
A Missense Variant in SLC39A4 in a Litter of Turkish Van Cats with Acrodermatitis Enteropathica

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Selection
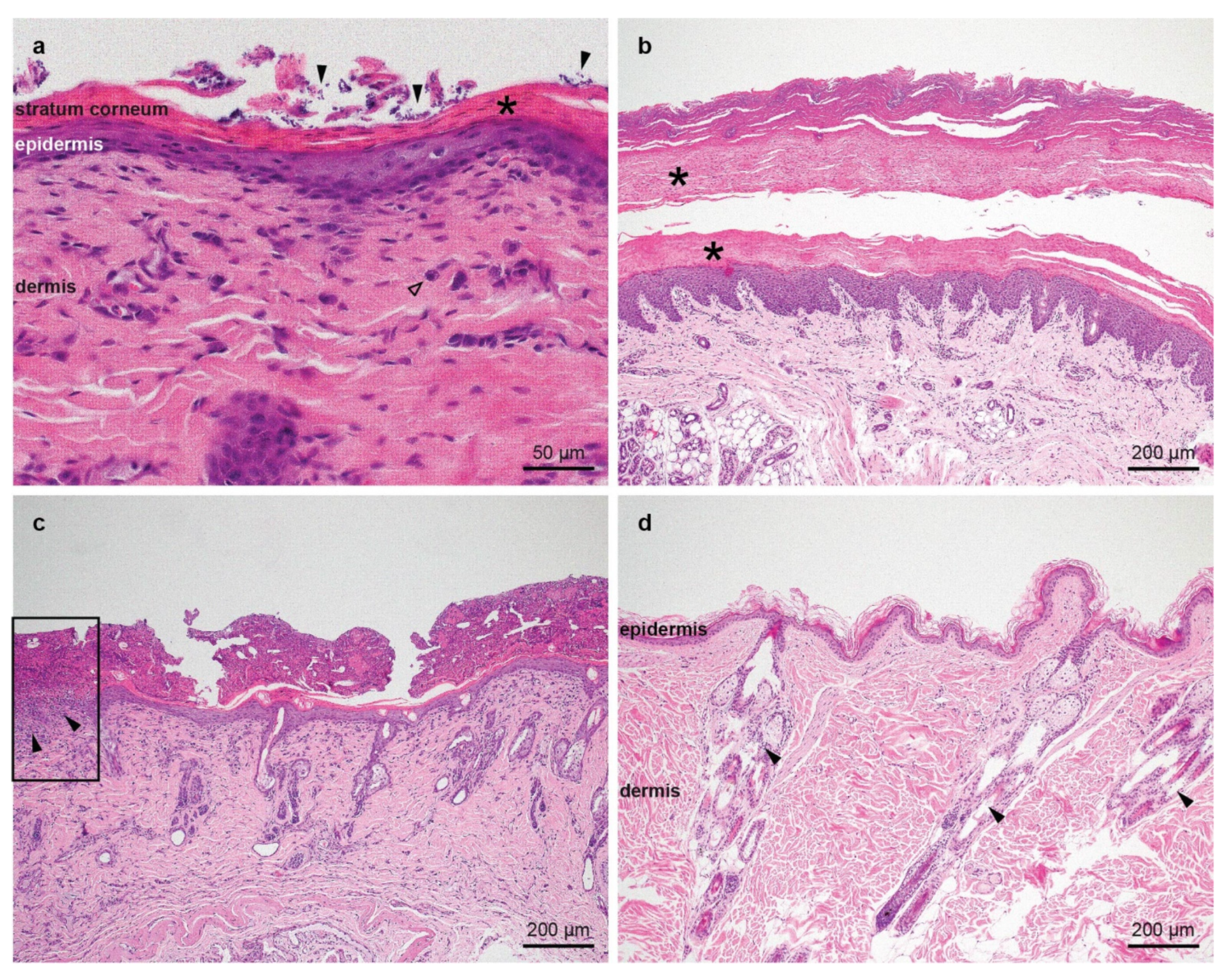
2.2. Histopathological Examinations
2.3. Whole Genome Sequencing
2.4. Variant Calling
2.5. Gene Analysis
2.6. Sanger Sequencing
3. Results
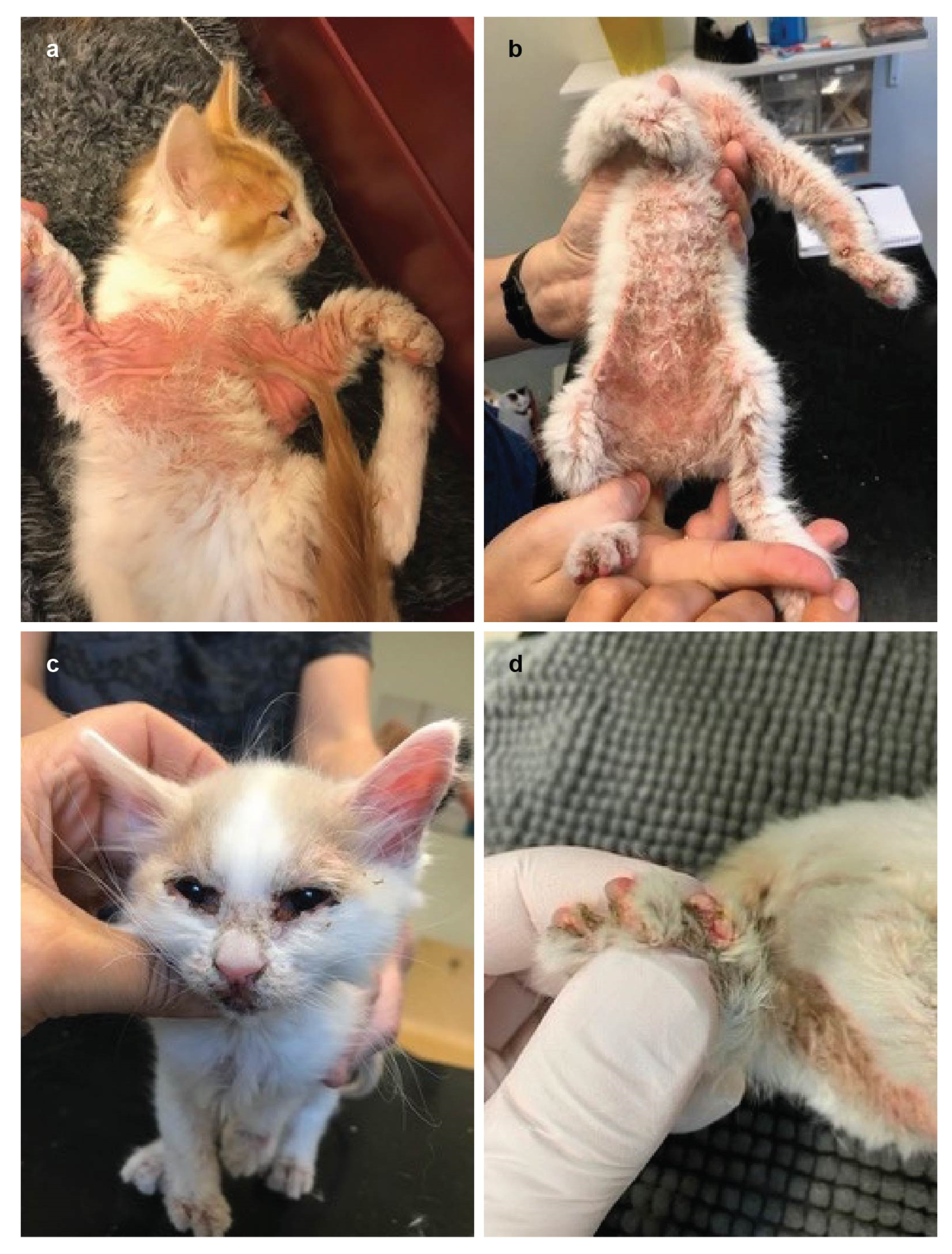
3.1. Family Anamnesis, Clinical Examinations and Histopathology
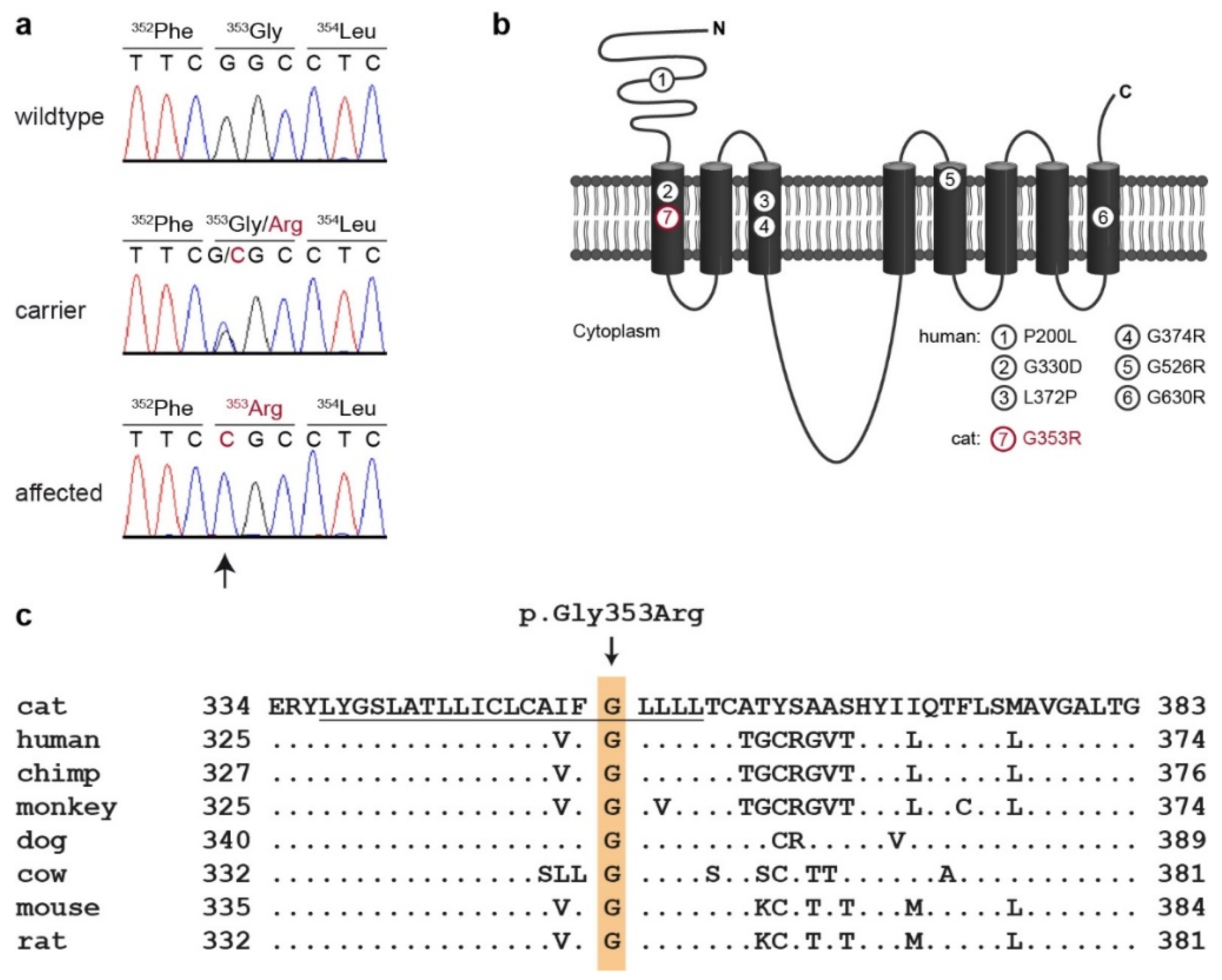
3.2. Genetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jezyk, P.F.; Haskins, M.E.; MacKay-Smith, W.E.; Patterson, D.F. Lethal Acrodermatitis in Bull Terriers. Am. Vet. Med. Assoc. 1986, 15, 833–839. [Google Scholar]
- McEwan, N.A. Lethal Acrodermatitis of Bull Terriers. Vet. Rec. 1990, 127, 95. [Google Scholar] [PubMed]
- Mcewan, N.A.; Mcneil, P.E.; Thompson, H.; Mccandlish, I.A.P. Diagnostic Features, Confirmation and Disease Progression in 28 Cases of Lethal Acrodermatitis of Bull Terriers. J. Small Anim. Pract. 2000, 41, 501–507. [Google Scholar] [CrossRef]
- Uchida, Y.; Moon-Fanelli, A.A.; Dodman, N.H.; Clegg, M.S.; Keen, C.L. Serum Concentrations of Zinc and Copper in Bull Terriers with Lethal Acrodermatitis and Tail-Chasing Behavior. Am. J. Vet. Res. 1997, 58, 808–810. [Google Scholar]
- Bauer, A.; Jagannathan, V.; Högler, S.; Richter, B.; McEwan, N.A.; Thomas, A.; Cadieu, E.; André, C.; Hytönen, M.K.; Lohi, H.; et al. MKLN1 Splicing Defect in Dogs with Lethal Acrodermatitis. PLoS Genet. 2018, 14, e1007264. [Google Scholar] [CrossRef]
- Adams, J.C.; Seed, B.; Lawler, J. Muskelin, a Novel Intracellular Mediator of Cell Adhesive and Cytoskeletal Responses to Thrombospondin-1. EMBO J. 1998, 17, 4964–4974. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, H.; Katoh, H.; Fujita, H.; Mori, K.; Negishi, M. Receptor Isoform-Specific Interaction of Prostaglandin EP3 Receptor with Muskelin. Biochem. Biophys. Res. Commun. 2000, 276, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Umeda, M.; Nishitani, H.; Nishimoto, T. A Novel Nuclear Protein, Twa1, and Muskelin Comprise a Complex with RanBPM. Gene 2003, 303, 47–54. [Google Scholar] [CrossRef]
- Prag, S.; Collett, G.D.M.; Adams, J.C. Molecular Analysis of Muskelin Identifies a Conserved Discoidin-like Domain That Contributes to Protein Self-Association. Biochem. J. 2004, 381, 547–559. [Google Scholar] [CrossRef]
- Kobayashi, N.; Yang, J.; Ueda, A.; Suzuki, T.; Tomaru, K.; Takeno, M.; Okuda, K.; Ishigatsubo, Y. RanBPM, Muskelin, P48EMLP, P44CTLH, and the Armadillo-Repeat Proteins ARMC8α and ARMC8β Are Components of the CTLH Complex. Gene 2007, 396, 236–247. [Google Scholar] [CrossRef]
- Valiyaveettil, M.; Bentley, A.A.; Gursahaney, P.; Hussien, R.; Chakravarti, R.; Kureishy, N.; Prag, S.; Adams, J.C. Novel Role of the Muskelin-RanBP9 Complex as a Nucleocytoplasmic Mediator of Cell Morphology Regulation. J. Cell Biol. 2008, 182, 727–739. [Google Scholar] [CrossRef]
- DeBenedittis, P.; Harmelink, C.; Chen, Y.; Wang, Q.; Jiao, K. Characterization of the Novel Interaction between Muskelin and TBX20, a Critical Cardiogenic Transcription Factor. Biochem. Biophys. Res. Commun. 2011, 409, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Heisler, F.F.; Loebrich, S.; Pechmann, Y.; Maier, N.; Zivkovic, A.R.; Tokito, M.; Hausrat, T.J.; Schweizer, M.; Bähring, R.; Holzbaur, E.L.F.; et al. Muskelin Regulates Actin Filament- and Microtubule-Based GABAA Receptor Transport in Neurons. Neuron 2011, 70, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Francis, O.; Han, F.; Adams, J.C. Molecular Phylogeny of a RING E3 Ubiquitin Ligase, Conserved in Eukaryotic Cells and Dominated by Homologous Components, the Muskelin/RanBPM/CTLH Complex. PLoS ONE 2013, 8, e75217. [Google Scholar] [CrossRef] [PubMed]
- Gueron, G.; Giudice, J.; Valacco, P.; Paez, A.; Elguero, B.; Toscani, M.; Jaworski, F.; Leskow, F.C.; Cotignola, J.; Marti, M.; et al. Heme-Oxygenase-1 Implications in Cell Morphology and the Adhesive Behavior of Prostate Cancer Cells. Oncotarget 2014, 5, 4087–4102. [Google Scholar] [CrossRef]
- Tripathi, B.K.; Lowy, D.R.; Zelenka, P.S. The Cdk5 Activator P39 Specifically Links Muskelin to Myosin II and Regulates Stress Fiber Formation and Actin Organization in Lens. Exp. Cell Res. 2015, 330, 186–198. [Google Scholar] [CrossRef][Green Version]
- Danbolt, N.; Closs, K. Acrodermatitis Enteropathica. Acta Derm. Venereol. 1942, 23, 127–169. [Google Scholar] [CrossRef]
- Machen, M.; Montgomery, T.; Holland, R.; Braselton, E.; Dunstan, R.; Brewer, G.; Yuzbasiyan-Gurkan, V. Bovine Hereditary Zinc Deficiency: Lethal Trait A 46. J. Vet. Diagn. Investig. 1996, 8, 219–227. [Google Scholar] [CrossRef]
- Moynahan, E.J.; Barnes, P.M. Zinc Deficiency and a Synthetic Diet for Lactose Intolerance. Lancet 1973, 301, 676–677. [Google Scholar] [CrossRef]
- Moynahan, E.J. Acrodermatitis Enteropathica: A Lethal Inherited Human Zinc-Deficiency Disorder. Lancet 1974, 304, 399–400. [Google Scholar] [CrossRef]
- Michaëlsson, G. Zinc Therapy in Acrodermatitis Enteropathica. Acta Derm. Venereol. 1974, 54, 377–381. [Google Scholar]
- Thyresson, N. Acrodermatitis Enteropathica. Report of a Case Healed with Zinc Therapy. Acta Derm. Venereol. 1974, 54, 383–385. [Google Scholar]
- Mccall, K.A.; Huang, C.-C.; Fierke, C.A. Function and Mechanism of Zinc Metalloenzymes. J. Nutr. 2000, 130, 1437S–1445S. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Counting the Zinc-Proteins Encoded in the Human Genome. J. Proteome Res. 2006, 5, 196–201. [Google Scholar] [CrossRef]
- Küry, S.; Dréno, B.; Bézieau, S.; Giraudet, S.; Kharfi, M.; Kamoun, R.; Moisan, J.P. Identification of SLC39A4, a Gene Involved in Acrodermatitis Enteropathica. Nat. Genet. 2002, 31, 239–240. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhou, B.; Kuo, Y.M.; Zemansky, J.; Gitschier, J. A Novel Member of a Zinc Transporter Family Is Defective in Acrodermatitis Enteropathica. Am. J. Hum. Genet. 2002, 71, 66–73. [Google Scholar] [CrossRef]
- Nakano, A.; Nakano, H.; Nomura, K.; Toyomaki, Y.; Hanada, K. Novel SLC39A4 Mutations in Acrodermatitis Enteropathica. J. Investig. Dermatol. 2003, 120, 963–966. [Google Scholar] [CrossRef]
- Yuzbasiyan-Gurkan, V.; Bartlett, E. Identification of a Unique Splice Site Variant in SLC39A4 in Bovine Hereditary Zinc Deficiency, Lethal Trait A46: An Animal Model of Acrodermatitis Enteropathica. Genomics 2006, 88, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Dufner-Beattie, J.; Wang, F.; Kuo, Y.M.; Gitschier, J.; Eide, D.; Andrews, G.K. The Acrodermatitis Enteropathica Gene ZIP4 Encodes a Tissue-Specific, Zinc-Regulated Zinc Transporter in Mice. J. Biol. Chem. 2003, 278, 33474–33481. [Google Scholar] [CrossRef]
- Steel, L.; Cousins, R.J. Kinetics of Zinc Absorption by Luminally and Vascularly Perfused Rat Intestine. Am. J. Physiol.-Gastrointest. Liver Physiol. 1985, 248, G46–G53. [Google Scholar] [CrossRef]
- Ford, D. Intestinal and Placental Zinc Transport Pathways. Proc. Nutr. Soc. 2004, 63, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Krebs, N.F. Zinc and Health: Overview of zinc absorption and excretion in the human gastrointestinal tract. J. Nutr. 2000, 130, 1374S–1377S. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, S.; Yorifuji, T.; Nishida, M.; Fukai, K.; Nakano, H. Acrodermatitis Enteropathica: A Hereditary Form of Zinc Deficiency. QJM Int. J. Med. 2021, 114, 270–271. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.M.; Moynahan, E.J. Zinc Deficiency in Acrodermatitis Enteropathica: Multiple Dietary Intolerance Treated with Synthetic Diet. J. R. Soc. Med. 1973, 66, 327–329. [Google Scholar] [CrossRef]
- Jagannathan, V.; Drögemüller, C.; Leeb, T.; Dog Biomedical Variant Database Consortium (DBVDC). A Comprehensive Biomedical Variant Catalogue Based on Whole Genome Sequences of 582 Dogs and Eight Wolves. Anim. Genet. 2019, 50, 695–704. [Google Scholar] [CrossRef]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A Program for Annotating and Predicting the Effects of Single Nucleotide Polymorphisms, SnpEff: SNPs in the Genome of Drosophila Melanogaster Strain W1118; Iso-2; Iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef]
- HomoloGene. Available online: https://www.ncbi.nlm.nih.gov/homologene (accessed on 27 July 2021).
- Basic Local Alignment Search Tool. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 27 July 2021).
- Wang, F.; Kim, B.E.; Dufner-Beattie, J.; Petris, M.J.; Andrews, G.; Eide, D.J. Acrodermatitis Enteropathica Mutations Affect Transport Activity, Localization and Zinc-Responsive Trafficking of the Mouse ZIP4 Zinc Transporter. Hum. Mol. Genet. 2004, 13, 563–571. [Google Scholar] [CrossRef]
- Gross, T.L.; Ihrke, P.J.; Walder, E.J.; Affolter, V.K. Skin Diseases of the Dog and Cat: Clinical and Histopathologic Diagnosis, 2nd ed.; Blackwell Science Ltd.: Oxford, UK, 2005. [Google Scholar]
- Miller, W.; Griffin, C.; Campbell, K. Muller and Kirk’s Small Animal Dermatology, 7th ed.; Saunders: Philadelphia, PA, USA, 2012. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Filtering Step | Variants |
|---|---|
| all variants in the affected cat | 5,518,410 |
| private variants | 56,054 |
| protein-changing private variants | 161 |
| protein-changing private variants in MKLN1 or SLC39A4 | 1 |
| Cats | G/G | G/C | C/C |
|---|---|---|---|
| Cases (n = 3) | - | - | 3 |
| Controls, Turkish Van cats from index family (n = 9) | 5 | 4 | - |
| Controls, other breeds (n = 173) | 173 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiener, S.; Cikota, R.; Welle, M.; Jagannathan, V.; Åhman, S.; Leeb, T. A Missense Variant in SLC39A4 in a Litter of Turkish Van Cats with Acrodermatitis Enteropathica. Genes 2021, 12, 1309. https://doi.org/10.3390/genes12091309
Kiener S, Cikota R, Welle M, Jagannathan V, Åhman S, Leeb T. A Missense Variant in SLC39A4 in a Litter of Turkish Van Cats with Acrodermatitis Enteropathica. Genes. 2021; 12(9):1309. https://doi.org/10.3390/genes12091309
Chicago/Turabian StyleKiener, Sarah, Robert Cikota, Monika Welle, Vidhya Jagannathan, Susanne Åhman, and Tosso Leeb. 2021. "A Missense Variant in SLC39A4 in a Litter of Turkish Van Cats with Acrodermatitis Enteropathica" Genes 12, no. 9: 1309. https://doi.org/10.3390/genes12091309
APA StyleKiener, S., Cikota, R., Welle, M., Jagannathan, V., Åhman, S., & Leeb, T. (2021). A Missense Variant in SLC39A4 in a Litter of Turkish Van Cats with Acrodermatitis Enteropathica. Genes, 12(9), 1309. https://doi.org/10.3390/genes12091309