
Exploring the Contribution to ADHD of Genes Involved in Mendelian Disorders Presenting with Hyperactivity and/or Inattention

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
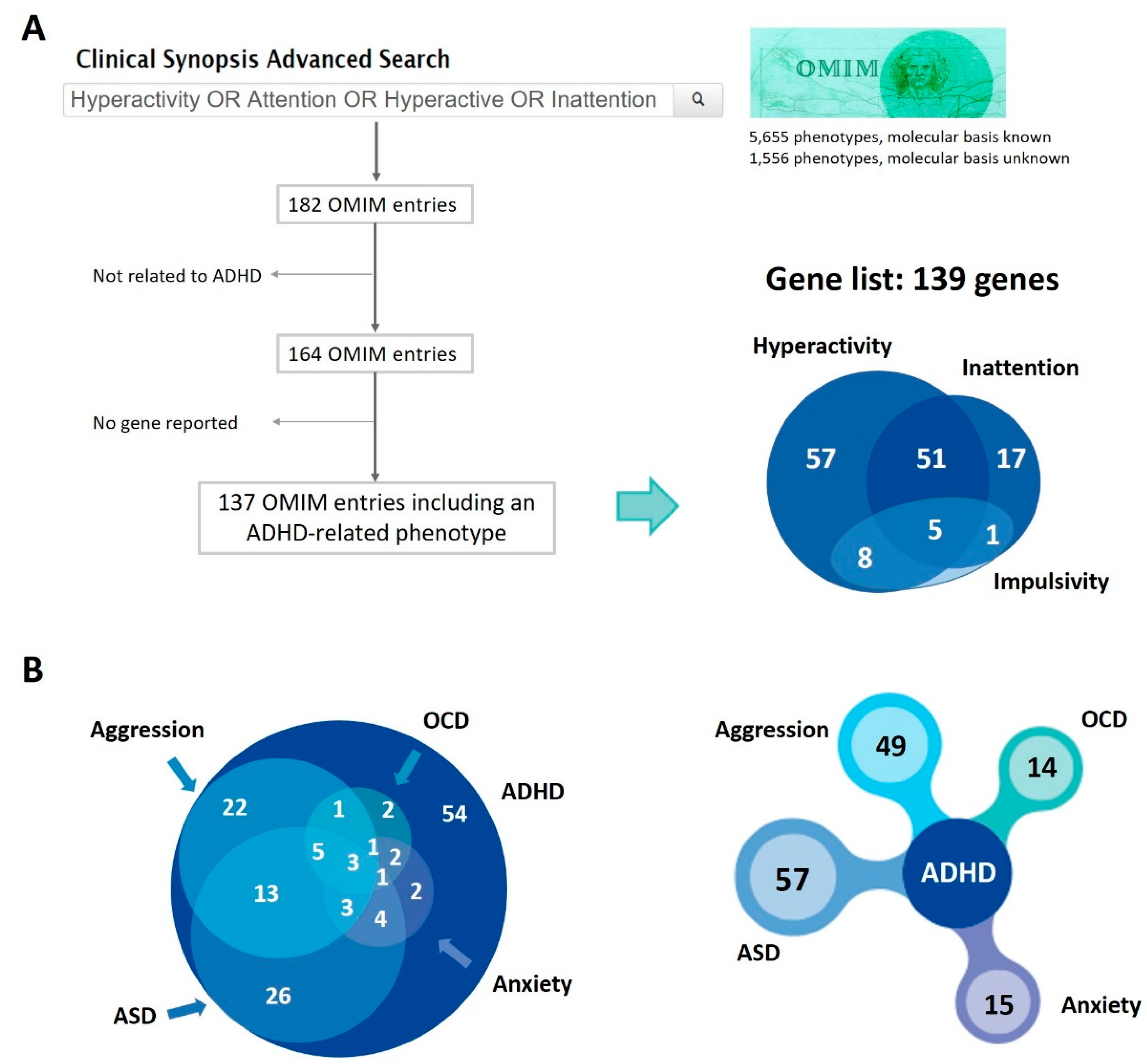
2.1. Gene Selection
2.2. Whole-Exome Sequencing
2.3. Rare Variant Association Analyses
2.4. Common Variant Gene-Based Association Studies
2.5. Gene-Based Analyses and Meta-Analysis with Comorbid Conditions
2.6. MetaXcan
2.7. Effect on Subcortical Brain Volumes
3. Results
3.1. Genes in Mendelian Disorders Presenting with ADHD Symptoms
3.2. Rare Variants in the OMIM Gene List and ADHD Symptomatology in Patients
3.3. Common Variants in the OMIM Gene List and ADHD
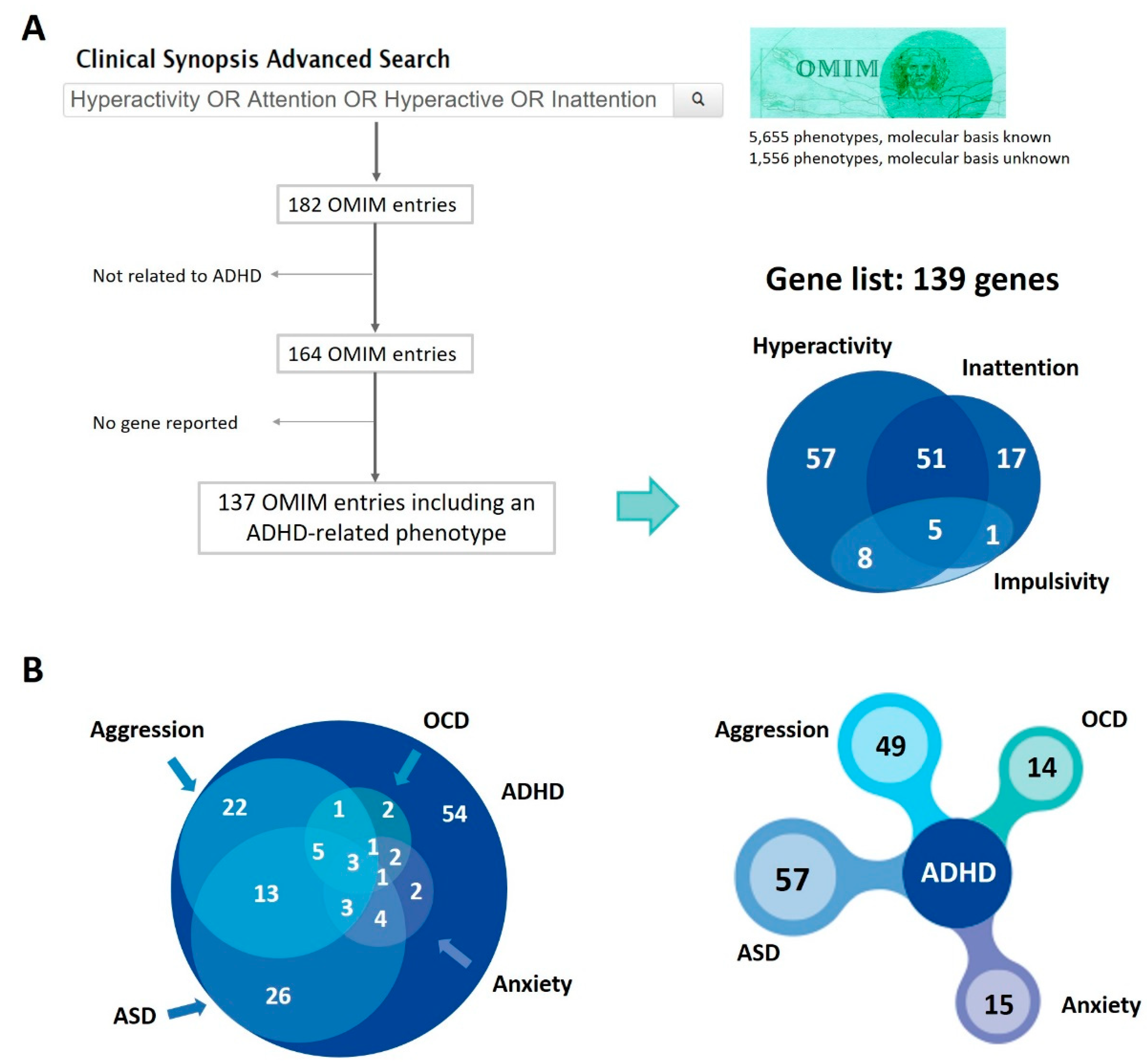
3.4. Contribution to Comorbidities of the Genes Associated with ADHD
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Faraone, S.V.; Asherson, P.; Banaschewski, T.; Biederman, J.; Buitelaar, J.K.; Ramos-Quiroga, J.A.; Rohde, L.A.; Sonuga-Barke, E.J.S.; Tannock, R.; Franke, B. Attention-deficit/hyperactivity disorder. Nat. Rev. Dis. Primers 2015, 1, 15020. [Google Scholar] [CrossRef] [PubMed]
- Reale, L.; Bartoli, B.; Cartabia, M.; Zanetti, M.; Costantino, M.A.; Canevini, M.P.; Termine, C.; Bonati, M.; Conte, S.; Renzetti, V.; et al. Comorbidity prevalence and treatment outcome in children and adolescents with ADHD. Eur. Child Adolesc. Psychiatry 2017, 26, 1443–1457. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Hartman, C.A.; Haavik, J.; Harro, J.; Klungsøyr, K.; Hegvik, T.A.; Wanders, R.; Ottosen, C.; Dalsgaard, S.; Faraone, S.V.; et al. Common psychiatric and metabolic comorbidity of adult attention-deficit/hyperactivity disorder: A population-based cross-sectional study. PLoS ONE 2018, 13, e0204516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demontis, D.; Walters, R.K.; Martin, J.; Mattheisen, M.; Als, T.D.; Agerbo, E.; Baldursson, G.; Belliveau, R.; Bybjerg-Grauholm, J.; Bækvad-Hansen, M.; et al. Discovery of the first genome-wide significant risk loci for attention deficit/hyperactivity disorder. Nat. Genet. 2019, 51, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Grove, J.; Ripke, S.; Als, T.D.; Mattheisen, M.; Walters, R.K.; Won, H.; Pallesen, J.; Agerbo, E.; Andreassen, O.A.; Anney, R.; et al. Identification of common genetic risk variants for autism spectrum disorder. Nat. Genet. 2019, 51, 431–444. [Google Scholar] [CrossRef] [Green Version]
- Tambuyzer, E.; Vandendriessche, B.; Austin, C.P.; Brooks, P.J.; Larsson, K.; Miller Needleman, K.I.; Valentine, J.; Davies, K.; Groft, S.C.; Preti, R.; et al. Therapies for rare diseases: Therapeutic modalities, progress and challenges ahead. Nat. Rev. Drug Discov. 2020, 19, 93–111. [Google Scholar] [CrossRef]
- Ramos-Fuentes, F.; González-Meneses, A.; Ars, E.; Hernández-Jaras, J. Genetic Diagnosis of Rare Diseases: Past and Present. Adv. Ther. 2020, 37, 29–37. [Google Scholar] [CrossRef]
- Corominas, J.; Klein, M.; Zayats, T.; Rivero, O.; Ziegler, G.C.; Pauper, M.; Neveling, K.; Poelmans, G.; Jansch, C.; Svirin, E.; et al. Identification of ADHD risk genes in extended pedigrees by combining linkage analysis and whole-exome sequencing. Mol. Psychiatry 2020, 25, 2047–2057. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [Green Version]
- Rovira, P.; Demontis, D.; Sánchez-Mora, C.; Zayats, T.; Klein, M.; Mota, N.R.; Weber, H.; Garcia-Martínez, I.; Pagerols, M.; Vilar-Ribó, L.; et al. Shared genetic background between children and adults with attention deficit/hyperactivity disorder. Neuropsychopharmacology 2020, 45, 1617–1626. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [Green Version]
- Auton, A.; Abecasis, G.R.; Altshuler, D.M.; Durbin, R.M.; Abecasis, G.R.; Bentley, D.R.; Chakravarti, A.; Clark, A.G.; Donnelly, P.; Eichler, E.E.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willer, C.J.; Li, Y.; Abecasis, G.R. METAL: Fast and efficient meta-analysis of genomewide association scans. Bioinformatics 2010, 26, 2190–2191. [Google Scholar] [CrossRef] [PubMed]
- Otowa, T.; Hek, K.; Lee, M.; Byrne, E.M.; Mirza, S.S.; Nivard, M.G.; Bigdeli, T.; Aggen, S.H.; Adkins, D.; Wolen, A.; et al. Meta-analysis of genome-wide association studies of anxiety disorders. Mol. Psychiatry 2016, 21, 1391–1399. [Google Scholar] [CrossRef]
- Pappa, I.; St Pourcain, B.; Benke, K.; Cavadino, A.; Hakulinen, C.; Nivard, M.G.; Nolte, I.M.; Tiesler, C.M.T.; Bakermans-Kranenburg, M.J.; Davies, G.E.; et al. A genome-wide approach to children’s aggressive behavior: The EAGLE consortium. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2016, 171, 562–572. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Roige, S.; Fontanillas, P.; Elson, S.L.; Gray, J.C.; De Wit, H.; MacKillop, J.; Palmer, A.A. Genome-wide association studies of impulsive personality traits (BIS-11 and UPPS-P) and drug experimentation in up to 22,861 adult research participants identify loci in the CACNA1I and CADM2 genes. J. Neurosci. 2019, 39, 2562–2572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, P.D.; Askland, K.D.; Barlassina, C.; Bellodi, L.; Bienvenu, O.J.; Black, D.; Bloch, M.; Brentani, H.; Burton, C.L.; Camarena, B.; et al. Revealing the complex genetic architecture of obsessive-compulsive disorder using meta-analysis. Mol. Psychiatry 2018, 23, 1181–1188. [Google Scholar] [CrossRef]
- Barbeira, A.N.; Dickinson, S.P.; Bonazzola, R.; Zheng, J.; Wheeler, H.E.; Torres, J.M.; Torstenson, E.S.; Shah, K.P.; Garcia, T.; Edwards, T.L.; et al. Exploring the phenotypic consequences of tissue specific gene expression variation inferred from GWAS summary statistics. Nat. Commun. 2018, 9, 1825. [Google Scholar] [CrossRef]
- Lonsdale, J.; Thomas, J.; Salvatore, M.; Phillips, R.; Lo, E.; Shad, S.; Hasz, R.; Walters, G.; Garcia, F.; Young, N.; et al. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef]
- Hibar, D.P.; Stein, J.L.; Renteria, M.E.; Arias-Vasquez, A.; Desrivières, S.; Jahanshad, N.; Toro, R.; Wittfeld, K.; Abramovic, L.; Andersson, M.; et al. Common genetic variants influence human subcortical brain structures. Nature 2015, 520, 224–229. [Google Scholar] [CrossRef] [Green Version]
- Zhang-James, Y.; Faraone, S.V. Genetic architecture for human aggression: A study of gene–phenotype relationship in OMIM. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2016, 171, 641–649. [Google Scholar] [CrossRef]
- Hoogman, M.; Muetzel, R.; Guimaraes, J.P.; Shumskaya, E.; Mennes, M.; Zwiers, M.P.; Jahanshad, N.; Sudre, G.; Wolfers, T.; Earl, E.A.; et al. Brain imaging of the cortex in ADHD: A coordinated analysis of large-scale clinical and population-based samples. Am. J. Psychiatry 2019, 176, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Maltezos, S.; Horder, J.; Coghlan, S.; Skirrow, C.; O’Gorman, R.; Lavender, T.J.; Mendez, M.A.; Mehta, M.; Daly, E.; Xenitidis, K.; et al. Glutamate/glutamine and neuronal integrity in adults with ADHD: A proton MRS study. Transl. Psychiatry 2014, 4, e373. [Google Scholar] [CrossRef] [PubMed]
- Lesch, K.P.; Merker, S.; Reif, A.; Novak, M. Dances with black widow spiders: Dysregulation of glutamate signalling enters centre stage in ADHD. Eur. Neuropsychopharmacol. 2013, 23, 479–491. [Google Scholar] [CrossRef]
- Cheng, J.; Liu, A.; Shi, M.Y.; Yan, Z. Disrupted glutamatergic transmission in prefrontal cortex contributes to behavioral abnormality in an animal model of ADHD. Neuropsychopharmacology 2017, 42, 2096–2104. [Google Scholar] [CrossRef]
- Sorokina, A.M.; Saul, M.; Goncalves, T.M.; Gogola, J.V.; Majdak, P.; Rodriguez-Zas, S.L.; Rhodes, J.S. Striatal transcriptome of a mouse model of ADHD reveals a pattern of synaptic remodeling. PLoS ONE 2018, 13, e0201553. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wu, D.; Xu, Q.; You, L.; Zhu, J.; Wang, J.; Liu, Z.; Yang, L.; Tong, M.; Hong, Q.; et al. The protective effect and potential mechanism of NRXN1 on learning and memory in ADHD rat models. Exp. Neurol. 2021, 344, 113806. [Google Scholar] [CrossRef]
- Ziegler, G.C.; Ehlis, A.C.; Weber, H.; Vitale, M.R.; Zöller, J.E.M.; Ku, H.P.; Schiele, M.A.; Kürbitz, L.I.; Romanos, M.; Pauli, P.; et al. A common cdh13 variant is associated with low agreeableness and neural responses to working memory tasks in adhd. Genes 2021, 12, 1356. [Google Scholar] [CrossRef]
- Vink, J.M.; Treur, J.L.; Pasman, J.A.; Schellekens, A. Investigating genetic correlation and causality between nicotine dependence and ADHD in a broader psychiatric context. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2020, 186, 423–429. [Google Scholar] [CrossRef]
- Bock, J.; Breuer, S.; Poeggel, G.; Braun, K. Early life stress induces attention-deficit hyperactivity disorder (ADHD)-like behavioral and brain metabolic dysfunctions: Functional imaging of methylphenidate treatment in a novel rodent model. Brain Struct. Funct. 2017, 222, 765–780. [Google Scholar] [CrossRef] [Green Version]
- Zhang-James, Y.; Fernàndez-Castillo, N.; Hess, J.L.; Malki, K.; Glatt, S.J.; Cormand, B.; Faraone, S.V. An integrated analysis of genes and functional pathways for aggression in human and rodent models. Mol. Psychiatry 2019, 24, 1655–1667. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Bacanu, S.A.; Yu, H.; Zhao, Z.; Jia, P.; Kendler, K.S.; Kranzler, H.R.; Gelernter, J.; Farrer, L.; Minica, C.; et al. Genetic Relationship between Schizophrenia and Nicotine Dependence. Sci. Rep. 2016, 6, 25671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ovenden, E.S.; Drögemöller, B.I.; Van Der Merwe, L.; Chiliza, B.; Asmal, L.; Emsley, R.A.; Warnich, L. Fine-mapping of antipsychotic response genome-wide association studies reveals novel regulatory mechanisms. Pharmacogenomics 2017, 18, 105–120. [Google Scholar] [CrossRef]
- Wittkowski, K.M.; Sonakya, V.; Bigio, B.; Tonn, M.K.; Shic, F.; Ascano, M.; Nasca, C.; Gold-Von Simson, G. A novel computational biostatistics approach implies impaired dephosphorylation of growth factor receptors as associated with severity of autism. Transl. Psychiatry 2014, 4, e354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowers, J.M.; Konopka, G. The role of the FOXP family of transcription factors in ASD. Dis. Markers 2012, 33, 251–260. [Google Scholar] [CrossRef]
- Li, W.; Pozzo-Miller, L. Dysfunction of the corticostriatal pathway in autism spectrum disorders. J. Neurosci. Res. 2020, 98, 2130–2147. [Google Scholar] [CrossRef]
- Zhao, J.; Chen, C.; Bell, R.L.; Qing, H.; Lin, Z. Identification of HIVEP2 as a dopaminergic transcription factor related to substance use disorders in rats and humans. Transl. Psychiatry 2019, 9, 247. [Google Scholar] [CrossRef]
- Murphy, C.E.; Kondo, Y.; Walker, A.K.; Rothmond, D.A.; Matsumoto, M.; Shannon Weickert, C. Regional, cellular and species difference of two key neuroinflammatory genes implicated in schizophrenia. Brain. Behav. Immun. 2020, 88, 826–839. [Google Scholar] [CrossRef]
- Murphy, C.E.; Lawther, A.J.; Webster, M.J.; Asai, M.; Kondo, Y.; Matsumoto, M.; Walker, A.K.; Weickert, C.S. Nuclear factor kappa B activation appears weaker in schizophrenia patients with high brain cytokines than in non-schizophrenic controls with high brain cytokines. J. Neuroinflamm. 2020, 17, 215. [Google Scholar] [CrossRef]
- Takao, K.; Kobayashi, K.; Hagihara, H.; Ohira, K.; Shoji, H.; Hattori, S.; Koshimizu, H.; Umemori, J.; Toyama, K.; Nakamura, H.K.; et al. Deficiency of schnurri-2, an MHC enhancer binding protein, induces mild chronic inflammation in the brain and confers molecular, neuronal, and behavioral phenotypes related to schizophrenia. Neuropsychopharmacology 2013, 38, 1409–1425. [Google Scholar] [CrossRef]
- Takagi, T.; Jin, W.; Taya, K.; Watanabe, G.; Mori, K.; Ishii, S. Schnurri-2 mutant mice are hypersensitive to stress and hyperactive. Brain Res. 2006, 1108, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.J.; Gitton, Y. The untold stories of the speech gene, the FOXP2 cancer gene. Genes Cancer 2018, 9, 818–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickey, S.L.; Berto, S.; Konopka, G. Chromatin Decondensation by FOXP2 Promotes Human Neuron Maturation and Expression of Neurodevelopmental Disease Genes. Cell Rep. 2019, 27, 1699–1711.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lüffe, T.M.; D’Orazio, A.; Bauer, M.; Gioga, Z.; Schoeffler, V.; Lesch, K.P.; Romanos, M.; Drepper, C.; Lillesaar, C. Increased locomotor activity via regulation of GABAergic signalling in foxp2 mutant zebrafish—implications for neurodevelopmental disorders. Transl. Psychiatry 2021, 11, 529. [Google Scholar] [CrossRef]

{kind=link}
| Gene Symbol | OMIM | ADHD Phenotype in Mendelian Disorder | Genomic Coordinates (hg38) | Number of Rare Variants Identified | p-Value | Adjusted p-Value |
|---|---|---|---|---|---|---|
| KIF11 | #152950 | Hyperactivity/inattention | 10:92593068-92655395 | 1 (10:92613567–A) | 4.81 × 10−6 | 2.43 × 10−4 |
| CRBN | #607417 | Hyperactivity/inattention | 3:3149633-3179717 | 3 (3:3150954–G; 3:3174156–T; rs201449042) | 3.03 × 10−6 | 2.43 × 10−4 |
| WAC | #616708 | Hyperactivity/inattention | 10:28532588-28623112 | 1 (rs201855730) | 1.03 × 10−4 | 3.45 × 10−3 |
| Gene Symbol | Mendelian Disorder | ADHD Phenotype in Mendelian Disorder | Comorbid Psychiatric Phenotypes in Mendelian Disorder | ADHD Gene-Based p-Value | Adjusted p-Value * | Significant Gene-Based Association of Comorbidities |
|---|---|---|---|---|---|---|
| MANBA | #248510 | Hyperactivity | Aggression | 5.99 × 10−8 | 6.65 × 10−6 | |
| UQCC2 | #615824 | Hyperactivity | ASD/Aggression | 5.88 × 10−4 | 0.0326 | |
| HIVEP2 | #616977 | Hyperactivity/impulsivity | ASD/Anxiety | 1.10 × 10−3 | 0.0407 | ASD/Anxiety |
| FOXP1 | #613670 | Hyperactivity | ASD/Aggression/OCD | 2.43 × 10−3 | 0.0674 | ASD/Aggression |
| KANSL1 | #610443 | Hyperactivity/impulsivity | ASD/Anxiety | 3.56 × 10−3 | 0.0790 | ASD |
| AUH | #250950 | Short attention span | - | 4.63 × 10−3 | 0.0857 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernàndez-Castillo, N.; Cabana-Domínguez, J.; Kappel, D.B.; Torrico, B.; Weber, H.; Lesch, K.-P.; Lao, O.; Reif, A.; Cormand, B. Exploring the Contribution to ADHD of Genes Involved in Mendelian Disorders Presenting with Hyperactivity and/or Inattention. Genes 2022, 13, 93. https://doi.org/10.3390/genes13010093
Fernàndez-Castillo N, Cabana-Domínguez J, Kappel DB, Torrico B, Weber H, Lesch K-P, Lao O, Reif A, Cormand B. Exploring the Contribution to ADHD of Genes Involved in Mendelian Disorders Presenting with Hyperactivity and/or Inattention. Genes. 2022; 13(1):93. https://doi.org/10.3390/genes13010093
Chicago/Turabian StyleFernàndez-Castillo, Noèlia, Judit Cabana-Domínguez, Djenifer B. Kappel, Bàrbara Torrico, Heike Weber, Klaus-Peter Lesch, Oscar Lao, Andreas Reif, and Bru Cormand. 2022. "Exploring the Contribution to ADHD of Genes Involved in Mendelian Disorders Presenting with Hyperactivity and/or Inattention" Genes 13, no. 1: 93. https://doi.org/10.3390/genes13010093
APA StyleFernàndez-Castillo, N., Cabana-Domínguez, J., Kappel, D. B., Torrico, B., Weber, H., Lesch, K.-P., Lao, O., Reif, A., & Cormand, B. (2022). Exploring the Contribution to ADHD of Genes Involved in Mendelian Disorders Presenting with Hyperactivity and/or Inattention. Genes, 13(1), 93. https://doi.org/10.3390/genes13010093