
Identification of Cdk8 and Cdkn2d as New Prame-Target Genes in 2C-like Embryonic Stem Cells

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
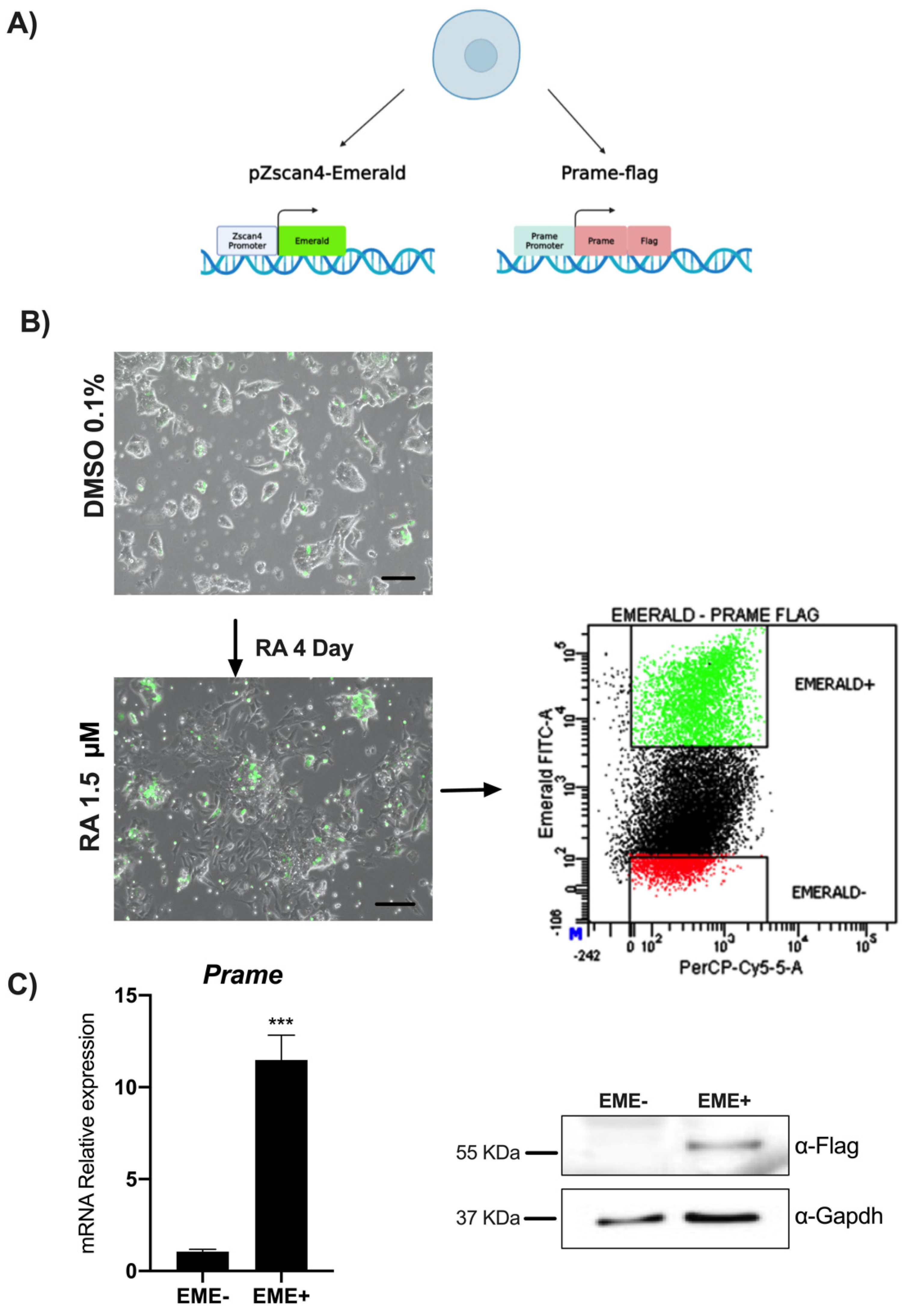
2.2. Flow Cytometry and Sorting of ESpZscan4-Em/Prame-FLAG Transgenic Cell Lines
2.3. Western Blot Analysis
2.4. qPCR Analysis
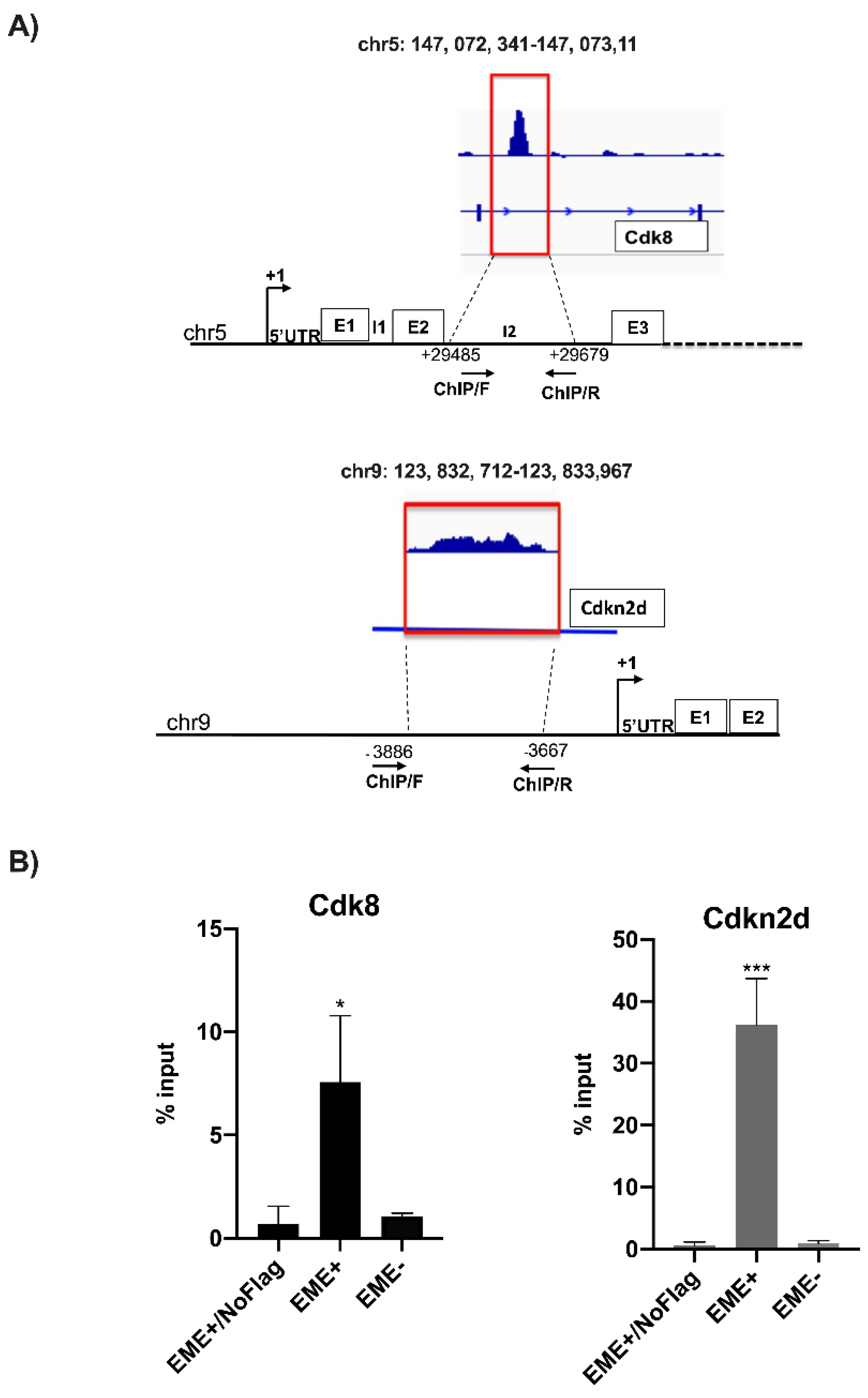
2.5. ChIP Analysis
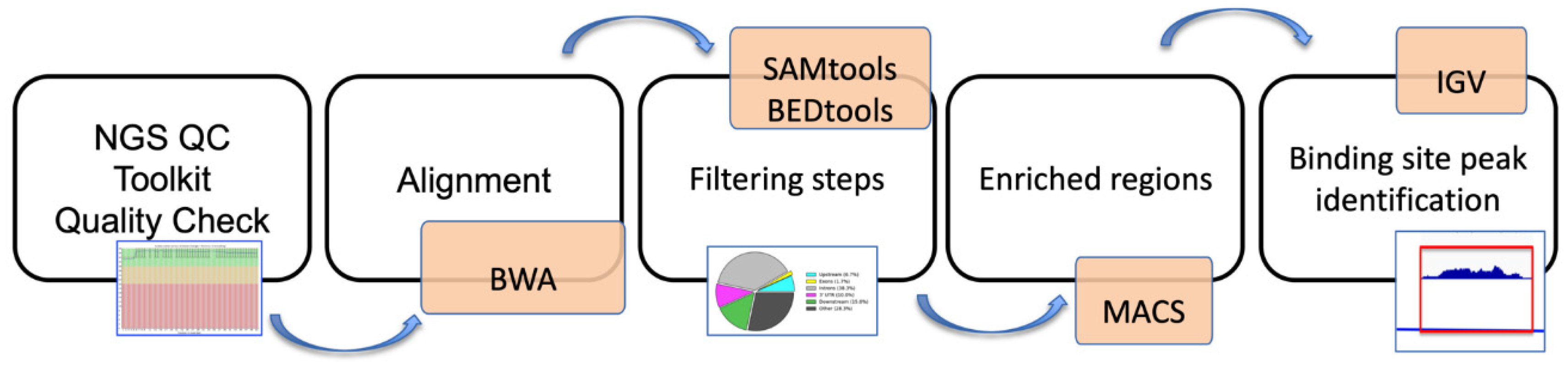
2.6. ChIP-Sequencing, Mapping, and Peak Analysis
2.7. Statistical Analysis
3. Results
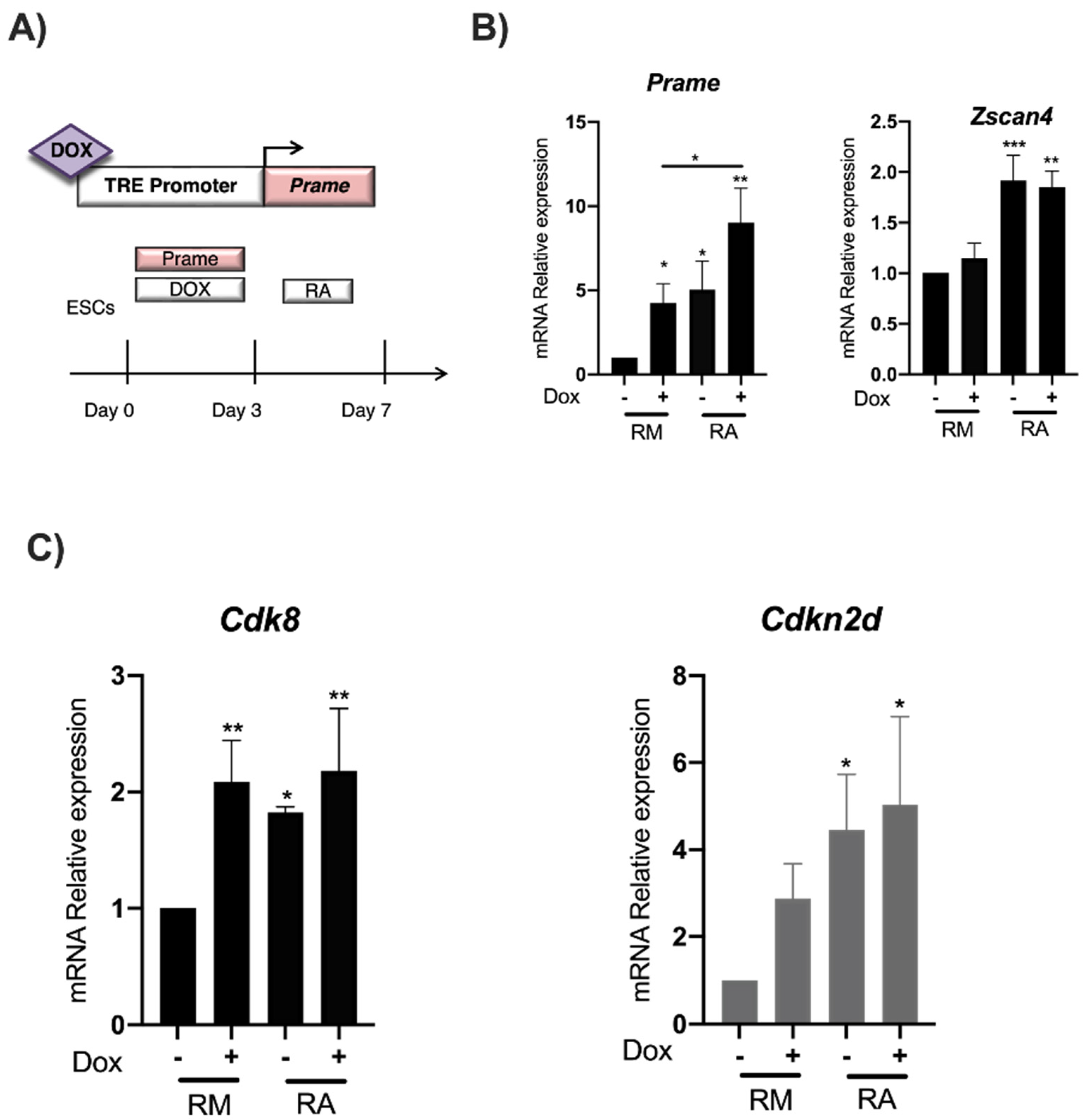
3.1. RA induces Zscan4 Metastate and Prame Expression in ESpZscan4-EME/PRAME-FLAG
3.2. Identification of Prame Target Genes by ChIP-Seq Analysis
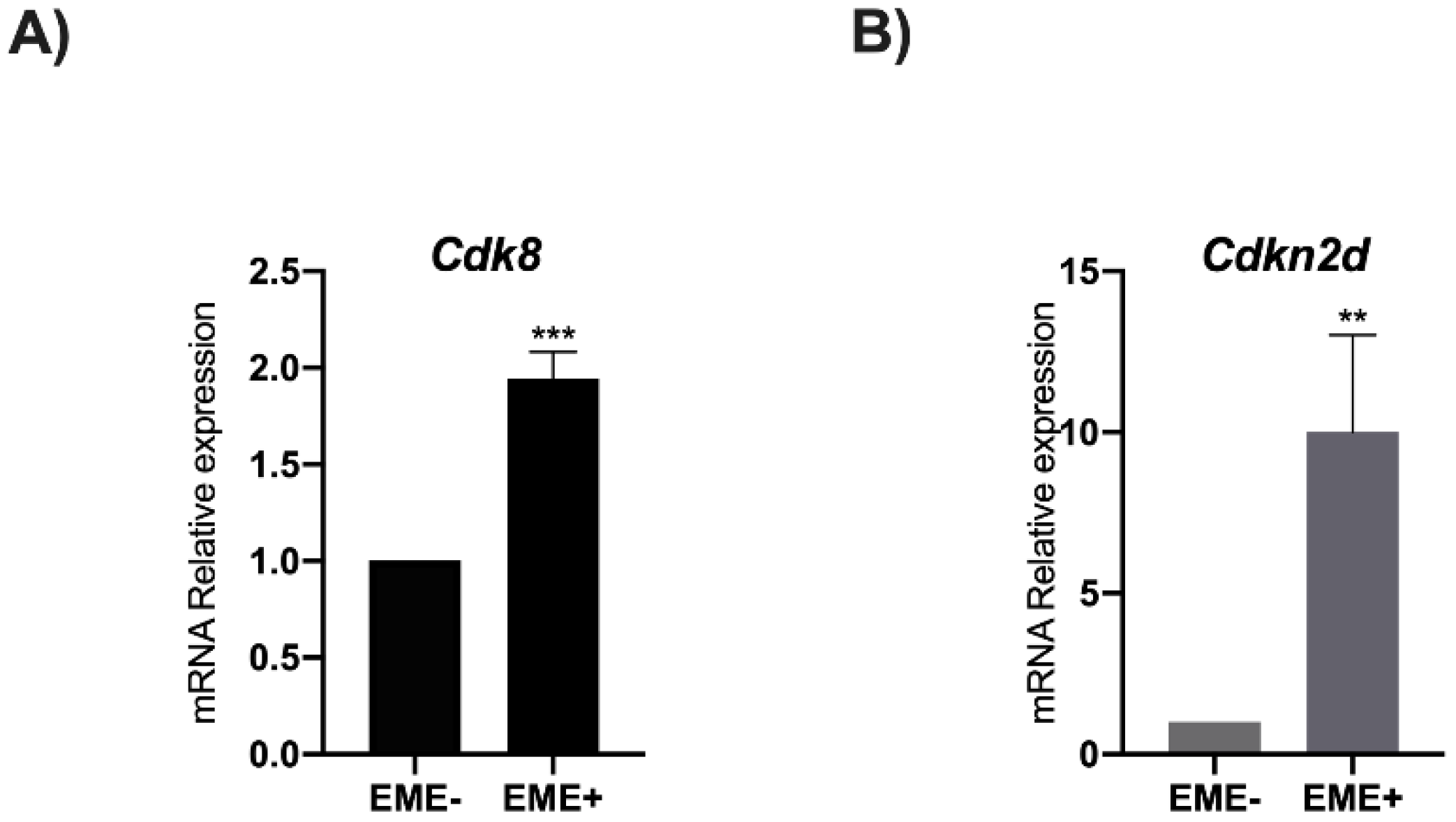
3.3. RA Induces Cdk8 and Cdkn2d Expression
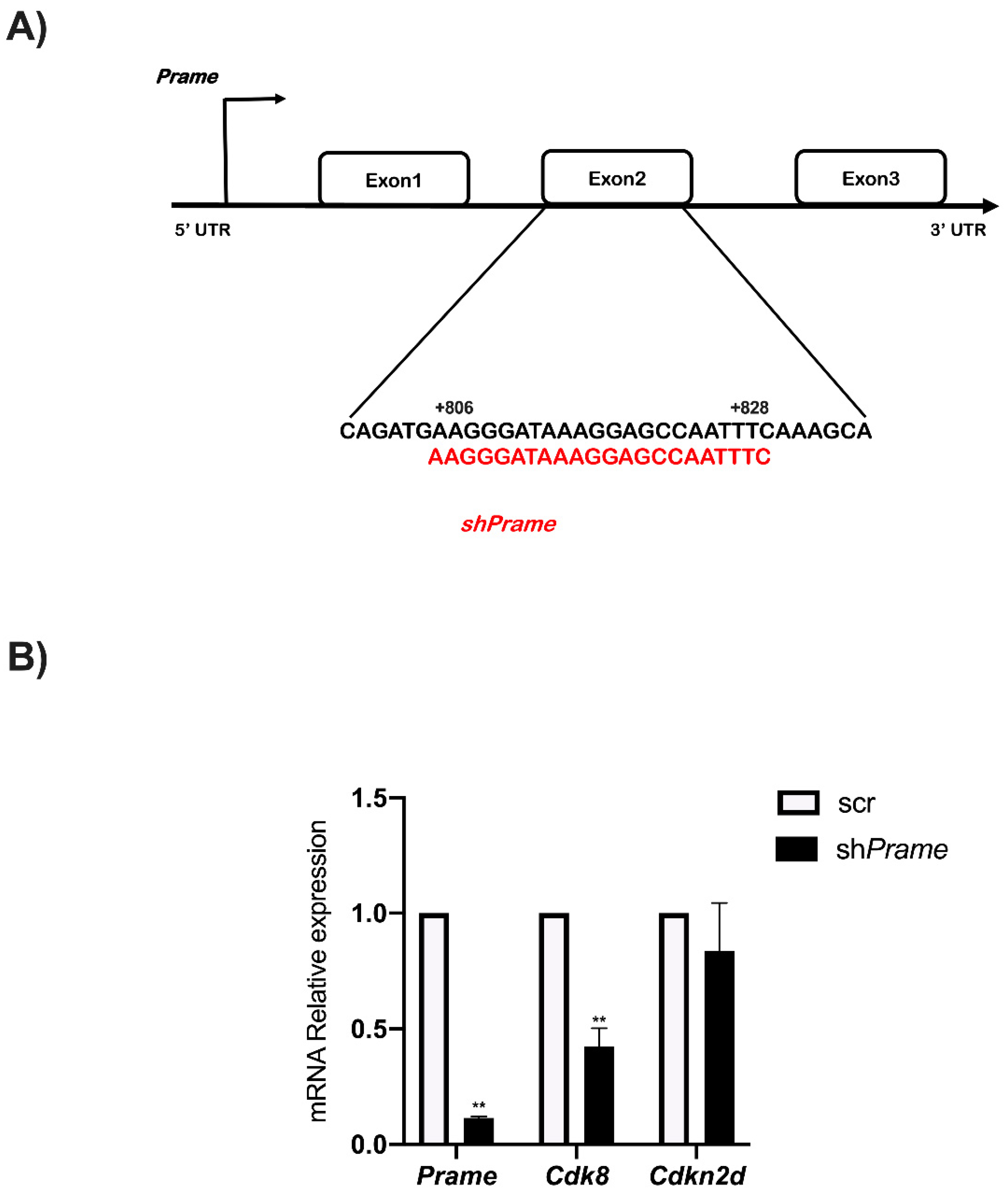
3.4. Prame Enhances Cdk8 and Cdkn2d Expression in ESGm12794cp2Lox Transgenic Cells
3.5. Prame Controls Cdk8 Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Madissoon, E.; Töhönen, V.; Vesterlund, L.; Katayama, S.; Unneberg, P.; Inzunza, J.; Hovatta, O.; Kere, J. Differences in gene expression between mouse and human for dynamically regulated genes in early embryo. PLoS ONE 2014, 9, 102949. [Google Scholar] [CrossRef]
- Li, J.; Yin, J.; Zhong, J.; Yang, Z.; Tang, A.; Li, S. Clinicopathological and Prognostic Significance of PRAME Overexpression in Human Cancer: A Meta-Analysis. Biomed. Res. Int. 2020, 2020, 8828579. [Google Scholar] [CrossRef] [PubMed]
- Graf, U.; Casanova, E.A.; Wyck, S.; Dalcher, D.; Gatti, M.; Vollenweider, E.; Okoniewski, M.J.; Weber, F.A.; Patel, S.S.; Schmid, M.W.; et al. Pramel7 mediates ground-state pluripotency through proteasomal-epigenetic combined pathways. Nat. Cell Biol. 2017, 19, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Amano, T.; Hirata, T.; Falco, G.; Monti, M.; Sharova, L.V.; Amano, M.; Sheer, S.; Hoang, H.G.; Piao, Y.; Stagg, C.A.; et al. Zscan4 restores the developmental potency of embryonic stem cells. Nat. Commun. 2013, 4, 1966. [Google Scholar] [CrossRef] [PubMed]
- Natale, C.F.; Angrisano, T.; Pistelli, L.; Falco, G.; Calabrò, V.; Netti, P.A.; Ventre, M. Topographic Cues Impact on Embryonic Stem Cell Zscan4-Metastate. Front. Bioeng. Biotechnol. 2020, 8, 178. [Google Scholar] [CrossRef]
- Falco, G.; Lee, S.M.; Stanghellini, I.; Bassey, U.C.; Hamatani, T.; Ko, M.S.H. Zscan4: A novel gene expressed exclusively in late 2-cell embryos and embryonic stem cells. Dev. Biol. 2007, 307, 539–550. [Google Scholar] [CrossRef]
- Dahl, J.A.; Jung, I.; Aanes, H.; Greggains, G.D.; Manaf, A.; Lerdrup, M.; Li, G.; Kuan, S.; Li, B.; Lee, A.Y.; et al. Broad histone H3K4me3 domains in mouse oocytes modulate maternal-to-zygotic transition. Nature 2016, 537, 548–552. [Google Scholar] [CrossRef]
- Tagliaferri, D.; De Angelis, M.T.; Russo, N.A.; Marotta, M.; Ceccarelli, M.; Del Vecchio, L.; De Felice, M.; Falco, G. Retinoic Acid Specifically Enhances Embryonic Stem Cell Metastate Marked by Zscan4. PLoS ONE 2016, 11, 0147683. [Google Scholar] [CrossRef]
- Cerulo, L.; Tagliaferri, D.; Marotta, P.; Zoppoli, P.; Russo, F.; Mazio, C.; De Felice, M.; Ceccarelli, M.; Falco, G. Identification of a novel gene signature of ES cells self-renewal fluctuation through system-wide analysis. PLoS ONE 2014, 9, 83235. [Google Scholar]
- Macfarlan, T.S.; Gifford, W.D.; Driscoll, S.; Lettieri, K.; Rowe, H.M.; Bonanomi, D.; Firth, A.; Singer, O.; Trono, D.; Pfaff, S.L.; et al. Embryonic stem cell potency fluctuates with endogenous retrovirus activity. Nature 2012, 487, 57–63. [Google Scholar] [CrossRef]
- Epping, M.T.; Wang, L.; Edel, M.J.; Carlée, L.; Hernandez, M.; Bernards, R. The human tumor antigen PRAME is a dominant repressor of retinoic acid receptor signaling. Cell 2005, 122, 835–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epping, M.T.; Wang, L.; Plumb, J.A.; Lieb, M.; Gronemeyer, H.; Brown, R.; Bernards, R. A functional genetic screen identifies retinoic acid signaling as a target of histone deacetylase inhibitors. Proc. Natl. Acad. Sci. USA 2007, 104, 17777–17782. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, G.; Tagliaferri, D.; Fusco, S.; Cirillo, C.; De Martino, I.; Addeo, M.; Mazzone, P.; Russo, N.A.; Natale, F.; Cardoso, M.C.; et al. A novel member of Prame family, Gm12794c, counteracts retinoic acid differentiation through the methyltransferase activity of PRC2. Cell Death Differ. 2020, 27, 345–362. [Google Scholar] [CrossRef] [PubMed]
- Oehler, V.G.; Guthrie, K.A.; Cummings, C.L.; Sabo, K.; Wood, B.L.; Gooley, T.; Yang, T.; Epping, M.T.; Shou, Y.; Pogoso-va-Agadjanyan, E.; et al. The preferentially expressed antigen in melanoma (PRAME) inhibits myeloid differentiation in normal hematopoietic and leukemic progenitor cells. Blood 2009, 114, 3299–3308. [Google Scholar] [CrossRef] [PubMed]
- Wadelin, F.; Fulton, J.; McEwan, P.A.; Spriggs, K.A.; Emsley, J.; Heery, D.M. Leucine-rich repeat protein PRAME: Expression, potential functions and clinical implications for leukaemia. Mol. Cancer 2010, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Adler, A.S.; McCleland, M.L.; Truong, T.; Lau, S.; Modrusan, Z.; Soukup, T.M.; Roose-Girma, M.; Blackwood, E.M.; Firestein, R. CDK8 maintains tumor dedifferentiation and embryonic stem cell pluripotency. Cancer Res. 2012, 72, 2129–2139. [Google Scholar] [CrossRef]
- Kim, W.Y.; Sharpless, N.E. The regulation of INK4/ARF in cancer and aging. Cell 2006, 127, 265–275. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, Y.; Yin, B.; Li, J.; Liu, G.; Xu, W.; Tang, S. Differential expression of Axin1, Cdc25c and Cdkn2d mRNA in 2-cell stage mouse blastomeres. Zygote 2012, 20, 305–310. [Google Scholar] [CrossRef]
- Troiano, A.; Pacelli, C.; Ruggieri, V.; Scrima, R.; Addeo, M.; Agriesti, F.; Lucci, V.; Cavaliere, G.; Mollica, M.P.; Caterino, M. ZSCAN4+ mouse embryonic stem cells have an oxidative and flexible metabolic profile. EMBO Rep. 2020, 21, 48942. [Google Scholar] [CrossRef]
- Vivo, M.; Fontana, R.; Ranieri, M.; Capasso, G.; Angrisano, T.; Pollice, A.; Calabrò, V.; La Mantia, G. p14ARF interacts with the focal adhesion kinase and protects cells from anoikis. Oncogene 2017, 36, 4913–4928. [Google Scholar] [CrossRef] [PubMed]
- Montano, E.; Pollice, A.; Lucci, V.; Falco, G.; Affinito, O.; La Mantia, G.; Vivo, M.; Angrisano, T. Pancreatic Progenitor Commitment Is Marked by an Increase in Ink4a/Arf Expression. Biomolecules 2021, 11, 1124. [Google Scholar] [CrossRef] [PubMed]
- Angrisano, T.; Pero, R.; Brancaccio, M.; Coretti, L.; Florio, E.; Pezone, A.; Calabrò, V.; Falco, G.; Keller, S.; Lembo, F.; et al. Cyclical DNA Methylation and Histone Changes Are Induced by LPS to Activate COX-2 in Human Intestinal Epithelial Cells. PLoS ONE 2016, 11, 0156671. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef]
- Huang, W.; Loganantharaj, R.; Schroeder, B.; Fargo, D.; Li, L. PAVIS: A tool for peak annotation and visualization. Bioinformatics 2013, 29, 3097–3099. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Deng, J.M.; Zhang, Z.; Behringer, R.; De Crombrugghe, B.; Maity, S.N. The B subunit of the CCAAT box binding transcription factor complex (CBF/NF-Y) is essential for early mouse development and cell proliferation. Cancer Res. 2003, 63, 8167–8172. [Google Scholar]
- Grskovic, M.; Chaivorapol, C.; Gaspar-Maia, A.; Li, H.; Ramalho-Santos, M. Systematic identification of cis-regulatory sequences active in mouse and human embryonic stem cells. PLoS ONE 2007, 3, 145. [Google Scholar] [CrossRef]
- Gurtner, A.; Manni, I.; Fuschi, P.; Mantovani, R.; Guadagni, F.; Sacchi, A.; Piaggio, G. Requirement for down-regulation of the CCAAT-binding activity of the NF-Y transcription factor during skeletal muscle differentiation. Mol. Biol. Cell 2003, 14, 2706–2715. [Google Scholar] [CrossRef]
- Costessi, A.; Mahrour, N.; Tijchon, E.; Stunnenberg, R.; Stoel, M.A.; Jansen, P.W.; Sela, D.; Martin-Brown, S.; Washburn, M.P.; Florens, L.; et al. The tumor antigen PRAME is a subunit of a Cul2 ubiquitin ligase and associates with active NFY promoters. EMBO J. 2011, 30, 3786–3798. [Google Scholar] [CrossRef]
- Wang, P.J.; McCarrey, J.R.; Yang, F.; Page, D.C. An abundance of X-linked genes expressed in spermatogonia. Nat. Genet. 2001, 27, 422–426. [Google Scholar] [CrossRef]
- Dadé, S.; Callebaut, I.; Mermillod, P.; Monget, P. Identification of a new expanding family of genes characterized by atypical LRR domains. Localization of a cluster preferentially expressed in oocyte. FEBS Lett. 2003, 555, 533–538. [Google Scholar] [CrossRef]
- Bortvin, A.; Eggan, K.; Skaletsky, H.; Akutsu, H.; Berry, D.L.; Yanagimachi, R.; Page, D.C.; Jaenisch, R. Incomplete reactivation of Oct4-related genes in mouse embryos cloned from somatic nuclei. Development 2003, 130, 1673–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cazzato, G.; Mangialardi, K.; Falcicchio, G.; Colagrande, A.; Ingravallo, G.; Arezzo, F.; Giliberti, G.; Trilli, I.; Loizzi, V.; Lettini, T.; et al. Preferentially Expressed Antigen in Melanoma (PRAME) and Human Malignant Melanoma: A Retrospective Study. Genes 2022, 13, 545. [Google Scholar] [CrossRef] [PubMed]
- Cinelli, P.; Casanova, E.A.; Uhlig, S.; Lochmatter, P.; Matsuda, T.; Yokota, T.; Rülicke, T.; Ledermann, B.; Bürki, K. Expression profiling in transgenic FVB/N embryonic stem cells overexpressing STAT3. BMC Dev. Biol. 2008, 8, 57. [Google Scholar] [CrossRef]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem cells, cancer, and cancer stem cells. Nature 2001, 414, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Menzl, I.; Zhang, T.; Berger-Becvar, A.; Grausenburger, R.; Heller, G.; Prchal-Murphy, M.; Edlinger, L.; Knab, M.V.; Uras, I.Z.; Grundschober, E.; et al. A kinase-independent role for CDK8 in BCR-ABL1+ leukemia. Nat. Commun. 2019, 10, 4741. [Google Scholar] [CrossRef]
- Sharko, A.C.; Lim, C.K.; McDermott, M.S.J.; Hennes, C.; Philavong, K.P.; Aiken, T.; Tatarskiy, V.V.; Roninson, I.B.; Broude, E.V. The Inhibition of CDK8/19 Mediator Kinases Prevents the Development of Resistance to EGFR-Targeting Drugs. Cells 2021, 10, 144. [Google Scholar] [CrossRef]
- Antonova, O.; Rukova, B.; Mladenov, B.; Rangelov, S.; Hammoudeh, Z.; Nesheva, D.; Staneva, R.; Spasova, V.; Grigorov, E.; Hadjidekova, S.; et al. Expression profiling of muscle-invasive and non-invasive bladder tumors for biomarkers identification related to drug resistance, sensitivity, and tumor progression. Biotechnol. Biotechnol. Equip. 2020, 34, 506–514. [Google Scholar] [CrossRef]
- Thoma, O.M.; Neurath, M.F.; Waldner, M.J. Cyclin-Dependent Kinase Inhibitors and Their Therapeutic Potential in Colorectal Cancer Treatment. Front. Pharm. 2021, 12, 757120. [Google Scholar] [CrossRef]
- Spear, J.M.; Lu, Z.; Russu, W.A. Pharmacological Inhibition of CDK8 in Triple-Negative Breast Cancer Cell Line MDA-MB-468 Increases E2F1 Protein, Induces Phosphorylation of STAT3 and Apoptosis. Molecules 2020, 25, 5728. [Google Scholar] [CrossRef]
- Xi, M.; Chen, T.; Wu, C.; Gao, X.; Wu, Y.; Luo, X.; Du, K.; Yu, L.; Cai, T.; Shen, R.; et al. CDK8 as a therapeutic target for cancers and recent developments in discovery of CDK8 inhibitors. Eur. J. Med. Chem. 2019, 164, 77–91. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence Fw (5′-3′) | Sequence Rv (5′-3′) | 1 Method |
|---|---|---|---|
| Cdkn2d | CCACCGGTATCCACTATGCT | GCACAGGACTAGTACCGGAG | qPRC |
| Cdk8 | TCCAGTGGTTGTAACATTCTGGT | TTCTGCAAATATACACCCTATAGCC | qPRC |
| Prame | TGCTGCCAAATTCCTTTCTC | GAGAGTTGGCAGCGATTCAT | qPRC |
| Gapdh | AATGGTGAAGGTCGGTGTG | GAAGATGGTGATGGGCTTCC | qPRC |
| Cdkn2d | TCCTCATGCTGGTTCTGTGT | AGGAGGGAAAACAAGGGCTT | ChIP |
| Cdk8 | TTAAATGAGACATGGGCGCG | GGAGTTTACCACCCGCTTTG | ChIP |
| Gene | Category | Chr | Description |
|---|---|---|---|
| Cdk8 | Intron | 5 | Cell division protein kinase 8 |
| Cdkn2d | Upstream, Downstream | 9 | Cyclin-dependent kinase 4 inhibitor D |
| uc009gks.1 | Upstream, Intron, 3′UTR, Downstream | 7 | Hypothetical protein LOC72244 |
| St6galnac1 | Intron | 11 | α-N-Acetyl-galactosaminide |
| Stard13 | Intron | 5 | stAR-related lipid transfer protein 13 isoform |
| Mir26b | Downstream | 1 | Mus musculus microRNA 26 b (Mir26b), microRNA |
| 5330429B09Rik | Downstream | 8 | Subname: Full = putative uncharacterized protein |
| Gm15319 | Intron | 8 | Testis-specific gene with ankyrin repeats |
| Gm3168 | 3′UTR | 8 | Subname: Full = Gag protein |
| AK053193 | Downstream | 10 | Mus musculus 9,5 days embryo parthenogenote cDNA, RIKEN full |
| Mir715 | Downstream | 17 | Mus musculus microRNA 715 (Mir715), microRNA |
| Zc3h7a | Intron | 16 | Zinc Finger CCCH-type containing 7A |
| Filip1l | Intron | 16 | Filamin A-interacting protein 1-like isoform 1 |
| AK041614 | Downstream | 15 | Mus musculus 3 days neonate thymus cDNA, RIKEN full-length enriched library |
| Gm10406 | Intron | 14 | Alpha7-takusan |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucci, V.; De Marino, E.; Tagliaferri, D.; Amente, S.; Pollice, A.; Calabrò, V.; Vivo, M.; Falco, G.; Angrisano, T. Identification of Cdk8 and Cdkn2d as New Prame-Target Genes in 2C-like Embryonic Stem Cells. Genes 2022, 13, 1745. https://doi.org/10.3390/genes13101745
Lucci V, De Marino E, Tagliaferri D, Amente S, Pollice A, Calabrò V, Vivo M, Falco G, Angrisano T. Identification of Cdk8 and Cdkn2d as New Prame-Target Genes in 2C-like Embryonic Stem Cells. Genes. 2022; 13(10):1745. https://doi.org/10.3390/genes13101745
Chicago/Turabian StyleLucci, Valeria, Elena De Marino, Daniela Tagliaferri, Stefano Amente, Alessandra Pollice, Viola Calabrò, Maria Vivo, Geppino Falco, and Tiziana Angrisano. 2022. "Identification of Cdk8 and Cdkn2d as New Prame-Target Genes in 2C-like Embryonic Stem Cells" Genes 13, no. 10: 1745. https://doi.org/10.3390/genes13101745
APA StyleLucci, V., De Marino, E., Tagliaferri, D., Amente, S., Pollice, A., Calabrò, V., Vivo, M., Falco, G., & Angrisano, T. (2022). Identification of Cdk8 and Cdkn2d as New Prame-Target Genes in 2C-like Embryonic Stem Cells. Genes, 13(10), 1745. https://doi.org/10.3390/genes13101745