
Autophosphorylation and Self-Activation of DNA-Dependent Protein Kinase


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
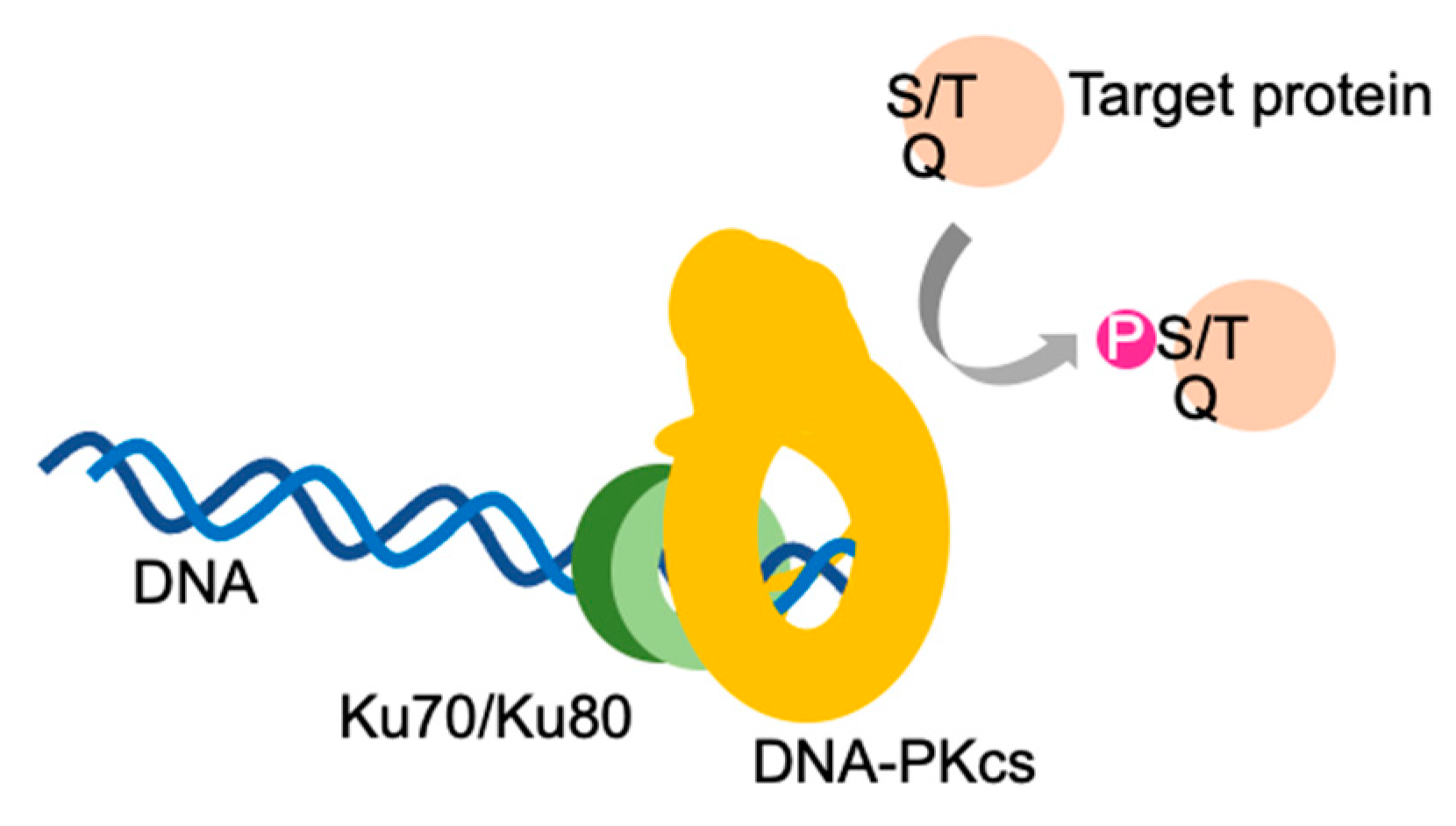
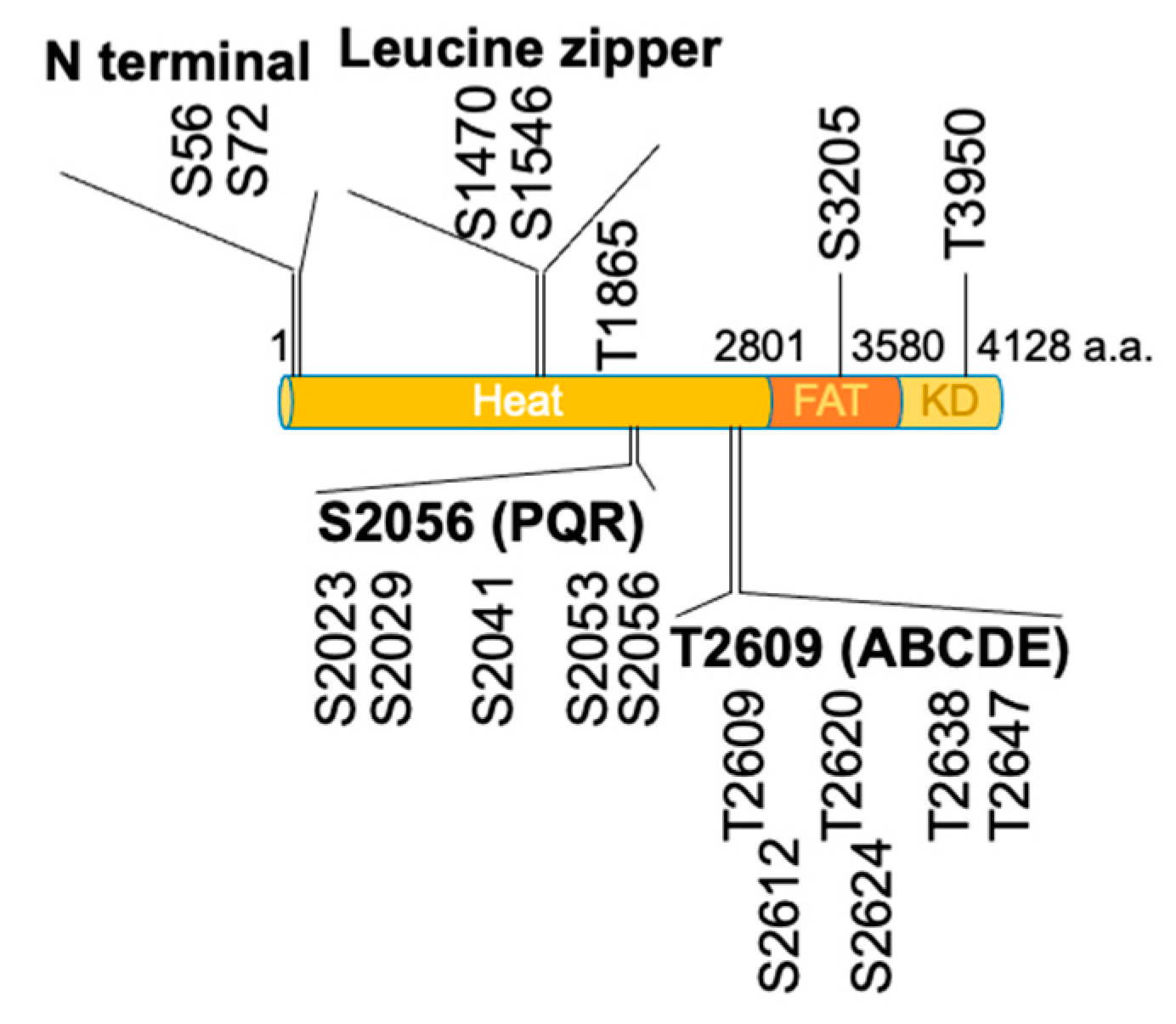
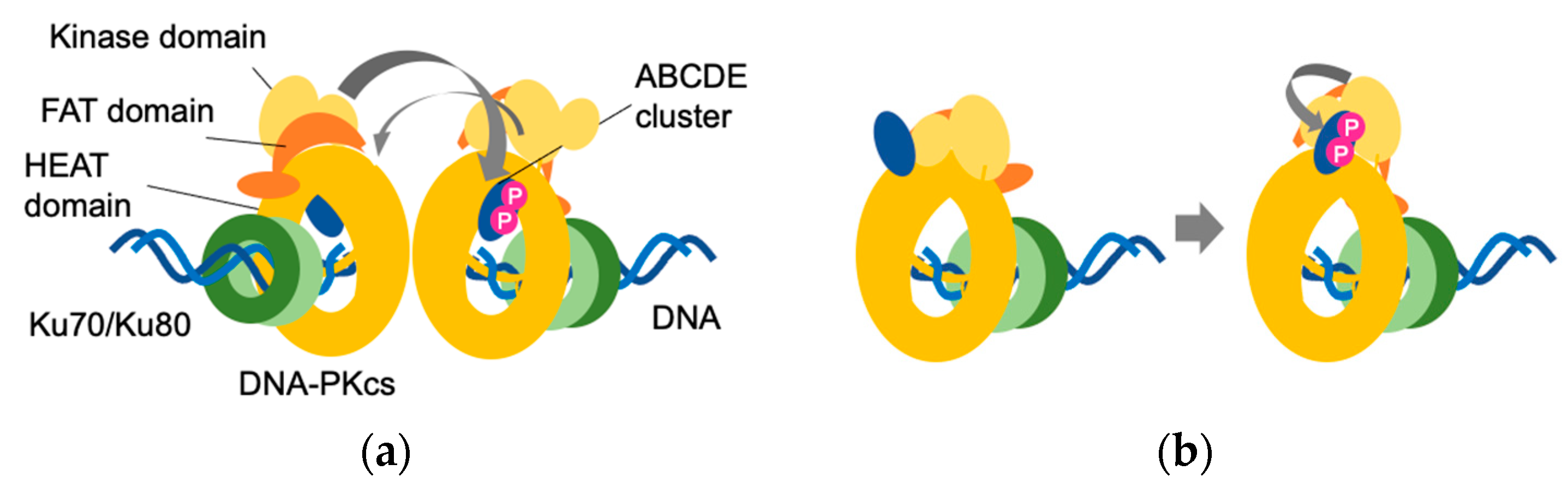
2. Self-Activation of a DNA-Dependent Protein Kinase Catalytic Subunit
3. Effects of DNA-PKcs on Development and Differentiation Determined by Comparing Knockout and Mutant Mice
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, R.X.; Zhou, P.K. DNA damage response signaling pathways and targets for radiotherapy sensitization in cancer. Signal Transduct. Target. Ther. 2020, 5, 60. [Google Scholar] [CrossRef]
- Jeggo, P.; Löbrich, M. Radiation-induced DNA damage responses. Radiat. Prot. Dosim. 2006, 122, 124–127. [Google Scholar] [CrossRef]
- Lanz, M.C.; Dibitetto, D.; Smolka, M.B. DNA damage kinase signaling: Checkpoint and repair at 30 years. EMBO J. 2019, 38, e101801. [Google Scholar] [CrossRef]
- Kidiyoor, G.R.; Kumar, A.; Foiani, M. ATR-mediated regulation of nuclear and cellular plasticity. DNA Repair 2016, 44, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackford, A.N.; Jackson, S.P. ATM, ATR, and DNA-PK: The Trinity at the heart of the DNA damage response. Mol. Cell 2017, 66, 801–817. [Google Scholar] [CrossRef] [Green Version]
- Elias-Villalobos, A.; Fort, P.; Helmlinger, D. New insights into the evolutionary conservation of the sole PIKK pseudokinase Tra1/TRRAP. Biochem. Soc. Trans. 2019, 47, 1597–1608. [Google Scholar] [CrossRef] [PubMed]
- Imseng, S.; Aylett, C.H.; Maier, T. Architecture and activation of phosphatidylinositol 3-kinase related kinases. Curr. Opin. Struct. Biol. 2018, 49, 177–189. [Google Scholar] [CrossRef]
- Lees-Miller, J.P.; Cobban, A.; Katsonis, P.; Bacolla, A.; Tsutakawa, S.E.; Hammel, M.; Meek, K.; Anderson, D.W.; Lichtarge, O.; Tainer, J.A.; et al. Uncovering DNA-PKcs ancient phylogeny, unique sequence motifs and insights for human disease. Prog. Biophys. Mol. Biol. 2021, 163, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Ensminger, M.; Löbrich, M. One end to rule them all: Non-homologous end-joining and homologous recombination at DNA double-strand breaks. Br. J. Radiol. 2020, 93, 20191054. [Google Scholar] [CrossRef]
- Ghosh, D.; Raghavan, S.C. Nonhomologous end joining: New accessory factors fine tune the machinery. Trends Genet. 2021, 37, 582–599. [Google Scholar] [CrossRef]
- Kurosawa, A.; Adachi, N. Functions and regulation of Artemis: A goddess in the maintenance of genome integrity. J. Radiat. Res. 2010, 51, 503–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodwin, J.F.; Knudsen, K.E. Beyond DNA repair: DNA-PK function in cancer. Cancer Discov. 2014, 4, 1126–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.H.Y.; Watanabe, G.; Gerodimos, C.A.; Ochi, T.; Blundell, T.L.; Jackson, S.P.; Lieber, M.R. Different DNA end configurations dictate which NHEJ components are most important for joining efficiency. J. Biol. Chem. 2016, 291, 24377–24389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, D.; Hayashihara, K.; Shima, H.; Higashide, M.; Terasawa, M.; Gasser, S.M.; Shinohara, M. The MRX complex ensures NHEJ fidelity through multiple pathways including Xrs2-FHA-dependent Tel1 activation. PLoS Genet. 2016, 12, e1005942. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, J.; Zhou, G.; Lajeunesse, M.; Le, N.; Stawicki, B.N.; Corcino, Y.L.; Berkner, K.L.; Runge, K.W. Nonhomologous end-joining with minimal sequence loss is promoted by the Mre11-Rad50-Nbs1-Ctp1 complex in Schizosaccharomyces pombe. Genetics 2017, 206, 481–496. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, K.; Shinohara, A.; Shinohara, M. Forkhead-associated domain of yeast Xrs2, a homolog of human Nbs1, promotes nonhomologous end joining through interaction with a ligase IV partner protein, Lif1. Genetics 2008, 179, 213–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochi, T.; Blackford, A.N.; Coates, J.; Jhujh, S.; Mehmood, S.; Tamura, N.; Travers, J.; Wu, Q.; Draviam, V.M.; Robinson, C.V.; et al. DNA repair. PAXX, a paralog of XRCC4 and XLF, interacts with Ku to promote DNA double-strand break repair. Science 2015, 347, 185–188. [Google Scholar] [CrossRef] [Green Version]
- Xing, M.; Yang, M.; Huo, W.; Feng, F.; Wei, L.; Jiang, W.; Ning, S.; Yan, Z.; Li, W.; Wang, Q.; et al. Interactome analysis identifies a new paralogue of XRCC4 in non-homologous end joining DNA repair pathway. Nat. Commun. 2015, 6, 6233. [Google Scholar] [CrossRef] [Green Version]
- Andres, S.N.; Vergnes, A.; Ristic, D.; Wyman, C.; Modesti, M.; Junop, M. A human XRCC4-XLF complex bridges DNA. Nucleic Acids Res. 2012, 40, 1868–1878. [Google Scholar] [CrossRef] [Green Version]
- Hammel, M.; Yu, Y.; Fang, S.; Lees-Miller, S.P.; Tainer, J.A. XLF regulates filament architecture of the XRCC4.ligase IV complex. Structure 2010, 18, 1431–1442. [Google Scholar] [CrossRef] [Green Version]
- Mahaney, B.L.; Hammel, M.; Meek, K.; Tainer, J.A.; Lees-Miller, S.P. XRCC4 and XLF form long helical protein filaments suitable for DNA end protection and alignment to facilitate DNA double strand break repair. Biochem. Cell Biol. 2013, 91, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Ropars, V.; Drevet, P.; Legrand, P.; Baconnais, S.; Amram, J.; Faure, G.; Marquez, J.A.; Pietrement, O.; Guerois, R.; Callebaut, I.; et al. Structural characterization of filaments formed by human Xrcc4-Cernunnos/XLF complex involved in nonhomologous DNA end-joining. Proc. Natl. Acad. Sci. USA 2011, 108, 12663–12668. [Google Scholar] [CrossRef] [Green Version]
- De Villartay, J.P. Congenital defects in V(D)J recombination. Br. Med. Bull. 2015, 114, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Lieber, M.R. Binding of inositol hexakisphosphate (IP6) to Ku but not to DNA-PKcs. J. Biol. Chem. 2002, 277, 10756–10759. [Google Scholar] [CrossRef] [Green Version]
- Anisenko, A.N.; Gottikh, M.B. Role of cellular DNA repair systems in HIV-1 replication. Mol. Biol. 2019, 53, 355–366. [Google Scholar] [CrossRef]
- Cui, X.; Yu, Y.; Gupta, S.; Cho, Y.M.; Lees-Miller, S.P.; Meek, K. Autophosphorylation of DNA-dependent protein kinase regulates DNA end processing and may also alter double-strand break repair pathway choice. Mol. Cell. Biol. 2005, 25, 10842–10852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uematsu, N.; Weterings, E.; Yano, K.; Morotomi-Yano, K.; Jakob, B.; Taucher-Scholz, G.; Mari, P.O.; van Gent, D.C.; Chen, B.P.; Chen, D.J. Autophosphorylation of DNA-PKCS regulates its dynamics at DNA double-strand breaks. J. Cell Biol. 2007, 177, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jette, N.; Lees-Miller, S.P. The DNA-dependent protein kinase: A multifunctional protein kinase with roles in DNA double strand break repair and mitosis. Prog. Biophys. Mol. Biol. 2015, 117, 194–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.J.; Lin, Y.F.; Chou, H.Y.; Yajima, H.; Fattah, K.R.; Lee, S.C.; Chen, B.P. Involvement of DNA-dependent protein kinase in normal cell cycle progression through mitosis. J. Biol. Chem. 2011, 286, 12796–12802. [Google Scholar] [CrossRef] [Green Version]
- Block, W.D.; Yu, Y.; Merkle, D.; Gifford, J.L.; Ding, Q.; Meek, K.; Lees-Miller, S.P. Autophosphorylation-dependent remodeling of the DNA-dependent protein kinase catalytic subunit regulates ligation of DNA ends. Nucleic Acids Res. 2004, 32, 4351–4357. [Google Scholar] [CrossRef] [Green Version]
- Crowe, J.L.; Wang, X.S.; Shao, Z.; Lee, B.J.; Estes, V.M.; Zha, S. DNA-PKcs phosphorylation at the T2609 cluster alters the repair pathway choice during immunoglobulin class switch recombination. Proc. Natl. Acad. Sci. USA 2020, 117, 22953–22961. [Google Scholar] [CrossRef]
- Ding, Q.; Reddy, Y.V.; Wang, W.; Woods, T.; Douglas, P.; Ramsden, D.A.; Lees-Miller, S.P.; Meek, K. Autophosphorylation of the catalytic subunit of the DNA-dependent protein kinase is required for efficient end processing during DNA double-strand break repair. Mol. Cell. Biol. 2003, 23, 5836–5848. [Google Scholar] [CrossRef] [Green Version]
- Meek, K. Activation of DNA-PK by hairpinned DNA ends reveals a stepwise mechanism of kinase activation. Nucleic Acids Res. 2020, 48, 9098–9108. [Google Scholar] [CrossRef]
- Shao, Z.; Flynn, R.A.; Crowe, J.L.; Zhu, Y.; Liang, J.; Jiang, W.; Aryan, F.; Aoude, P.; Bertozzi, C.R.; Estes, V.M.; et al. DNA-PKcs has KU-dependent function in rRNA processing and haematopoiesis. Nature 2020, 579, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yajima, H.; Huynh, H.; Zheng, J.; Callen, E.; Chen, H.T.; Wong, N.; Bunting, S.; Lin, Y.F.; Li, M.; et al. Congenital bone marrow failure in DNA-PKcs mutant mice associated with deficiencies in DNA repair. J. Cell. Biol. 2011, 193, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.R.; Corpina, R.A.; Goldberg, J. Structure of the Ku heterodimer bound to DNA and its implications for double-strand break repair. Nature 2001, 412, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Y.; Cary, R.B.; Chen, D.J.; Peterson, S.R.; Stewart, P.L. Cryo-EM imaging of the catalytic subunit of the DNA-dependent protein kinase. J. Mol. Biol. 1998, 284, 1075–1081. [Google Scholar] [CrossRef]
- Leuther, K.K.; Hammarsten, O.; Kornberg, R.D.; Chu, G. Structure of DNA-dependent protein kinase: Implications for its regulation by DNA. EMBO J. 1999, 18, 1114–1123. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Calzada, A.; Maman, J.D.; Spagnolo, L.; Pearl, L.H.; Llorca, O. Three-dimensional structure and regulation of the DNA-dependent protein kinase catalytic subunit (DNA-PKcs). Structure 2005, 13, 243–255. [Google Scholar] [CrossRef]
- Chen, X.; Xu, X.; Chen, Y.; Cheung, J.C.; Wang, H.; Jiang, J.; de Val, N.; Fox, T.; Gellert, M.; Yang, W. Structure of an activated DNA-PK and its implications for NHEJ. Mol. Cell 2021, 81, 801–810 e803. [Google Scholar] [CrossRef]
- Sharif, H.; Li, Y.; Dong, Y.; Dong, L.; Wang, W.L.; Mao, Y.; Wu, H. Cryo-EM structure of the DNA-PK holoenzyme. Proc. Natl. Acad. Sci. USA 2017, 114, 7367–7372. [Google Scholar] [CrossRef] [Green Version]
- Sibanda, B.L.; Chirgadze, D.Y.; Ascher, D.B.; Blundell, T.L. DNA-PKcs structure suggests an allosteric mechanism modulating DNA double-strand break repair. Science 2017, 355, 520–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.; Liu, M.; Tian, Y.; Wang, J.; Xu, Y. Cryo-EM structure of human DNA-PK holoenzyme. Cell Res. 2017, 27, 1341–1350. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.W.; Lees-Miller, S.P. The nuclear serine/threonine protein kinase DNA-PK. Crit. Rev. Eukaryot. Gene. Expr. 1992, 2, 283–314. [Google Scholar] [PubMed]
- Cary, R.B.; Peterson, S.R.; Wang, J.; Bear, D.G.; Bradbury, E.M.; Chen, D.J. DNA looping by Ku and the DNA-dependent protein kinase. Proc. Natl. Acad. Sci. USA 1997, 94, 4267–4272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, R.B.; Yaneva, M.; Lieber, M.R. Productive and nonproductive complexes of Ku and DNA-dependent protein kinase at DNA termini. Mol. Cell. Biol. 1998, 18, 5908–5920. [Google Scholar] [CrossRef] [Green Version]
- Hanakahi, L.A.; Bartlet-Jones, M.; Chappell, C.; Pappin, D.; West, S.C. Binding of inositol phosphate to DNA-PK and stimulation of double-strand break repair. Cell 2000, 102, 721–729. [Google Scholar] [CrossRef] [Green Version]
- Hanakahi, L.A.; West, S.C. Specific interaction of IP6 with human Ku70/80, the DNA-binding subunit of DNA-PK. EMBO J. 2002, 21, 2038–2044. [Google Scholar] [CrossRef] [Green Version]
- Gat, Y.; Schuller, J.M.; Lingaraju, M.; Weyher, E.; Bonneau, F.; Strauss, M.; Murray, P.J.; Conti, E. InsP6 binding to PIKK kinases revealed by the cryo-EM structure of an SMG1-SMG8-SMG9 complex. Nat. Struct. Mol. Biol. 2019, 26, 1089–1093. [Google Scholar] [CrossRef]
- Chen, B.P.; Chan, D.W.; Kobayashi, J.; Burma, S.; Asaithamby, A.; Morotomi-Yano, K.; Botvinick, E.; Qin, J.; Chen, D.J. Cell cycle dependence of DNA-dependent protein kinase phosphorylation in response to DNA double strand breaks. J. Biol. Chem. 2005, 280, 14709–14715. [Google Scholar] [CrossRef] [Green Version]
- Lafont, F.; Fleury, F.; Benhelli-Mokrani, H. DNA-PKcs Ser2056 auto-phosphorylation is affected by an O-GlcNAcylation/phosphorylation interplay. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129705. [Google Scholar] [CrossRef]
- Toulany, M.; Lee, K.J.; Fattah, K.R.; Lin, Y.F.; Fehrenbacher, B.; Schaller, M.; Chen, B.P.; Chen, D.J.; Rodemann, H.P. Akt promotes post-irradiation survival of human tumor cells through initiation, progression, and termination of DNA-PKcs-dependent DNA double-strand break repair. Mol. Cancer Res. 2012, 10, 945–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Estes, V.M.; Wang, X.S.; Shao, Z.; Lee, B.J.; Lin, X.; Crowe, J.L.; Zha, S. Phosphorylation at S2053 in murine (S2056 in Human) DNA-PKcs is dispensable for lymphocyte development and class switch recombination. J. Immunol. 2019, 203, 178–187. [Google Scholar] [CrossRef]
- Reddy, Y.V.; Ding, Q.; Lees-Miller, S.P.; Meek, K.; Ramsden, D.A. Non-homologous end joining requires that the DNA-PK complex undergo an autophosphorylation-dependent rearrangement at DNA ends. J. Biol. Chem. 2004, 279, 39408–39413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neal, J.A.; Xu, Y.; Abe, M.; Hendrickson, E.; Meek, K. Restoration of ATM expression in DNA-PKcs-deficient cells inhibits signal end joining. J. Immunol. 2016, 196, 3032–3042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neal, J.A.; Dang, V.; Douglas, P.; Wold, M.S.; Lees-Miller, S.P.; Meek, K. Inhibition of homologous recombination by DNA-dependent protein kinase requires kinase activity, is titratable, and is modulated by autophosphorylation. Mol. Cell. Biol. 2011, 31, 1719–1733. [Google Scholar] [CrossRef] [Green Version]
- Neal, J.A.; Sugiman-Marangos, S.; VanderVere-Carozza, P.; Wagner, M.; Turchi, J.; Lees-Miller, S.P.; Junop, M.S.; Meek, K. Unraveling the complexities of DNA-dependent protein kinase autophosphorylation. Mol. Cell. Biol. 2014, 34, 2162–2175. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Chaudhuri, J.; Zhu, C.; Davidson, L.; Weaver, D.T.; Alt, F.W. A targeted DNA-PKcs-null mutation reveals DNA-PK-independent functions for KU in V(D)J recombination. Immunity 1998, 9, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Crowe, J.L.; Liu, X.; Nakajima, S.; Wang, Y.; Li, C.; Lee, B.J.; Dubois, R.L.; Liu, C.; Yu, X.; et al. Differential phosphorylation of DNA-PKcs regulates the interplay between end-processing and end-ligation during nonhomologous end-joining. Mol. Cell 2015, 58, 172–185. [Google Scholar] [CrossRef] [Green Version]
- Taccioli, G.E.; Amatucci, A.G.; Beamish, H.J.; Gell, D.; Xiang, X.H.; Torres Arzayus, M.I.; Priestley, A.; Jackson, S.P.; Marshak Rothstein, A.; Jeggo, P.A.; et al. Targeted disruption of the catalytic subunit of the DNA-PK gene in mice confers severe combined immunodeficiency and radiosensitivity. Immunity 1998, 9, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Matsunaga, S.; Lin, Y.F.; Sishc, B.; Shang, Z.; Sui, J.; Shih, H.Y.; Zhao, Y.; Foreman, O.; Story, M.D.; et al. Spontaneous tumor development in bone marrow-rescued DNA-PKcs(3A/3A) mice due to dysfunction of telomere leading strand deprotection. Oncogene 2016, 35, 3909–3918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, K.M.; Sharpless, N.E.; Gao, Y.; Sekiguchi, J.M.; Ferguson, D.O.; Zhu, C.; Manis, J.P.; Horner, J.; DePinho, R.A.; Alt, F.W. DNA ligase IV deficiency in mice leads to defective neurogenesis and embryonic lethality via the p53 pathway. Mol. Cell 2000, 5, 993–1002. [Google Scholar] [CrossRef]
- Nacht, M.; Strasser, A.; Chan, Y.R.; Harris, A.W.; Schlissel, M.; Bronson, R.T.; Jacks, T. Mutations in the p53 and SCID genes cooperate in tumorigenesis. Genes Dev. 1996, 10, 2055–2066. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Mills, K.D.; Ferguson, D.O.; Lee, C.; Manis, J.; Fleming, J.; Gao, Y.; Morton, C.C.; Alt, F.W. Unrepaired DNA breaks in p53-deficient cells lead to oncogenic gene amplification subsequent to translocations. Cell 2002, 109, 811–821. [Google Scholar] [CrossRef] [Green Version]
- Shadrina, O.; Garanina, I.; Korolev, S.; Zatsepin, T.; Van Assche, J.; Daouad, F.; Wallet, C.; Rohr, O.; Gottikh, M. Analysis of RNA binding properties of human Ku protein reveals its interactions with 7SK snRNA and protein components of 7SK snRNP complex. Biochimie 2020, 171–172, 110–123. [Google Scholar] [CrossRef]
- Yoo, S.; Dynan, W.S. Characterization of the RNA binding properties of Ku protein. Biochemistry 1998, 37, 1336–1343. [Google Scholar] [CrossRef]
- Britton, S.; Coates, J.; Jackson, S.P. A new method for high-resolution imaging of Ku foci to decipher mechanisms of DNA double-strand break repair. J. Cell Biol. 2013, 202, 579–595. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurosawa, A. Autophosphorylation and Self-Activation of DNA-Dependent Protein Kinase. Genes 2021, 12, 1091. https://doi.org/10.3390/genes12071091
Kurosawa A. Autophosphorylation and Self-Activation of DNA-Dependent Protein Kinase. Genes. 2021; 12(7):1091. https://doi.org/10.3390/genes12071091
Chicago/Turabian StyleKurosawa, Aya. 2021. "Autophosphorylation and Self-Activation of DNA-Dependent Protein Kinase" Genes 12, no. 7: 1091. https://doi.org/10.3390/genes12071091
APA StyleKurosawa, A. (2021). Autophosphorylation and Self-Activation of DNA-Dependent Protein Kinase. Genes, 12(7), 1091. https://doi.org/10.3390/genes12071091