
Mapping and Analysis of Swi5 and Sfr1 Phosphorylation Sites

 , , , , , and
, , , , , and 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain Construction and General S. pombe Methods
2.2. Microscopy
2.3. Spot Tests
2.4. Western Blot Analysis
2.5. Protein Purification and Analysis
3. Results and Discussion
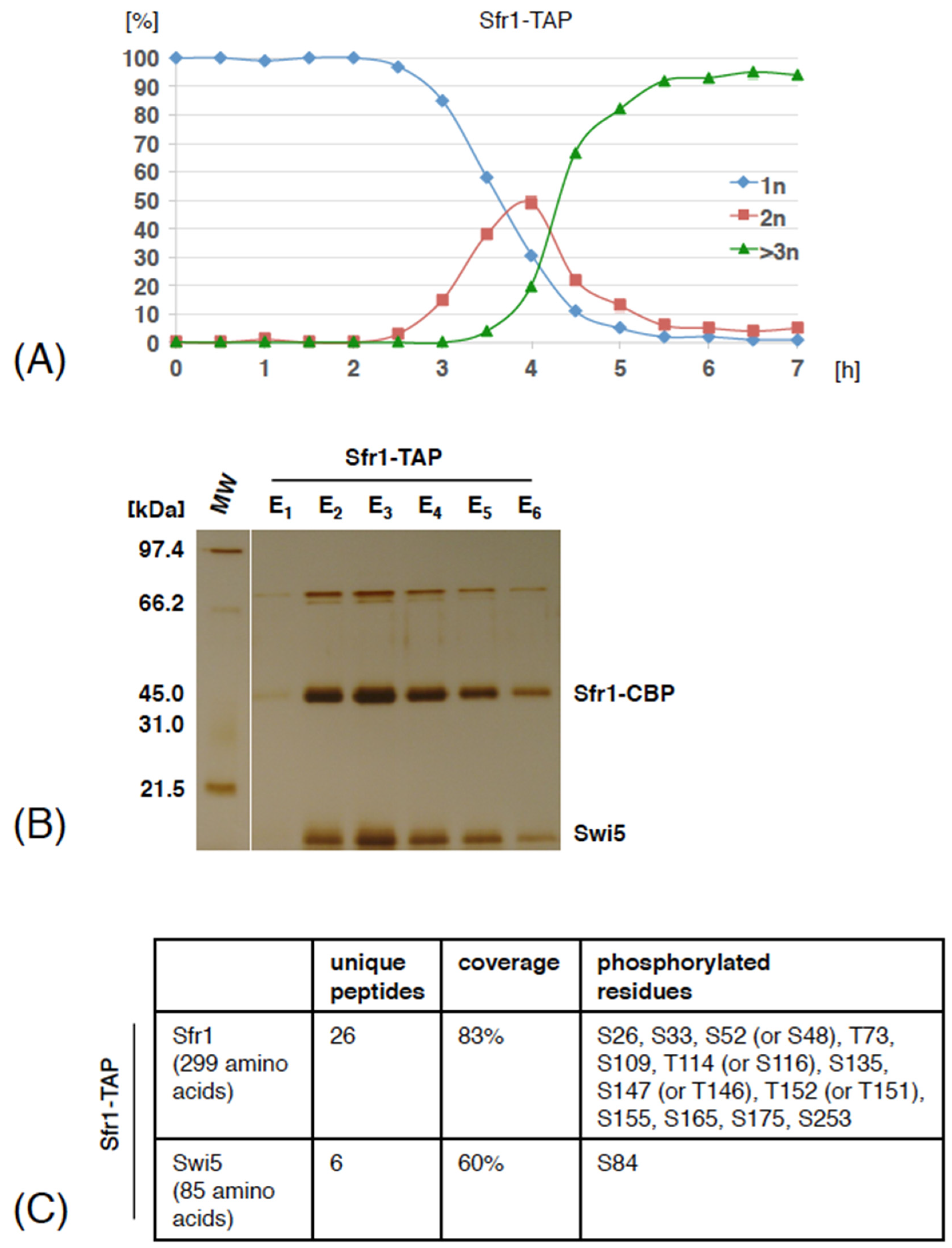
3.1. Swi5 and Sfr1 Are Phosphorylated during Meiosis
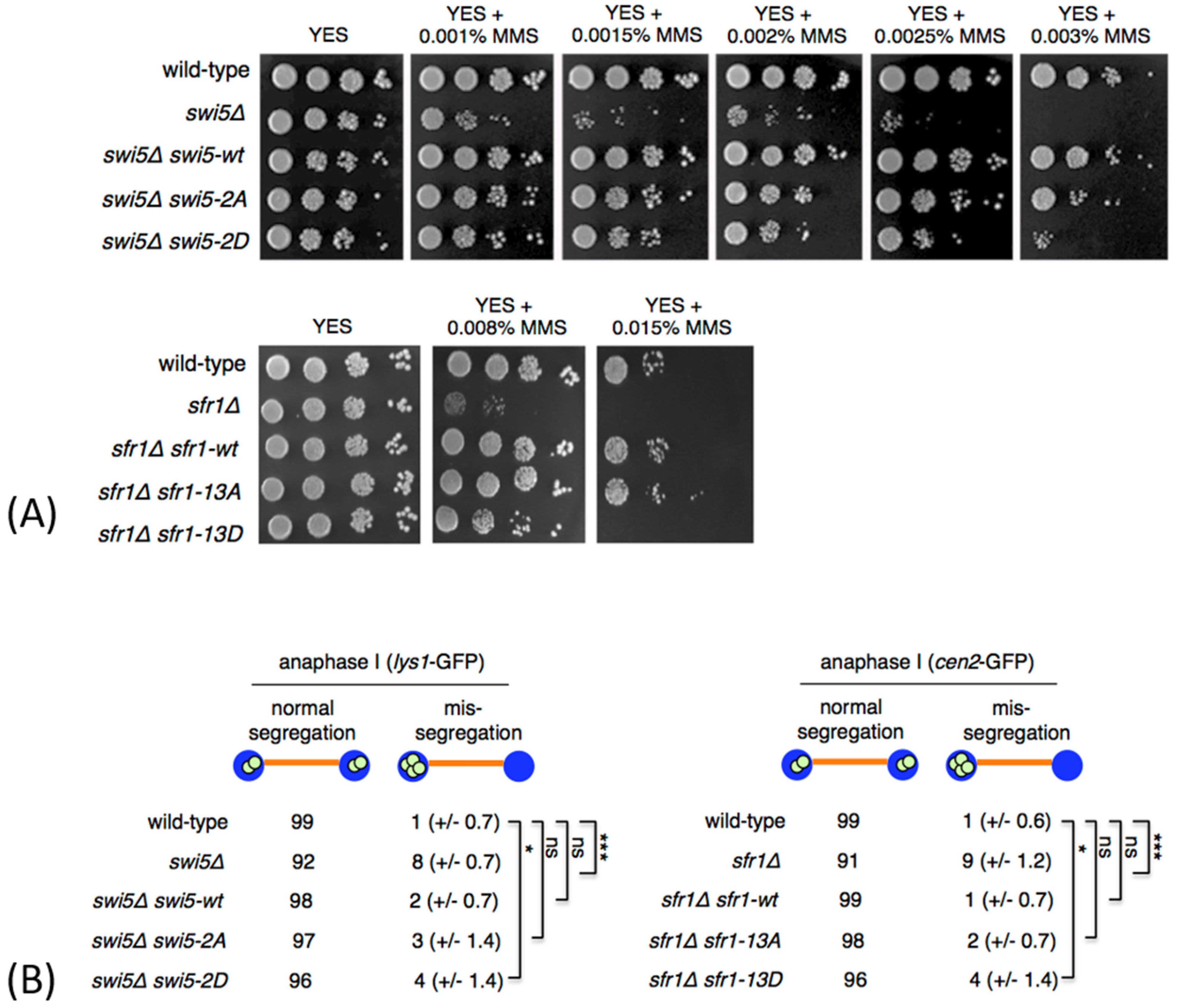
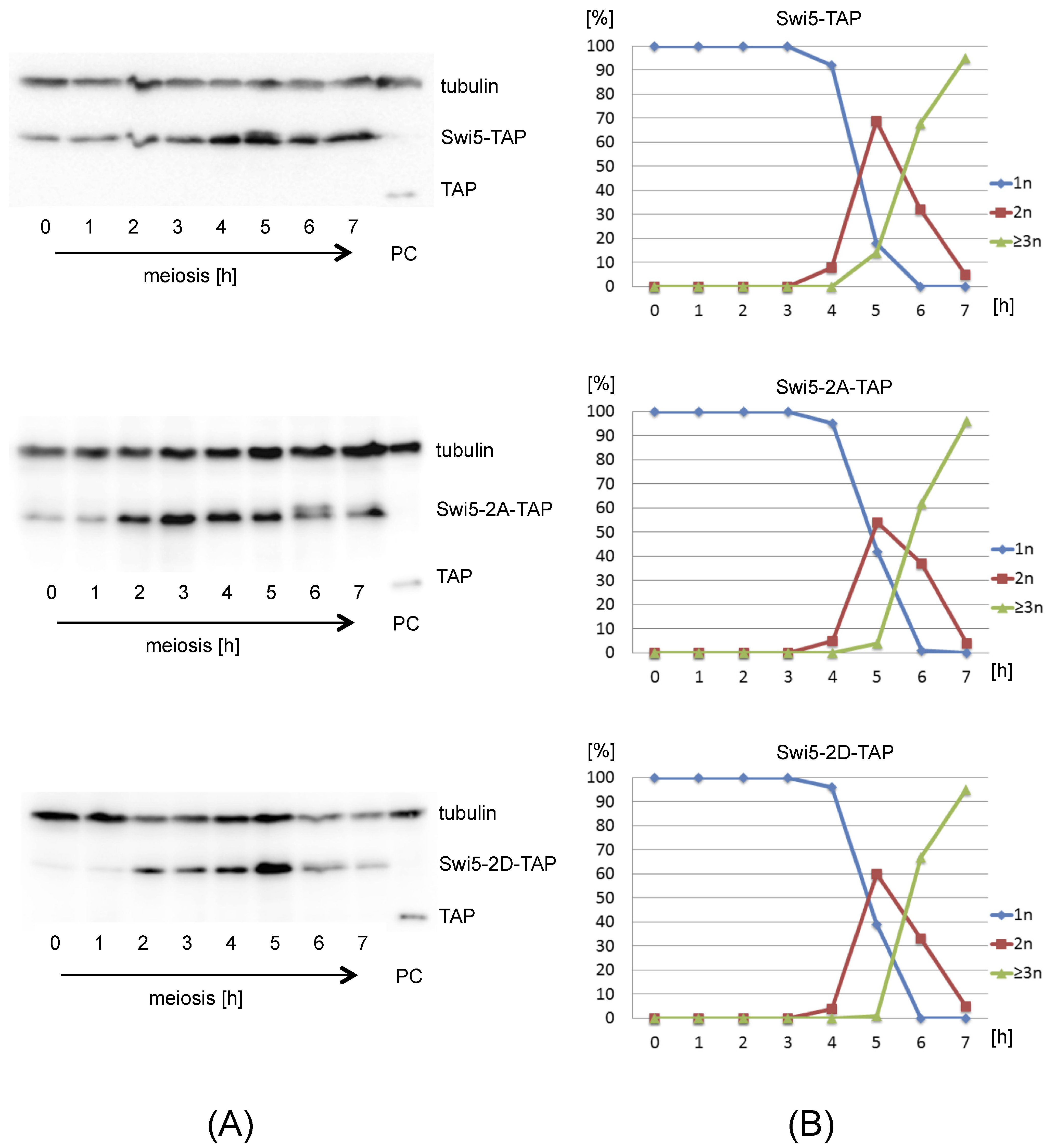
3.2. Phosphomimetic swi5 and sfr1 Mutants Are Not Fully Functional

Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SDS-PAGE | sodium dodecyl sulfate–polyacrylamide gel electrophoresis |
| YES | yeast extract with supplements medium |
| EMM | Edinburgh minimal medium |
| MMS | methyl-methanesulfonate |
| CBP | calmodulin binding peptide |
| TAP | tandem affinity purification |
References
- Jasin, M.; Rothstein, R. Repair of Strand Breaks by Homologous Recombination. Cold Spring Harb. Perspect. Biol. 2013, 5, a012740. [Google Scholar] [CrossRef]
- Ferrari, S.R.; Grubb, J.; Bishop, D.K. The Mei5-Sae3 Protein Complex Mediates Dmc1 Activity in Saccharomyces cerevisiae. J. Biol. Chem. 2009, 284, 11766–11770. [Google Scholar] [CrossRef] [PubMed]
- Akamatsu, Y.; Tsutsui, Y.; Morishita, T.; Siddique, M.S.; Kurokawa, Y.; Ikeguchi, M.; Yamao, F.; Arcangioli, B.; Iwasaki, H. Fission Yeast Swi5/Sfr1 and Rhp55/Rhp57 Differentially Regulate Rhp51-Dependent Recombination Outcomes. EMBO J. 2007, 26, 1352–1362. [Google Scholar] [CrossRef] [PubMed]
- Hyppa, R.W.; Smith, G.R. Crossover Invariance Determined by Partner Choice for Meiotic DNA Break Repair. Cell 2010, 142, 243–255. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lorenz, A.; Osman, F.; Sun, W.; Nandi, S.; Steinacher, R.; Whitby, M.C. The Fission Yeast FANCM Ortholog Directs Non-Crossover Recombination During Meiosis. Science 2012, 336, 1585–1588. [Google Scholar] [CrossRef]
- Argunhan, B.; Sakakura, M.; Afshar, N.; Kurihara, M.; Ito, K.; Maki, T.; Kanamaru, S.; Murayama, Y.; Tsubouchi, H.; Takahashi, M.; et al. Cooperative interactions facilitate stimulation of Rad51 by the Swi5-Sfr1 auxiliary factor complex. eLife 2020, 9. [Google Scholar] [CrossRef]
- Haruta, N.; Kurokawa, Y.; Murayama, Y.; Akamatsu, Y.; Unzai, S.; Tsutsui, Y.; Iwasaki, H. The Swi5–Sfr1 complex stimulates Rhp51/Rad51 and Dmc1-mediated DNA strand exchange in vitro. Nat. Struct. Mol. Biol. 2006, 13, 823–830. [Google Scholar] [CrossRef]
- Kuwabara, N.; Murayama, Y.; Hashimoto, H.; Kokabu, Y.; Ikeguchi, M.; Sato, M.; Mayanagi, K.; Tsutsui, Y.; Iwasaki, H.; Shimizu, T. Mechanistic Insights into the Activation of Rad51-Mediated Strand Exchange from the Structure of a Recombination Activator, the Swi5-Sfr1 Complex. Structure 2012, 20, 440–449. [Google Scholar] [CrossRef]
- Loidl, J. Conservation and Variability of Meiosis across the Eukaryotes. Annu. Rev. Genet. 2016, 50, 293–316. [Google Scholar] [CrossRef]
- Nambiar, M.; Chuang, Y.-C.; Smith, G.R. Distributing meiotic crossovers for optimal fertility and evolution. DNA Repair 2019, 81, 102648. [Google Scholar] [CrossRef]
- Akamatsu, Y.; Jasin, M. Role for the Mammalian Swi5-Sfr1 Complex in DNA Strand Break Repair through Homologous Recombination. PLoS Genet. 2010, 6, e1001160. [Google Scholar] [CrossRef] [PubMed]
- Khasanov, F.K.; Salakhova, A.F.; Khasanova, O.S.; Grishchuk, A.L.; Chepurnaja, O.V.; Korolev, V.G.; Kohli, J.; Bashkirov, V.I. Genetic analysis reveals different roles of Schizosaccharomyces pombe sfr1/dds20 in meiotic and mitotic DNA recombination and repair. Curr. Genet. 2008, 54, 197–211. [Google Scholar] [CrossRef][Green Version]
- Ellermeier, C.; Schmidt, H.; Smith, G.R. Swi5 Acts in Meiotic DNA Joint Molecule Formation in Schizosaccharomyces pombe. Genetics 2004, 168, 1891–1898. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Akamatsu, Y.; Dziadkowiec, D.; Ikeguchi, M.; Shinagawa, H.; Iwasaki, H. Two different Swi5-containing protein complexes are involved in mating-type switching and recombination repair in fission yeast. Proc. Natl. Acad. Sci. USA 2003, 100, 15770–15775. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Chen, J. The Role of the Human SWI5-MEI5 Complex in Homologous Recombination Repair. J. Biol. Chem. 2011, 286, 9888–9893. [Google Scholar] [CrossRef]
- Krejci, L.; Altmannova, V.; Spirek, M.; Zhao, X. Homologous recombination and its regulation. Nucleic Acids Res. 2012, 40, 5795–5818. [Google Scholar] [CrossRef] [PubMed]
- Phadnis, N.; Cipak, L.; Polakova, S.B.; Hyppa, R.W.; Cipakova, I.; Anrather, D.; Karvaiova, L.; Mechtler, K.; Smith, G.R.; Gregan, J. Casein Kinase 1 and Phosphorylation of Cohesin Subunit Rec11 (SA3) Promote Meiotic Recombination through Linear Element Formation. PLoS Genet. 2015, 11, e1005225. [Google Scholar] [CrossRef]
- Moreno, S.; Klar, A.; Nurse, P. Molecular genetic analysis of fission yeast Schizosaccharomyces pombe. Methods Enzymol. 1991, 194, 795–823. [Google Scholar] [CrossRef]
- Forsburg, S.L.; Rhind, N. Basic methods for fission yeast. Yeast 2006, 23, 173–183. [Google Scholar] [CrossRef]
- Gregan, J.; Rabitsch, P.K.; Rumpf, C.; Novatchkova, M.; Schleiffer, A.; Nasmyth, K. High-throughput knockout screen in fission yeast. Nat. Protoc. 2006, 1, 2457–2464. [Google Scholar] [CrossRef]
- Cipak, L.; Gupta, S.; Rajovic, I.; Jin, Q.-W.; Anrather, D.; Ammerer, G.; McCollum, D.; Gregan, J. Crosstalk between casein kinase II and Ste20-related kinase Nak1. Cell Cycle 2013, 12, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Rabitsch, K.P.; Gregan, J.; Schleiffer, A.; Javerzat, J.-P.; Eisenhaber, F.; Nasmyth, K. Two Fission Yeast Homologs of Drosophila Mei-S332 Are Required for Chromosome Segregation during Meiosis I and II. Curr. Biol. 2004, 14, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Cipak, L.; Hyppa, R.W.; Smith, G.R.; Gregan, J. ATP analog-sensitive Pat1 protein kinase for synchronous fission yeast meiosis at physiological temperature. Cell Cycle 2012, 11, 1626–1633. [Google Scholar] [CrossRef] [PubMed]
- Cipak, L.; Spirek, M.; Novatchkova, M.; Chen, Z.; Rumpf, C.; Lugmayr, W.; Mechtler, K.; Ammerer, G.; Csaszar, E.; Gregan, J. An Improved Strategy for Tandem Affinity Purification-Tagging of Schizosaccharomyces pombe Genes. Proteomics 2009, 9, 4825–4828. [Google Scholar] [CrossRef] [PubMed]
- Carpy, A.; Krug, K.; Graf, S.; Koch, A.; Popic, S.; Hauf, S.; Macek, B. Absolute Proteome and Phosphoproteome Dynamics during the Cell Cycle of Schizosaccharomyces pombe (Fission Yeast). Mol. Cell. Proteom. 2014, 13, 1925–1936. [Google Scholar] [CrossRef] [PubMed]
- Sivakova, B.; Jurcik, J.; Lukacova, V.; Selicky, T.; Cipakova, I.; Barath, P.; Cipak, L. Label-Free Quantitative Phosphoproteomics of the Fission Yeast Schizosaccharomyces pombe Using Strong Anion Exchange- and Porous Graphitic Carbon-Based Fractionation Strategies. Int. J. Mol. Sci. 2021, 22, 1747. [Google Scholar] [CrossRef]
- Koch, A.; Krug, K.; Pengelley, S.; Macek, B.; Hauf, S. Mitotic Substrates of the Kinase Aurora with Roles in Chromatin Regulation Identified Through Quantitative Phosphoproteomics of Fission Yeast. Sci. Signal. 2011, 4, rs6. [Google Scholar] [CrossRef]
- Swaffer, M.P.; Jones, A.W.; Flynn, H.R.; Snijders, B.; Nurse, P. Quantitative Phosphoproteomics Reveals the Signaling Dynamics of Cell-Cycle Kinases in the Fission Yeast Schizosaccharomyces pombe. Cell Rep. 2018, 24, 503–514. [Google Scholar] [CrossRef]
- Kettenbach, A.N.; Deng, L.; Wu, Y.; Baldissard, S.; Adamo, M.E.; Gerber, S.; Moseley, J.B. Quantitative Phosphoproteomics Reveals Pathways for Coordination of Cell Growth and Division by the Conserved Fission Yeast Kinase Pom1*. Mol. Cell. Proteom. 2015, 14, 1275–1287. [Google Scholar] [CrossRef]
- Wyatt, M.D.; Pittman, D.L. Methylating Agents and DNA Repair Responses: Methylated Bases and Sources of Strand Breaks. Chem. Res. Toxicol. 2006, 19, 1580–1594. [Google Scholar] [CrossRef]
- Martín-Castellanos, C.; Blanco, M.; Rozalén, A.E.; Pérez-Hidalgo, L.; García, A.I.; Conde, F.M.; Mata, J.; Ellermeier, C.; Davis, L.; San-Segundo, P.; et al. A Large-Scale Screen in S. pombe Identifies Seven Novel Genes Required for Critical Meiotic Events. Curr. Biol. 2005, 15, 2056–2062. [Google Scholar] [CrossRef] [PubMed]
- Nabeshima, K.; Nakagawa, T.; Straight, A.F.; Murray, A.; Chikashige, Y.; Yamashita, Y.M.; Hiraoka, Y.; Yanagida, M. Dynamics of Centromeres during Metaphase–Anaphase Transition in Fission Yeast: Dis1 Is Implicated in Force Balance in Metaphase Bipolar Spindle. Mol. Biol. Cell 1998, 9, 3211–3225. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.; Hiraoka, Y. Monopolar spindle attachment of sister chromatids is ensured by two distinct mechanisms at the first meiotic division in fission yeast. EMBO J. 2003, 22, 2284–2296. [Google Scholar] [CrossRef] [PubMed]
- Mata, J.; Lyne, R.; Burns, G.; Bahler, J. The transcriptional program of meiosis and sporulation in fission yeast. Nat. Genet. 2002, 32, 143–147. [Google Scholar] [CrossRef]
- Dephoure, N.; Gould, K.; Gygi, S.P.; Kellogg, D.R. Mapping and analysis of phosphorylation sites: A quick guide for cell biologists. Mol. Biol. Cell 2013, 24, 535–542. [Google Scholar] [CrossRef]
- Huraiova, B.; Kanovits, J.; Polakova, S.B.; Cipak, L.; Benko, Z.; Sevcovicova, A.; Anrather, D.; Ammerer, G.; Duncan, C.D.S.; Mata, J.; et al. Proteomic Analysis of Meiosis and Characterization of Novel Short Open Reading Frames in the Fission Yeastschizosaccharomyces pombe. Cell Cycle 2020, 19, 1777–1785. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sevcovicova, A.; Plava, J.; Gazdarica, M.; Szabova, E.; Huraiova, B.; Gaplovska-Kysela, K.; Cipakova, I.; Cipak, L.; Gregan, J. Mapping and Analysis of Swi5 and Sfr1 Phosphorylation Sites. Genes 2021, 12, 1014. https://doi.org/10.3390/genes12071014
Sevcovicova A, Plava J, Gazdarica M, Szabova E, Huraiova B, Gaplovska-Kysela K, Cipakova I, Cipak L, Gregan J. Mapping and Analysis of Swi5 and Sfr1 Phosphorylation Sites. Genes. 2021; 12(7):1014. https://doi.org/10.3390/genes12071014
Chicago/Turabian StyleSevcovicova, Andrea, Jana Plava, Matej Gazdarica, Eva Szabova, Barbora Huraiova, Katarina Gaplovska-Kysela, Ingrid Cipakova, Lubos Cipak, and Juraj Gregan. 2021. "Mapping and Analysis of Swi5 and Sfr1 Phosphorylation Sites" Genes 12, no. 7: 1014. https://doi.org/10.3390/genes12071014
APA StyleSevcovicova, A., Plava, J., Gazdarica, M., Szabova, E., Huraiova, B., Gaplovska-Kysela, K., Cipakova, I., Cipak, L., & Gregan, J. (2021). Mapping and Analysis of Swi5 and Sfr1 Phosphorylation Sites. Genes, 12(7), 1014. https://doi.org/10.3390/genes12071014