
Diversity of Modes of Reproduction and Sex Determination Systems in Invertebrates, and the Putative Contribution of Genetic Conflict


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Diverse Modes of Reproduction in Invertebrates, but «Sex» as an Ancient Feature
1.1. Reproductive Modes: Asexual and Sexual, and One Versus Two Parents
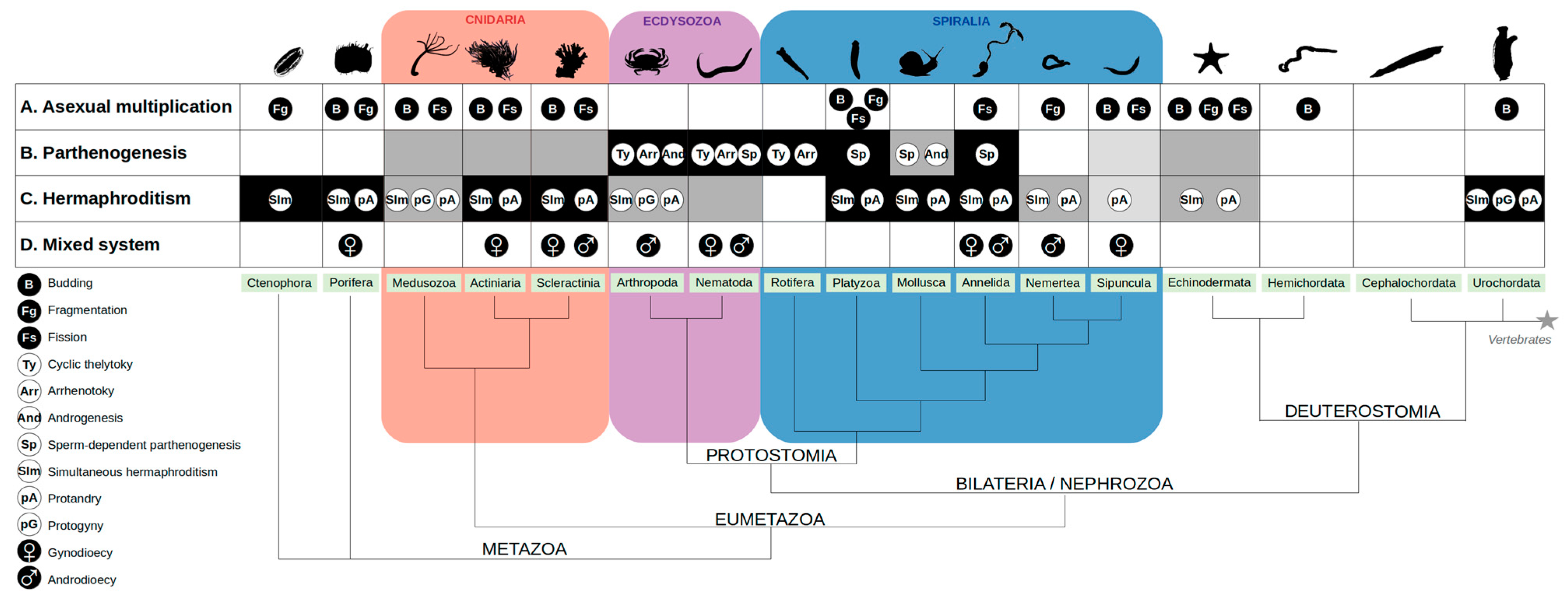
1.2. The Diversity of Reproductive Modes in Invertebrates
1.2.1. Asexual Lineages Are Usually Short Lived, but There Are Exceptions
1.2.2. Coexistence of Sexual and Asexual Mechanisms within the Same Species: When Invertebrates Benefit from Both Modes of Reproduction
1.2.3. Some Modes of Parthenogenesis Maintain Diversity without Sex
1.2.4. Fertilization without Genetic Exchange
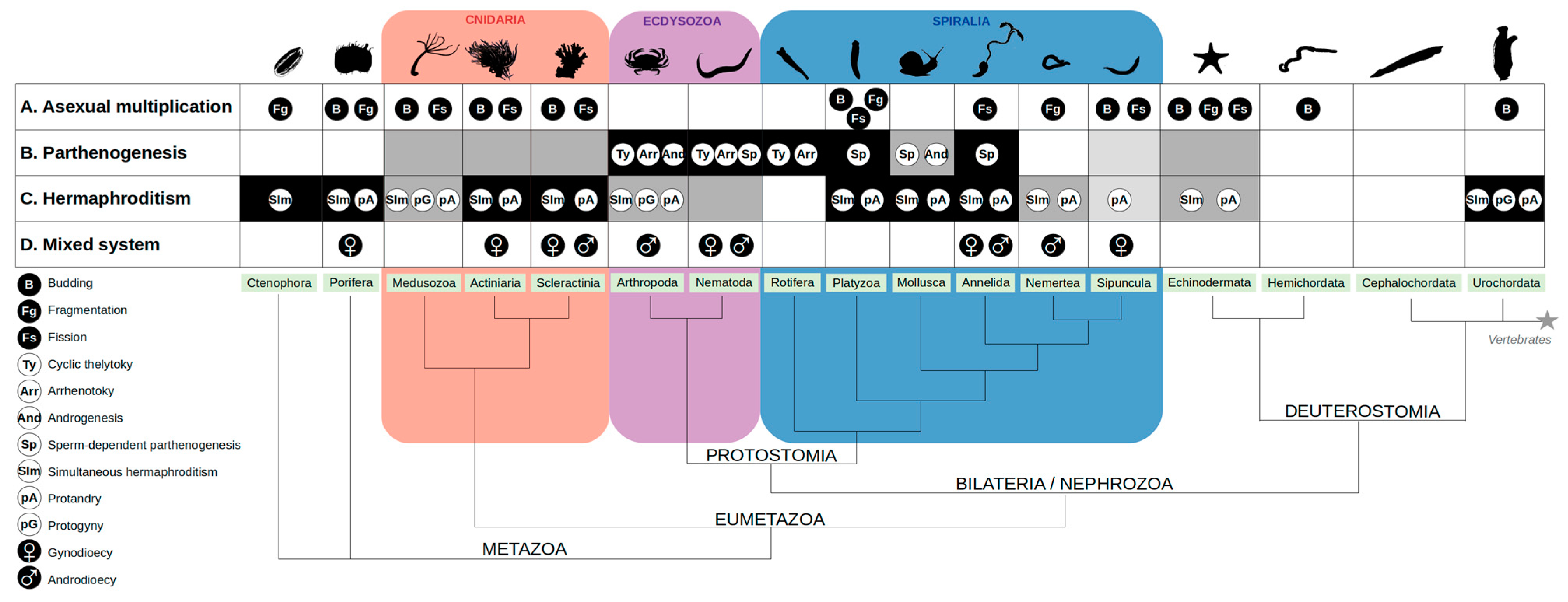
1.3. Sexual Reproduction: From Separate Sexes to Hermaphrodites, and Back
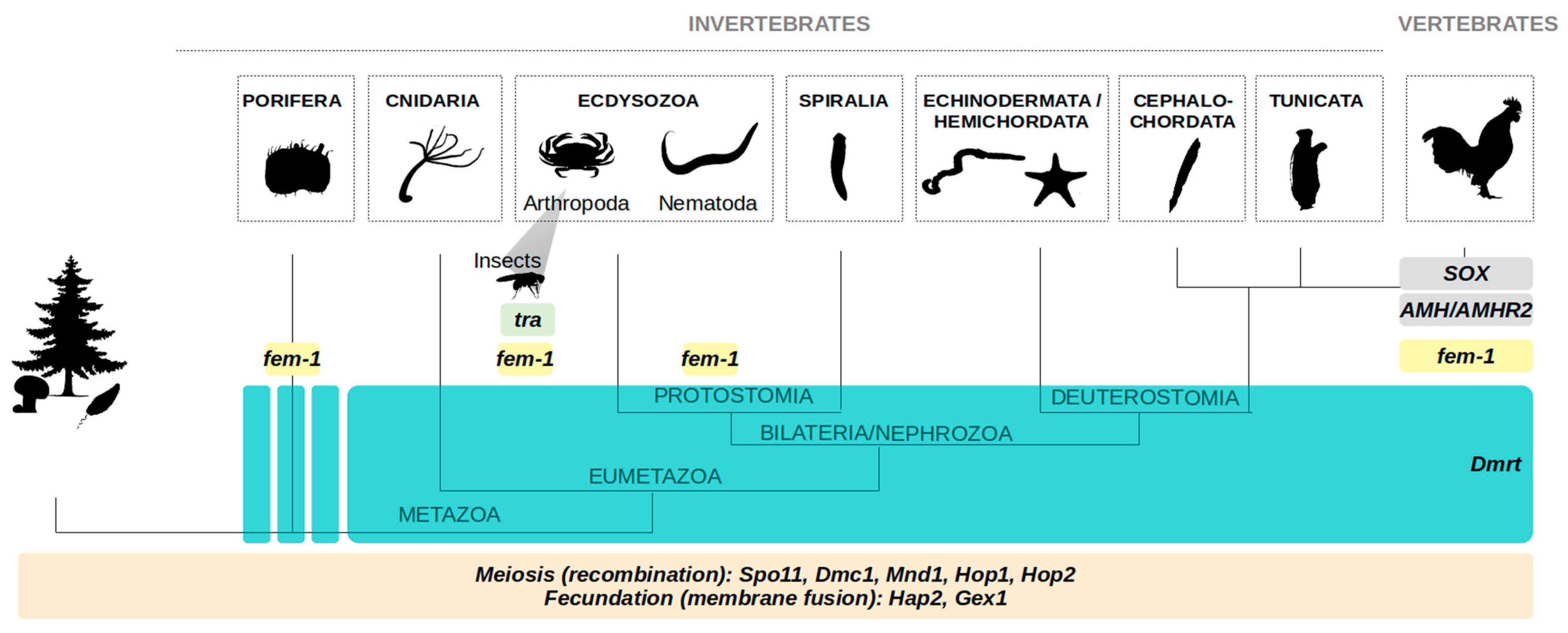
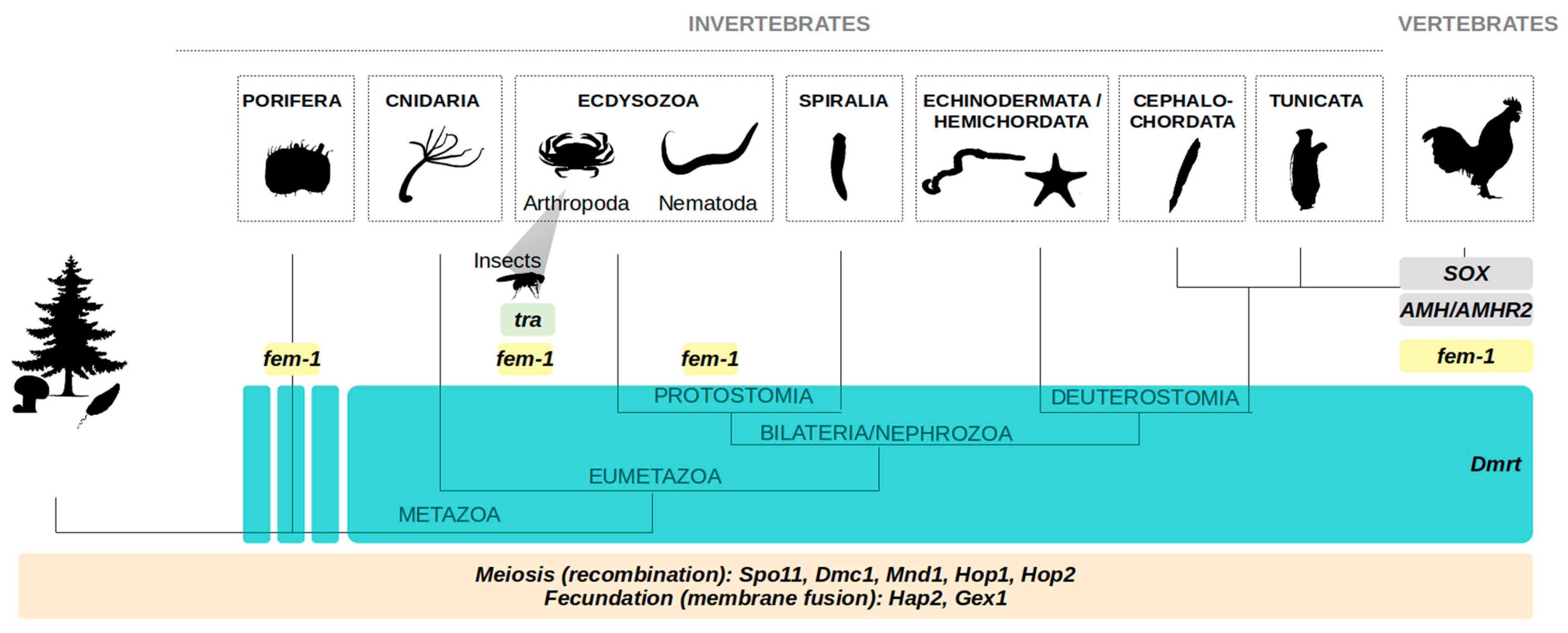
1.4. Sexual Reproduction as an Ancestral Feature: Some Insights from Shared Molecular Pathways
1.4.1. Meiosis and Fertilization Genes in Eukaryotes
1.4.2. Dmrt Genes in All Animals
1.4.3. Tra Gene in all Insects, and fem-1 in Ecdysozoa (and More)
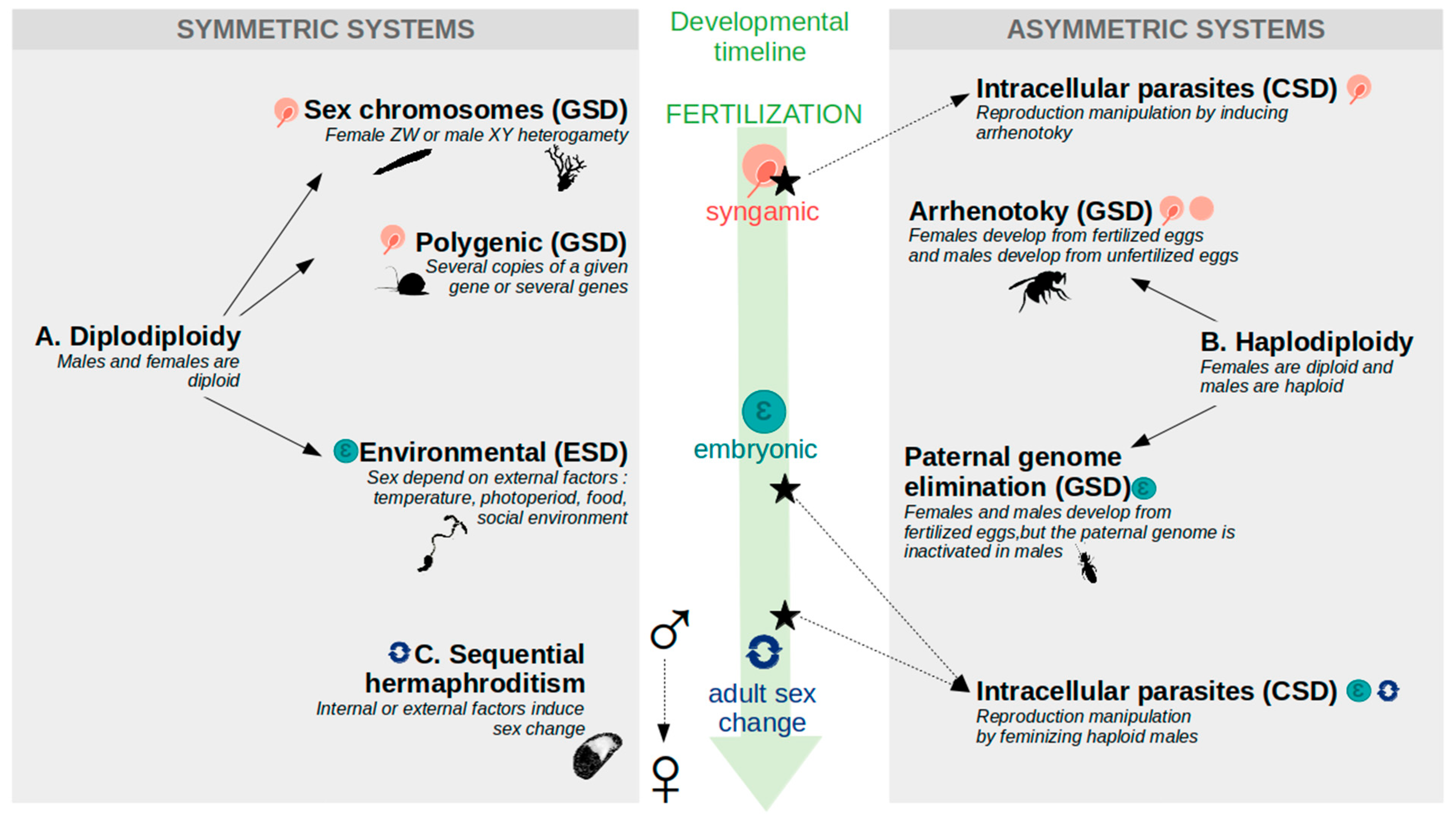
2. Invertebrate Species: A Legion of Primary Sex Determination Systems
2.1. Mechanisms and Timing of Sex Determination
2.2. Genetic Sex Determination System (GSD): Sex Determined at Fertilization
2.2.1. Sex Chromosomes
2.2.2. Polygenic Sex Determination (PSD)
2.2.3. Haplodiploidy
2.3. Environmental Sex Determination (ESD) during Embryonic Development
2.4. Sequential Hermaphrodites: Sex Changes Later in Life
2.5. Intracellular Parasites Can Highjack Sex Determination
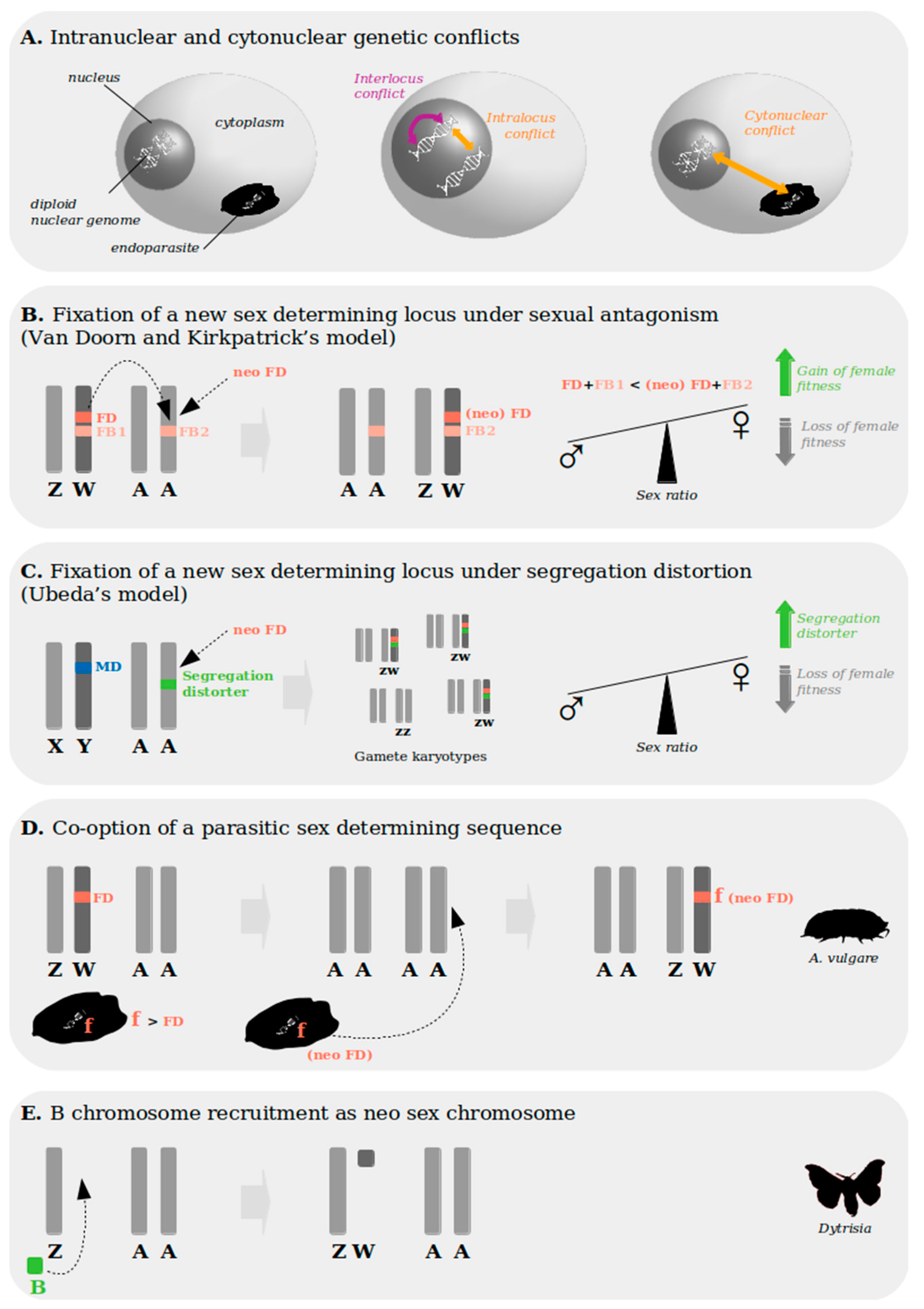
3. Genetic Conflict as a Potential Evolutionary Force Shaping Modes of Reproduction and Sex Determination Dynamics
3.1. Genetic Conflict and Transitions between Modes of Reproduction
3.1.1. Cytonuclear Conflict Can Promote the Switch to Asexuality
3.1.2. Genetic Conflicts Over Parental Transmission Driving Hermaphroditism
3.2. Genetic Conflict and Transitions between Sex Determination Systems
3.2.1. Hypotheses for the Establishment of Haplodiploidy: Parental and Host–Parasite Conflicts
3.2.2. Genetic Conflict and Sex Chromosome Turnover
3.3. Future Directions: Testing the Role of Genetic Conflict in the Age of Genomics
4. Conclusions
5. Glossary
5.1. Definitions in Alphabetical Order
5.2. Acronyms for Molecular Players of the Sex Determination Cascade
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Subramoniam, T. Mode of reproduction: Invertebrate animals. Encycl. Reprod. 2018, 6, 32–40. [Google Scholar]
- Esperk, T.; Tammaru, T.; Nylin, S.; Teder, T. Achieving high sexual size dimorphism in insects: Females add instars. Ecol. Entomol. 2007, 32, 243–256. [Google Scholar] [CrossRef]
- Mori, E.; Mazza, G.; Lovari, S. Sexual Dimorphism. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shakelford, T., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–7. [Google Scholar]
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Sex determination: Why so many ways of doing it? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef] [Green Version]
- Bell, G. The Masterpiece of Nature: The Evolution and Genetics of Sexuality; University of California: Berkeley, CA, USA, 1982. [Google Scholar]
- Smith, J.M. The Evolution of Sex; Cambridge University Press: Cambridge, UK, 1978. [Google Scholar]
- Schön, I.; Van Dijk, P.; Martens, K. Lost Sex: The Evolutionary Biology of Parthenogenesis; Springer: Dordrecht, The Netherlands, 2009; pp. 1–615. [Google Scholar]
- Judson, O.P.; Normark, B.B. Ancient asexual scandals. Trends Ecol. Evol. 1996, 11, 41–46. [Google Scholar] [CrossRef]
- Heethoff, M.; Norton, R.A.; Scheu, S.; Maraun, M. Parthenogenesis in oribatid mites (Acari, Oribatida): Evolution without sex. In Lost Sex: The Evolutionary Biology of Parthenogenesis; Springer: Dordrecht, The Netherlands, 2009; pp. 241–257. [Google Scholar]
- Schön, I.; Rossetti, G.; Martens, K. Darwinulid ostracods: Ancient asexual scandals or scandalous gossip. In Lost Sex: The Evolutionary Biology of Parthenogenesis; Springer: Dordrecht, The Netherlands, 2009; pp. 217–240. [Google Scholar]
- Schwander, T. Evolution: The end of an ancient asexual scandal. Curr. Biol. 2016, 26, R233–R235. [Google Scholar] [CrossRef] [Green Version]
- Debortoli, N.; Li, X.; Eyres, I.; Fontaneto, D.; Hespeels, B.; Tang, C.Q.; Flot, J.F.; Van Doninck, K. Genetic exchange among bdelloid rotifers is more likely due to horizontal gene transfer than to meiotic sex. Curr. Biol. 2016, 26, 723–732. [Google Scholar] [CrossRef] [Green Version]
- Signorovitch, A.; Hur, J.; Gladyshev, E.; Meselson, M. Allele sharing and evidence for sexuality in a mitochondrial clade of bdelloid rotifers. Genetics 2015, 200, 581–590. [Google Scholar] [CrossRef] [Green Version]
- Vakhrusheva, O.A.; Mnatsakanova, E.A.; Galimov, Y.R.; Neretina, T.V.; Gerasimov, E.S.; Naumenko, S.A.; Ozerova, S.G.; Zalevsky, A.O.; Yushenova, I.A.; Rodriguez, F.; et al. Genomic signatures of recombination in a natural population of the bdelloid rotifer Adineta Vaga. Nat. Commun. 2020, 11, 6421. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.G.; Nowell, R.W.; Barraclough, T.G. Cross-contamination explains “inter and intraspecific horizontal genetic transfers” between asexual bdelloid rotifers. Curr. Biol. 2018, 28, 2436–2444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, A.; Schaefer, I.; Glanz, J.; Schwander, T.; Maraun, M.; Scheu, S.; Bast, J. Effective purifying selection in ancient asexual oribatid mites. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Henry, L.; Schwander, T.; Crespi, B.J. Deleterious mutation accumulation in asexual timema stick insects. Mol. Biol. Evol. 2012, 29, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Siebert, S.; Juliano, C.E. Sex, polyps, and medusae: Determination and maintenance of sex in cnidarians. Mol. Reprod. Dev. 2017, 84, 105–119. [Google Scholar] [CrossRef]
- Pearcy, M.; Aron, S.; Doums, C.; Keller, L. Conditional use of sex and parthenogenesis for worker and queen production in ants. Science 2004, 306, 1780–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, L.; Jabbour-Zahab, R.; Mosna, M.; Haag, C.R.; Lenormand, T. Not so clonal asexuals: Unraveling the secret sex life of Artemia parthenogenetica. Evol. Lett. 2021, 5, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Koch, U.; Von Elert, E.; Straile, D. Food quality triggers the reproductive mode in the cyclical parthenogen Daphnia (Cladocera). Oecologia 2009, 159, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Engelstädter, J. Asexual but not clonal: Evolutionary processes in automictic populations. Genetics 2017, 206, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- Stenberg, P.; Saura, A. Meiosis and its deviations in polyploid animals. Cytogenet. Genome Res. 2013, 140, 185–203. [Google Scholar] [CrossRef]
- Nougué, O.; Rode, N.O.; Jabbour-zahab, R.; Ségard, A.; Chevin, L.M.; Haag, C.R.; Lenormand, T. Automixis in Artemia: Solving a century-old controversy. J. Evol. Biol. 2015, 28, 2337–2348. [Google Scholar] [CrossRef]
- Simon, J.C.; Stoeckel, S.; Tagu, D. Evolutionary and functional insights into reproductive strategies of aphids. Comptes Rendus Biol. 2010, 333, 488–496. [Google Scholar] [CrossRef]
- Simon, J.-C.; Delmotte, F.; Rispe, C.; Crease, T. Crease Phylogenetic relationships between parthenogens and their sexual relatives: The possible routes to parthenogenesis in animals. Biol. J. Linn. Soc. 2003, 79, 151–163. [Google Scholar] [CrossRef]
- Taylor, R.W.; Imai, H.T.; Hasegawa, E.; Beaton, C.D. A unique conjunction: Evidence for gynogenesis accompanying haplodiploid sex determination in the australian ant Myrmecia impaternata. Psyche 2018, 2018, 2832690. [Google Scholar] [CrossRef]
- Mantovani, B.; Scali, V. Preliminary report on a hybridogenetic stick-insect (Phasmatodea): The first case among invertebrates. Invertebr. Reprod. Dev. 1990, 18, 185–188. [Google Scholar] [CrossRef]
- Schwander, T.; Oldroyd, B.P. Androgenesis: Where males hijack eggs to clone themselves. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150534. [Google Scholar] [CrossRef]
- Pilger, J.F. Reproductive biology and development of Themiste lageniformis, a parthenogenic sipunculan. Bull. Mar. Sci. 1987, 41, 59–67. [Google Scholar]
- Jarne, P.; Auld, J.R. Animals mix it up too: The distribution of self-fertilization among hermaphroditic animals. Evolution 2006, 60, 1816–1824. [Google Scholar] [CrossRef] [PubMed]
- Normark, B.B.; Ross, L. Genetic conflict, kin and the origins of novel genetic systems. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lively, C.M.; Johnson, S.G. Brooding and the evolution of parthenogenesis: Strategy models and evidence from aquatic invertebrates. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1994, 256, 89–95. [Google Scholar]
- Combosch, D.J.; Vollmer, S.V. Mixed asexual and sexual reproduction in the Indo-Pacific reef coral Pocillopora damicornis. Ecol. Evol. 2013, 3, 3379–3387. [Google Scholar]
- Sasson, D.A.; Ryan, J.F. A reconstruction of sexual modes throughout animal evolution. BMC Evol. Biol. 2017, 17, 242. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, P.E. Gametogenesis and spawning in a hermaphroditic population of Golfingia minuta (sipuncula). J. Mar. Biol. Assoc. 1975, 55, 69–82. [Google Scholar] [CrossRef]
- Allsop, D.J.; West, S.A. Sex-ratio evolution in sex changing animals. Evolution (N.Y.) 2004, 58, 1019–1027. [Google Scholar]
- Weeks, S.C. The role of androdioecy and gynodioecy in mediating evolutionary transitions between dioecy and hermaphroditism in the animalia. Evolution 2012, 66, 3670–3686. [Google Scholar] [CrossRef] [PubMed]
- Singaravelu, G.; Singson, A. New insights into the mechanism of fertilization in nematodes. Int. Rev. Cell Mol. Biol. 2011, 289, 211–238. [Google Scholar] [PubMed] [Green Version]
- Gardner, A.; Ross, L. The evolution of hermaphroditism by an infectious male- derived cell lineage: An inclusive-fitness analysis. Am. Nat. 2011, 178, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Eppley, S.M.; Jesson, L.K. Moving to mate: The evolution of separate and combined sexes in multicellular organisms. J. Evol. Biol. 2008, 21, 727–736. [Google Scholar] [CrossRef]
- Charnov, E.L. Simultaneous hermaphroditism and sexual selection. Proc. Natl. Acad. Sci. USA 1979, 76, 2480–2484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basch, P.F. Why do schistosomes have separate sexes? Parasitol. Today 1990, 6, 160–163. [Google Scholar] [CrossRef]
- Kerr, A.M.; Baird, A.H.; Hughes, T.P. Correlated evolution of sex and reproductive mode in corals (Anthozoa: Scleractinia). Proc. R. Soc. B Biol. Sci. 2011, 278, 75–81. [Google Scholar] [CrossRef]
- Brantner, J.S.; Ott, D.W.; Duff, R.J.; Sanoamuang, L.O.; Simhachalam, G.P.; Babu, K.K.S.; Weeks, S.C. Androdioecy and hermaphroditism in five species of clam shrimps (Crustacea: Branchiopoda: Spinicaudata) from India and Thailand. Invertebr. Biol. 2013, 132, 27–37. [Google Scholar] [CrossRef]
- Ramesh, M.A.; Malik, S.B.; Logsdon, J.M. A phylogenomic inventory of meiotic genes: Evidence for sex in Giardia and an early eukaryotic origin of meiosis. Curr. Biol. 2005, 15, 185–191. [Google Scholar]
- Speijer, D.; Lukeš, J.; Eliáš, M. Sex is a ubiquitous, ancient, and inherent attribute of eukaryotic life. Proc. Natl. Acad. Sci. USA 2015, 112, 8827–8834. [Google Scholar] [CrossRef] [Green Version]
- Herpin, A.; Schartl, M. Plasticity of gene-regulatory networks controlling sex determination: Of masters, slaves, usual suspects, newcomers, and usurpators. EMBO Rep. 2015, 16, 1260–1274. [Google Scholar] [CrossRef] [Green Version]
- Kopp, A. Dmrt genes in the development and evolution of sexual dimorphism. Trends Genet. 2012, 28, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Wexler, J.R.; Plachetzki, D.C.; Kopp, A. Pan-metazoan phylogeny of the DMRT gene family: A framework for functional studies. Dev. Genes Evol. 2014, 224, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Picard, M.A.L.; Cosseau, C.; Mouahid, G.; Duval, D.; Grunau, C.; Toulza, È.; Allienne, J.F.; Boissier, J. The roles of Dmrt (Double sex/Male-abnormal-3 Related Transcription factor) genes in sex determination and differentiation mechanisms: Ubiquity and diversity across the animal kingdom. Comptes Rendus Biol. 2015, 338, 451–462. [Google Scholar] [CrossRef]
- Riesgo, A.; Farrar, N.; Windsor, P.J.; Giribet, G.; Leys, S.P. The analysis of eight transcriptomes from all poriferan classes reveals surprising genetic complexity in sponges. Mol. Biol. Evol. 2014, 31, 1102–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.; Kobayashi, K.; Watanabe, H.; Iguchi, T. Environmental sex determination in the branchiopod crustacean Daphnia magna: Deep conservation of a Doublesex gene in the sex-determining pathway. PLoS Genet. 2011, 7, e1001345. [Google Scholar] [CrossRef] [Green Version]
- Chong, T.; Collins, J.J.; Brubacher, J.L.; Zarkower, D.; Newmark, P.A. A sex-specific transcription factor controls male identity in a simultaneous hermaphrodite. Nat. Commun. 2013, 4, 1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhulst, E.C.; van de Zande, L.; Beukeboom, L.W. Insect sex determination: It all evolves around transformer. Curr. Opin. Genet. Dev. 2010, 20, 376–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasselmann, M.; Gempe, T.; Schiøtt, M.; Nunes-Silva, C.G.; Otte, M.; Beye, M. Evidence for the evolutionary nascence of a novel sex determination pathway in honeybees. Nature 2008, 454, 519–522. [Google Scholar] [CrossRef]
- Song, C.; Cui, Z.; Hui, M.; Liu, Y.; Li, Y. Molecular characterization and expression profile of three Fem-1 genes in Eriocheir sinensis provide a new insight into crab sex-determining mechanism. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2015, 189, 6–14. [Google Scholar] [CrossRef]
- Shi, H.; Chen, B.; He, Z. Cloning and expression analysis of the prolixicin gene from Locusta migratoria manilensis (meyen). Chin. J. Appl. Environ. Biol. 2013, 19, 236–240. [Google Scholar] [CrossRef]
- Ma, K.Y.; Liu, Z.Q.; Lin, J.Y.; Li, J.L.; Qiu, G.F. Molecular characterization of a novel ovary-specific gene fem-1 homolog from the oriental river prawn, Macrobrachium nipponense. Gene 2016, 575, 244–252. [Google Scholar] [CrossRef]
- Lipani, C.; Vitturi, R.; Sconzo, G.; Barbata, G. Karyotype analysis of the sea urchin Paracentrotus lividus (echinodermata): Evidence for a heteromorphic chromosome sex mechanism. Mar. Biol. 1996, 127, 67–72. [Google Scholar] [CrossRef]
- Pratlong, M.; Haguenauer, A.; Chenesseau, S.; Brener, K.; Mitta, G.; Toulza, E.; Bonabaud, M.; Rialle, S.; Aurelle, D.; Pontarotti, P. Evidence for a genetic sex determination in Cnidaria, the Mediterranean red coral (Corallium rubrum). R. Soc. Open Sci. 2017, 4, 160880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bliznina, A.; Masunaga, A.; Mansfield, M.J.; Tan, Y.; Liu, A.W.; West, C.; Rustagi, T.; Chien, H.C.; Kumar, S.; Pichon, J.; et al. Telomere-to-telomere assembly of the genome of an individual Oikopleura dioica from Okinawa using Nanopore-based sequencing. BMC Genom. 2021, 22, 222. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Wu, X.; Su, L.; Shang, C.; Li, X.; Wang, Y.; Li, G. A ZZ/ZW sex chromosome system in cephalochordate Amphioxus. Genetics 2020, 214, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Fraïsse, C.; Picard, M.A.L.; Vicoso, B. The deep conservation of the Lepidoptera Z chromosome suggests a non-canonical origin of the W. Nat. Commun. 2017, 8, 1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicoso, B.; Bachtrog, D. Numerous Transitions of Sex Chromosomes in Diptera. PLoS Biol. 2015, 13, e1002078. [Google Scholar] [CrossRef] [Green Version]
- Meccariello, A.; Salvemini, M.; Primo, P.; Hall, B.; Koskinioti, P.; Dalíková, M.; Gravina, A.; Gucciardino, M.A.; Forlenza, F.; Gregoriou, M.E.; et al. Maleness-on-the-Y (MoY) orchestrates male sex determination in major agricultural fruit fly pests. Science 2019, 365, 1457–1460. [Google Scholar] [CrossRef]
- Sharma, A.; Heinze, S.D.; Wu, Y.; Kohlbrenner, T.; Morilla, I.; Brunner, C.; Wimmer, E.A.; Van De Zande, L.; Robinson, M.D.; Beukeboom, L.W.; et al. Male sex in houseflies is determined by Mdmd, a paralog of the generic splice factor gene CWC22. Science 2017, 356, 642–645. [Google Scholar] [CrossRef]
- Krzywinska, E.; Dennison, N.J.; Lycett, G.J.; Krzywinski, J. A maleness gene in the malaria mosquito Anopheles gambiae. Science 2016, 353, 67–69. [Google Scholar] [CrossRef] [Green Version]
- Hall, A.B.; Basu, S.; Jiang, X.; Qi, Y.; Timoshevskiy, V.A.; Biedler, J.K.; Sharakhova, M.V.; Elahi, R.; Anderson, M.A.E.; Chen, X.G.; et al. A male-determining factor in the mosquito Aedes aegypti. Science 2015, 348, 1268–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicoso, B. Molecular and evolutionary dynamics of animal sex-chromosome turnover. Nat. Ecol. Evol. 2019, 3, 1632–1641. [Google Scholar] [CrossRef] [PubMed]
- Hamm, R.L.; Meisel, R.P.; Scott, J.G. The evolving puzzle of autosomal versus Y-linked male determination in Musca domestica. G3 Genes Genomes Genet. 2015, 5, 371–384. [Google Scholar]
- Moore, E.C.; Roberts, R.B. Polygenic sex determination. Curr. Biol. 2013, 23, R510–R512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallman, K.D. The sex-determining mechanism of the platyfish, Xiphophorus maculatus. In Genetics and Mutagenesis of Fish; Springer: Berlin/Heidelberg, Germany, 1973; pp. 19–28. [Google Scholar]
- Veyrunes, F.; Chevret, P.; Catalan, J.; Castiglia, R.; Watson, J.; Dobigny, G.; Robinson, T.J.; Britton-Davidian, J. A novel sex determination system in a close relative of the house mouse. Proc. R. Soc. B Biol. Sci. 2010, 277, 1049–1056. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.L.; Marí, A.; Braasch, I.; Amores, A.; Hohenlohe, P.; Batzel, P.; Postlethwait, J.H. Multiple sex-associated regions and a putative sex chromosome in zebrafish revealed by RAD mapping and population genomics. PLoS ONE 2012, 7, e40701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusa, Y. Nuclear sex-determining genes cause large sex-ratio variation in the apple snail Pomacea canaliculata. Genetics 2007, 175, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Alexander, H.J.; Richardson, J.M.L.; Edmands, S.; Anholt, B.R. Sex without sex chromosomes: Genetic architecture of multiple loci independently segregating to determine sex ratios in the copepod Tigriopus californicus. J. Evol. Biol. 2015, 28, 2196–2207. [Google Scholar] [CrossRef]
- Blackmon, H.; Ross, L.; Bachtrog, D. Sex determination, sex chromosomes, and karyotype evolution in insects. J. Hered. 2017, 108, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Hodson, C.N.; Hamilton, P.T.; Dilworth, D.; Nelson, C.J.; Curtis, C.I.; Perlman, S.J. Paternal genome elimination in Liposcelis booklice (Insecta: Psocodea). Genetics 2017, 206, 1091–1100. [Google Scholar] [CrossRef] [Green Version]
- Van Wilgenburg, E.; Driessen, G.; Beukeboom, L.W. Single locus complementary sex determination in Hymenoptera: An “unintelligent” design? Front. Zool. 2006, 3, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beukeboom, L.W.; van de Zande, L. Genetics of sex determination in the haplodiploid wasp Nasonia vitripennis (Hymenoptera: Chalcidoidea). J. Genet. 2010, 89, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Walker, G. Sex determination in the larvae of the parasitic barnacle Heterosaccus lunatus: An experimental approach. J. Exp. Mar. Biol. Ecol. 2005, 318, 31–38. [Google Scholar] [CrossRef]
- McCabe, J.; Dunn, A.M. Adaptive significance of environmental sex determination in an amphipod. J. Evol. Biol. 1997, 10, 515–527. [Google Scholar] [CrossRef]
- Berec, L.; Schembri, P.J.; Boukal, D.S. Sex determination in Bonellia viridis (Echiura: Bonelliidae): Population dynamics and evolution. Oikos 2005, 108, 473–484. [Google Scholar] [CrossRef] [Green Version]
- Chiba, S. A review of ecological and evolutionary studies on hermaphroditic decapod crustaceans. Plankt. Benthos Res. 2007, 2, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef]
- Cordaux, R.; Bouchon, D.; Grève, P. The impact of endosymbionts on the evolution of host sex-determination mechanisms. Trends Genet. 2011, 27, 332–341. [Google Scholar] [CrossRef]
- Van Doorn, G.S.; Kirkpatrick, M. Turnover of sex chromosomes induced by sexual conflict. Nature 2007, 449, 909–912. [Google Scholar] [CrossRef]
- Úbeda, F.; Patten, M.M.; Wild, G. On the origin of sex chromosomes from meiotic drive. Proc. R. Soc. B Biol. Sci. 2014, 282, 20141932. [Google Scholar] [CrossRef]
- Leclercq, S.; Thézé, J.; Chebbi, M.A.; Giraud, I.; Moumen, B.; Ernenwein, L.; Grève, P.; Gilbert, C.; Cordaux, R. Birth of a W sex chromosome by horizontal transfer of Wolbachia bacterial symbiont genome. Proc. Natl. Acad. Sci. USA 2016, 113, 15036–15041. [Google Scholar] [CrossRef] [Green Version]
- Russell, J.E.; Stouthamer, R. The genetics and evolution of obligate reproductive parasitism in Trichogramma pretiosum infected with parthenogenesis-inducing Wolbachia. Heredity 2011, 106, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Rošciszewska1, E.L.; Soldán2, T. Morphology of accessory ovaries in adult males of Perla marginata (Plecoptera: Perlidae). Eur. J. Entomol. 1999, 96, 45–51. [Google Scholar]
- Normark, B.B. Unusual gametic and genetic systems. Sperm Biol. 2009, 507–538. [Google Scholar]
- Ross, L.; Pen, I.; Shuker, D.M. Genomic conflict in scale insects: The causes and consequences of bizarre genetic systems. Biol. Rev. 2010, 85, 807–828. [Google Scholar] [CrossRef] [PubMed]
- Normark, B.B. Perspective: Maternal kin groups and the origins of asymmetric genetic systems—Genomic imprinting, haplodiploidy, and parthenogenesis. Evolution 2006, 60, 631–642. [Google Scholar] [PubMed]
- de la Filia, A.G.; Fenn-Moltu, G.; Ross, L. No evidence for an intragenomic arms race under paternal genome elimination in Planococcus mealybugs. J. Evol. Biol. 2019, 32, 491–504. [Google Scholar] [CrossRef] [Green Version]
- Blackmon, H.; Hardy, N.B.; Ross, L. The evolutionary dynamics of haplodiploidy: Genome architecture and haploid viability. Evolution 2015, 69, 2971–2978. [Google Scholar] [CrossRef]
- Engelstadter, J.; Hurst, G.D.D. Can maternally transmitted endosymbionts facilitate the evolution of haplodiploidy? J. Evol. Biol. 2006, 19, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Normark, B.B. Haplodiploidy as an outcome of coevolution between male-killing cytoplasmic elements and their hosts. Evolution 2004, 58, 790–798. [Google Scholar] [CrossRef]
- Ross, L.; Shuker, D.M.; Normark, B.B.; Pen, I. The role of endosymbionts in the evolution of haploid-male genetic systems in scale insects (Coccoidea). Ecol. Evol. 2012, 2, 1071–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlin, S.; Lessard, S. Theoretical Studies on Sex Ratio Evolution; Princeton University Press: Princeton, NJ, USA, 1987. [Google Scholar]
- Werren, J.H.; Beukeboom, L.W. Sex determination, sex ratios, and genetic conflict. Annu. Rev. Ecol. Syst. 1998, 29, 233–261. [Google Scholar] [CrossRef] [Green Version]
- Mank, J.E.; Hosken, D.J.; Wedell, N. Conflict on the sex chromosomes: Cause, effect, and complexity. Cold Spring Harb. Perspect. Biol. 2014, 6, a017715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Doorn, G.S.; Kirkpatrick, M. Transitions between male and female heterogamety caused by sex-antagonistic selection. Genetics 2010, 186, 629–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozielska, M.; Weissing, F.J.; Beukeboom, L.W.; Pen, I. Segregation distortion and the evolution of sex-determining mechanisms. Heredity 2010, 104, 100–112. [Google Scholar] [CrossRef] [Green Version]
- Meiklejohn, C.D.; Tao, Y. Genetic conflict and sex chromosome evolution. Trends Ecol. Evol. 2010, 25, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chebbi, M.A.; Becking, T.; Moumen, B.; Giraud, I.; Gilbert, C.; Peccoud, J.; Cordaux, R. The genome of Armadillidium vulgare (Crustacea, Isopoda) provides insights into sex chromosome evolution in the context of cytoplasmic sex determination. Mol. Biol. Evol. 2019, 36, 727–741. [Google Scholar] [CrossRef] [Green Version]
- Camacho, J.P.M.; Sharbel, T.F.; Beukeboom, L.W. B-chromosome evolution. Philos. Trans. R. Soc. B Biol. Sci. 2000, 355, 163–178. [Google Scholar] [CrossRef]
- Carvalho, A.B. Origin and evolution of the Drosophila Y chromosome. Curr. Opin. Genet. Dev. 2002, 12, 664–668. [Google Scholar] [CrossRef]
- Nokkala, S.; Grozeva, S.; Kuznetsova, V.; Maryanska-Nadachowska, A. The origin of the achiasmatic XY sex chromosome system in Cacopsylla peregrina (Psylloidea, Homoptera). Genetica 2003, 119, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Hackstein, J.H.P.; Hochstenbach, R.; Hauschteck-Jungen, E.; Beukeboom, L.W. Is the Y chromosome of Drosophila an evolved supernumerary chromosome? BioEssays 1996, 18, 317–323. [Google Scholar] [CrossRef] [Green Version]
- Kuznetsova, V.G.; Nokkala, S.; Maryanska-Nadachowska, A. Karyotypes, sex chromosome systems, and male meiosis in Finnish psyllids (Homoptera: Psylloidea). Folia Biol. 1997, 45, 143–152. [Google Scholar]
- Lukhtanov, V.A. Sex chromatin and sex chromosome systems in nonditrysian Lepidoptera (Insecta). J. Zool. Syst. Evol. Res. 2000, 38, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Kirkpatrick, M. Sex-specific selection and sex-biased gene expression in humans and flies. PLoS Genet. 2016, 12, e1006170. [Google Scholar] [CrossRef]
- Lucotte, E.A.; Laurent, R.; Heyer, E.; Ségurel, L.; Toupance, B. Detection of allelic frequency differences between the sexes in humans: A signature of sexually antagonistic selection. Genome Biol. Evol. 2016, 8, 1489–1500. [Google Scholar] [CrossRef] [Green Version]
- Ruzicka, F.; Dutoit, L.; Czuppon, P.; Jordan, C.Y.; Li, X.; Olito, C.; Runemark, A.; Svensson, E.I.; Yazdi, H.P.; Connallon, T. The search for sexually antagonistic genes: Practical insights from studies of local adaptation and statistical genomics. Evol. Lett. 2020, 4, 398–415. [Google Scholar] [CrossRef]
- Wijchers, P.J.; Festenstein, R.J. Epigenetic regulation of autosomal gene expression by sex chromosomes. Trends Genet. 2011, 27, 132–140. [Google Scholar] [CrossRef]
- Son, J.H.; Kohlbrenner, T.; Heinze, S.; Beukeboom, L.W.; Bopp, D.; Meisel, R.P. Minimal effects of proto-Y chromosomes on house fly gene expression in spite of evidence that selection maintains stable polygenic sex determination. Genetics 2019, 213, 313–327. [Google Scholar] [CrossRef]
- Meisel, R.P.; Gonzales, C.A.; Luu, H. The house fly y Chromosome is young and minimally differentiated from its ancient X chromosome partner. Genome Res. 2017, 27, 1417–1426. [Google Scholar] [CrossRef] [Green Version]
- Son, J.H.; Meisel, R.P. Gene-level, but not chromosome-wide, divergence between a very young house fly proto-Y chromosome and its homologous proto-X chromosome. Mol. Biol. Evol. 2021, 38, 606–618. [Google Scholar] [CrossRef] [PubMed]
- de la Filia, A.G.; Mongue, A.J.; Dorrens, J.; Lemon, H.; Laetsch, D.R.; Ross, L. Males that silence their father’s genes: Genomic imprinting of a complete haploid genome. Mol. Biol. Evol. 2021, 38, 2566–2581. [Google Scholar] [CrossRef] [PubMed]
- Jaron, K.S.; Bast, J.; Nowell, R.W.; Ranallo-Benavidez, T.R.; Robinson-Rechavi, M.; Schwander, T. Genomic features of parthenogenetic animals. J. Hered. 2021, 112, 19–33. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Picard, M.A.L.; Vicoso, B.; Bertrand, S.; Escriva, H. Diversity of Modes of Reproduction and Sex Determination Systems in Invertebrates, and the Putative Contribution of Genetic Conflict. Genes 2021, 12, 1136. https://doi.org/10.3390/genes12081136
Picard MAL, Vicoso B, Bertrand S, Escriva H. Diversity of Modes of Reproduction and Sex Determination Systems in Invertebrates, and the Putative Contribution of Genetic Conflict. Genes. 2021; 12(8):1136. https://doi.org/10.3390/genes12081136
Chicago/Turabian StylePicard, Marion Anne Lise, Beatriz Vicoso, Stéphanie Bertrand, and Hector Escriva. 2021. "Diversity of Modes of Reproduction and Sex Determination Systems in Invertebrates, and the Putative Contribution of Genetic Conflict" Genes 12, no. 8: 1136. https://doi.org/10.3390/genes12081136
APA StylePicard, M. A. L., Vicoso, B., Bertrand, S., & Escriva, H. (2021). Diversity of Modes of Reproduction and Sex Determination Systems in Invertebrates, and the Putative Contribution of Genetic Conflict. Genes, 12(8), 1136. https://doi.org/10.3390/genes12081136