular organisms [
8,
9]. We isolated 10 homologs of Wnt from the
H. dujardinii transcriptome and described dynamically expressed patterns in the adult sponge, larvae, during oogenesis and regeneration [
13]. To reveal if the cascades are able to function, we annotate the main components of Wnt and TGF-beta pathways
H. dujardinii transcriptome. Subsequent analysis of the involvement of Wnt and TGF-beta pathways in
H. dujardinii primmorph development was done by the evaluation of gene expression using RNA-seq and describing ligand expression patterns in primmorphs by WMISH. Data about gene activity were compared with ultrastructural observations on the reaggregation and primmorph development in
H. dujardinii.
The core components of the Wnt and TGF-beta signaling pathways in
H. dujardinii demonstrate a high conservation level of protein sequence and domain architecture. The presence of critical amino acid residues in Frizzled and LRP5/6 suggests that most of the known protein–protein interactions that exist in the bilaterian Wnt pathway appear to be possible in
H. dujardinii. Interestingly, Axin—the core protein of bCDC—was not identified in
H. dujardinii. Axin was also not found in the
S. ciliatum, although it is present in
A. queenslandica [
8,
9]. Meanwhile, LRP5/6, which is responsible for binding to Axin in the cytoplasm, has all known conserved domains, as well as five cytoplasmic PPPSPxS motifs phosphorylated by GSK3b and CK1 (
Figure 3a,c). Mutational analysis showed that substitution of serine to alanine in these motifs interferes with signal transduction into the cell, and LRP6, devoid of the extracellular domain, works as a constitutively active receptor. In addition, one motif out of five is sufficient for signal transduction [
27,
28,
29,
31,
40,
41]. It is the phosphorylated PPPSPxS motif that serves as the docking site for the cytoplasmic Axin [
29]. In Bilateria, Axin binds GSK3b, CK1, beta-catenin, APC, and Dishevelled; it acts as a scaffold for efficient phosphorylation of beta-catenin. Perhaps, in sponges, Axin homologs do not play such a key role as in Bilateria, and some other protein performs its functions. However, in this case, the role of conservative sites of interaction with it on LRP5/6 remains unclear. Obviously, the structure of the Wnt pathway in sponges lacking Axin requires further study from a biochemical perspective. Thus, despite the absence of one of the main components of bCDC, we found sites for its binding in other proteins.
Meanwhile, despite the completeness of the description of the bCDC participants and their interactions at the molecular level, much in the functioning of this protein complex remains disputable. Moreover, it was shown that adhesion junction proteins are co-immunoprecipitated together with beta-catenin in
Ephydatia muelleri but not participants of the Wnt pathway [
42]. In contrast, in mammalian cells, bCDC components are co-immunoprecipitated together or with beta-catenin [
43,
44]. However, it seems that beta-catenin probably retains the conservative role as a substrate for GSK3b, since inhibition of GSK3b leads to the disappearance of phosphorylated beta-catenin [
42].
Despite the presence of all conserved domains and critical amino acid residues, Frizzleds from sponges are closer to each other and orthologs from Ctenophora than to any of the described receptor families in Bilateria + Cnidaria. We can see that the Frizzleds’ sequences from sponges and comb jelly form a clade, sister to the Fzd/8 and Fzd1/2/7 branches. HduFzdA clusters into a subtree with two receptors from calcareous sponge S. ciliatum and ctenophore MlFzdB. HduFzdB is included in a compact group with sequences of other demosponges, A. queenslandica, Suberites domuncula, and E. muelleri. HduFzdC–HduFzdF, together with the proteins from A. queenslandica and E. muelleri, form a clade with ctenophoran MlFzdA. However, we must pay attention to the length of branches inside the clade of Porifera + Ctenophora sequences, considering the possible influence of long branch attraction. Based on this, we assume that Frizzled in sponges and ctenophores occurred independently of those of other multicellular organisms.
TGF-beta superfamily proteins together with Wnt participate in the specification of body axes and cell fates. These secreted pleiotropic factors play critical roles in embryogenesis and adult tissue homeostasis by regulating cell proliferation, differentiation, death, and migration in context-dependent manner. Mammal genomes contain 33 genes coding TGF-beta superfamily proteins from BMP, Activin, GDF, TGF-beta, and some other families. In
H. dujardinii, we identified eight TGF-beta proteins, three of which could be classified as TGF-beta sensu stricto, six receptors, and six SMAD proteins. Type I and Type II receptors in
H. dujardinii both demonstrate a high level of sequence divergence, although SMADs clearly fall into SMAD1/5/8, SMAD2/3, and SMAD4 families. Interestingly, representatives of SMAD6 or inhibitory SMAD were not found. SMAD proteins work in heteromeric complexes R-SMAD + Co-SMAD, and I-SMAD is a level of negative regulation of signaling activity. I-SMAD molecules are able to compete with Co-SMAD in binding with R-SMAD, not allowing the protein complex to activate the expression of target genes [
33].
RNA-seq analysis of pathway components demonstrates differences in expression level for at least seven ligands:
HduWntD, HduWntG,
HduWntH,
HduWntK, HduTGFbD,
HduTGFbF, and
HduTGFbG. Other components demonstrate different expression levels between paralogs as well. Basically, we observed similarities between replicas; however, for a few transcripts (
HduFzdB,
HduLRP5/6), striking differences between replicas of the same stage were observed. We suppose that this results from the heterogeneity of the primmorphs population, which usually shows a slightly different rate of development even within a single culture [
10,
45]. Nevertheless, 5 out of 18 ligands Wnt and TGF-beta demonstrate significant differences in expression level between the studied stages of cell reaggregation. Attention is drawn to the scatter of expression levels within the
Wnt and
TGF-beta: the map shows only relative values, making some transcripts seem close to the zero expression level. However, these transcripts are expressed during primmorph development, which is confirmed by the WMISH data.
Our first experiments of WMISH with probes to
Wnt and
TGF-beta demonstrate polar and gradient expression for several signaling molecules in primmorphs. Most of
TGF-beta express uniformly at high or low levels at studied stages from early-stage primmorphs to progressed developing primmorphs (1, 3, 6, 9, 12, and 20 days post dissociation), but
HduTGFbC demonstrates a gradient pattern of expression in progressed developing primmorphs. Uniform expression of several
Wnt genes was not documented in
Figure 7, although several
Wnts such as TGF-beta express uniformly at different stages and levels that are demonstrated by RNA-seq. We found broad gradient patterns for some
Wnt and
TGF-beta with stock at one primmorph pole already at 24 hpd. It differs from similar experiments with reaggregation of
Hydra cells where expression of
Wnt starts from a cluster of several cells [
46].




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}