
Genetic Diversity and Population Genetic Structure of Aedes albopictus in the Yangtze River Basin, China
 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Sampling
2.2. Genotyping of Mosquito Samples
2.3. Data Analysis
3. Results
3.1. Genetic Diversity and Variation
3.2. Population Structure and Differentiation Based on Microsatellite Analysis
3.3. Haplotype Diversity and Network Analysis Based on CoxI Sequences
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Gratz, N.G. Critical review of the vector status of Aedes albopictus. Med. Vet. Entomol. 2004, 18, 215–227. [Google Scholar] [CrossRef]
- Bonizzoni, M.; Gasperi, G.; Chen, X.; James, A.A. The invasive mosquito species Aedes albopictus: Current knowledge and future perspectives. Trends Parasitol. 2013, 29, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Tatem, A.J.; Hay, S.I.; Rogers, D.J. Global traffic and disease vector dispersal. Proc. Natl. Acad. Sci. USA 2006, 103, 6242–6247. [Google Scholar] [CrossRef]
- Tatem, A.J.; Rogers, D.J.; Hay, S.I. Global transport networks and infectious disease spread. Adv. Parasitol. 2006, 62, 293–343. [Google Scholar]
- Wu, F.; Liu, Q.; Lu, L.; Wang, J.; Song, X.; Ren, D. Distribution of Aedes albopictus (Diptera: Culicidae) in northwestern China. Vector-Borne Zoonotic Dis. 2011, 11, 1181–1186. [Google Scholar] [CrossRef]
- Liu, K.; Hou, X.; Wang, Y.; Sun, J.; Xiao, J.; Li, R.; Lu, L.; Xu, L.; Sang, S.; Hu, J.; et al. The driver of dengue fever incidence in two high-risk areas of China: A comparative study. Sci. Rep. 2019, 9, 19510. [Google Scholar] [CrossRef] [PubMed]
- Reiter, P. Aedes albopictus and the world trade in used tires, 1988–1995: The shape of things to come? J. Am. Mosq. Control Assoc. 1998, 14, 83–94. [Google Scholar] [PubMed]
- Laird, M.; Calder, L.; Thornton, R.C.; Syme, R.; Holder, P.; Mogi, M. Japanese Aedes albopictus among four mosquito species reaching New Zealand in used tires. J. Am. Mosq. Control Assoc. 1994, 10, 14–23. [Google Scholar]
- Craven, R.B.; Eliason, D.A.; Francy, D.B.; Reiter, P.; Campos, E.G.; Jakob, W.L.; Smith, G.C.; Bozzi, C.J.; Moore, C.G.; Maupin, G.O. Importation of Aedes albopictus and other exotic mosquito species into the United States in used tires from Asia. J. Am. Mosq. Control Assoc. 1988, 4, 138–142. [Google Scholar] [PubMed]
- Moore, C.G.; Mitchell, C.J. Aedes albopictus in the United States: Ten-year presence and public health implications. Emerg. Infect. Dis. 1997, 3, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Reiter, P.; Sprenger, D. The used tire trade: A mechanism for the worldwide dispersal of container breeding mosquitoes. J. Am. Mosq. Control Assoc. 1987, 3, 494–501. [Google Scholar]
- Benedict, M.Q.; Levine, R.S.; Hawley, W.A.; Lounibos, L.P. Spread of the tiger: Global risk of invasion by the mosquito Aedes albopictus. Vector Borne Zoonotic Dis. 2007, 7, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Scholte, E.J.; Dijkstra, E.; Blok, H.; De Vries, A.; Takken, W.; Hofhuis, A.; Koopmans, M.; De Boer, A.; Reusken, C.B.E.M. Accidental importation of the mosquito Aedes albopictus into the Netherlands: A survey of mosquito distribution and the presence of dengue virus. Med. Vet. Entomol. 2008, 22, 352–358. [Google Scholar] [CrossRef]
- Bennett, K.; Martínez, C.G.; Almanza, A.; Rovira, J.R.; McMillan, W.O.; Enriquez, V.; Barraza, E.; Diaz, M.; Sanchez-Galan, J.; Whiteman, A.; et al. High infestation of invasive Aedes mosquitoes in used tires along the local transport network of Panama. Parasit. Vectors 2019, 12, 264. [Google Scholar] [CrossRef]
- Lwande, O.W.; Obanda, V.; Lindström, A.; Ahlm, C.; Evander, M.; Näslund, J.; Bucht, G. Globe-Trotting Aedes aegypti and Aedes albopictus: Risk Factors for Arbovirus Pandemics. Vector Borne Zoonotic Dis. 2020, 20, 71–81. [Google Scholar] [CrossRef]
- Sherpa, S.; Blum, M.G.B.; Capblancq, T.; Cumer, T.; Rioux, D.; Després, L. Unravelling the invasion history of the Asian tiger mosquito in Europe. Mol. Ecol. 2019, 28, 2360–2377. [Google Scholar] [CrossRef] [PubMed]
- Egizi, A.; Kiser, J.; Abadam, C.; Fonseca, D.M. The hitchhiker’s guide to becoming invasive: Exotic mosquitoes spread across a US state by human transport not autonomous flight. Mol. Ecol. 2016, 25, 3033–3047. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez-Justicia, A.; Koenraadt, C.J.M.; Stroo, A.; van Lammeren, R.; Takken, W. Risk-Based and Adaptive Invasive Mosquito Surveillance at Lucky Bamboo and Used Tire Importers in the Netherlands. J. Am. Mosq. Control Assoc. 2020, 36, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Guagliardo, S.A.; Morrison, A.C.; Barboza, J.L.; Requena, E.; Astete, H.; Vazquez-Prokopec, G.; Kitron, U. River boats contribute to the regional spread of the dengue vector Aedes aegypti in the Peruvian Amazon. PLoS Negl. Trop. Dis. 2015, 9, e0003648. [Google Scholar] [CrossRef]
- Miller, M.J.; Loaiza, J.R. Geographic expansion of the invasive mosquito Aedes albopictus across Panama—Implications for control of dengue and Chikungunya viruses. PLoS Negl. Trop. Dis. 2015, 9, e0003383. [Google Scholar] [CrossRef] [PubMed]
- Hofhuis, A.; Reimerink, J.; Reusken, C.B.E.M.; Scholte, E.J.; De Boer, A.; Takken, W.; Koopmans, M. The hidden passenger of lucky bamboo: Do imported Aedes albopictus mosquitoes cause dengue virus transmission in the Netherlands? Vector Borne Zoonotic Dis. 2009, 9, 217–220. [Google Scholar] [CrossRef]
- Díaz-Nieto, L.M.; Chiappero, M.B.; de Astarloa, C.D.; Maciá, A.; Gardenal, C.N.; Berón, C.M. Genetic Evidence of Expansion by Passive Transport of Aedes (Stegomyia) aegypti in Eastern Argentina. PLoS Negl. Trop. Dis. 2016, 10, e0004839. [Google Scholar] [CrossRef]
- Fonzi, E.; Higa, Y.; Bertuso, A.G.; Futami, K.; Minakawa, N. Human-Mediated Marine Dispersal Influences the Population Structure of Aedes aegypti in the Philippine Archipelago. PLoS Negl. Trop. Dis. 2015, 9, e0003829. [Google Scholar] [CrossRef] [PubMed]
- Guagliardo, S.A.J.; Lee, Y.; Pierce, A.A.; Wong, J.; Chu, Y.Y.; Morrison, A.C.; Astete, H.; Brosi, B.; Vazquez-Prokopec, G.; Scott, T.W.; et al. The genetic structure of Aedes aegypti populations is driven by boat traffic in the Peruvian Amazon. PLoS Negl. Trop. Dis. 2019, 13, e0007552. [Google Scholar] [CrossRef]
- Battaglia, V.; Gabrieli, P.; Brandini, S.; Capodiferro, M.R.; Javier, P.A.; Chen, X.-G.; Achilli, A.; Semino, O.; Gomulski, L.M.; Malacrida, A.R.; et al. The Worldwide Spread of the Tiger Mosquito as Revealed by Mitogenome Haplogroup Diversity. Front. Genet. 2016, 7, 208. [Google Scholar] [CrossRef]
- Goubert, C.; Minard, G.; Vieira, C.; Boulesteix, M. Population genetics of the Asian tiger mosquito Aedes albopictus, an invasive vector of human diseases. Heredity 2016, 117, 125–134. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, J.; Song, Z.; He, Y.; Zheng, Z.; Fan, P.; Yang, D.; Zhou, G.; Zhong, D.; Zheng, X. Patterns of spatial genetic structures in Aedes albopictus (Diptera: Culicidae) populations in China. Parasit. Vectors 2019, 12, 552. [Google Scholar] [CrossRef]
- Schmidt, T.L.; Swan, T.; Chung, J.; Karl, S.; Demok, S.; Yang, Q.; Field, M.A.; Muzari, M.O.; Ehlers, G.; Brugh, M.; et al. Spatial population genomics of a recent mosquito invasion. Mol. Ecol. 2021, 30, 1174–1189. [Google Scholar] [CrossRef]
- Maynard, A.J.; Ambrose, L.; Cooper, R.D.; Chow, W.K.; Davis, J.B.; Muzari, M.O.; van den Hurk, A.F.; Hall-Mendelin, S.; Hasty, J.M.; Burkot, T.R.; et al. Tiger on the prowl: Invasion history and spatio-temporal genetic structure of the Asian tiger mosquito Aedes albopictus (Skuse 1894) in the Indo-Pacific. PLoS Negl. Trop. Dis. 2017, 11, e0005546. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-D.; Gao, J.; Xing, D.; Guo, X.-X.; Dong, Y.-D.; Li, C.-X.; Zheng, Z.; Ma, Z.; Wu, Z.-M.; Zhu, X.-J.; et al. Fine-scale genetic structure and wolbachia infection of Aedes albopictus (Diptera: Culicidae) in Nanjing city, China. Front. Genet. 2022, 13, 827655. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhang, H.-D.; Guo, X.-X.; Xing, D.; Dong, Y.-D.; Lan, C.-J.; Wang, G.; Li, C.-J.; Li, C.-X.; Zhao, T.-Y. Dispersal patterns and population genetic structure of Aedes albopictus (Diptera: Culicidae) in three different climatic regions of China. Parasit. Vectors 2021, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Bonacum, J.; DeSalle, R.; O’Grady, P.; Olivera DS, C.G.; Wintermute, J.; Zilversmit, M. New nuclear and mitochondrial primers for systematics and comparative genomics in Drosophilidae. Drosoph. Inf. Serv. 2001, 84, 201–204. [Google Scholar]
- Goudet, J. FSTAT, a Program to Estimate and Test Gene Diversities and Fixation Indices (Version 2.9.3). Updated from Goudet (1995). 2001. Available online: http://www.unil.ch/izea/softwares/fstat.html (accessed on 5 July 2021).
- Rousset, F. genepop’007: A complete re-implementation of the genepop software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Slatkin, M. A measure of population subdivision based on microsatellite allele frequencies. Genetics 1995, 139, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Piry, S.; Luikart, G.; Cornuet, J.-M. Bottleneck: A Computer Program for Detecting Recent Reductions in the Effective Population Size Using Allele Frequency Data. 2017. Available online: http://www.ensam.inra.fr/URLB (accessed on 5 July 2021).
- Rozas, J.; Sánchez-DelBarrio, J.C.; Messeguer, X.; Rozas, R. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 2003, 19, 2496–2497. [Google Scholar] [CrossRef]
- Fröhlich, F.; Schmidt, S.L. Rational design of transcranial current stimulation (TCS) through mechanistic insights into cortical network dynamics. Front. Hum. Neurosci. 2013, 7, 804. [Google Scholar] [CrossRef]
- Múrias dos Santos, A.; Cabezas, M.P.; Tavares, A.I.; Xavier, R.; Branco, M. tcsBU: A tool to extend TCS network layout and visualization. Bioinformatics 2016, 32, 627–628. [Google Scholar] [CrossRef] [PubMed]
- Ditta, A.; Zhou, Z.; Cai, X.; Shehzad, M.; Wang, X.; Okubazghi, K.W.; Xu, Y.; Hou, Y.; Iqbal, M.S.; Khan, M.K.R.; et al. Genome-Wide Mining and Characterization of SSR Markers for Gene Mapping and Gene Diversity in Gossypium barbadense L. and Gossypium darwinii G. Watt Accessions. Agronomy 2018, 8, 181. [Google Scholar] [CrossRef]
- Nei, M.; Maruyama, T.; Chakraborty, R. The bottleneck effect and genetic variability in populations. Evolution 1975, 29, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Maudet, C.; Miller, C.; Bassano, B.; Breitenmoser-Würsten, C.; Gauthier, D.; Obexer-Ruff, G.; Michallet, J.; Taberlet, P.; Luikart, G. Microsatellite DNA and recent statistical methods in wildlife conservation management: Applications in Alpine ibex [Capra ibex (ibex)]. Mol. Ecol. 2002, 11, 421–436. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar]
- Takezaki, N.; Nei, M. Genetic distances and reconstruction of phylogenetic trees from microsatellite DNA. Genetics 1996, 144, 389–399. [Google Scholar] [CrossRef]
- Multini, L.C.; de Souza, A.L.D.S.; Marrelli, M.T.; Wilke, A.B.B. Population structuring of the invasive mosquito Aedes albopictus (Diptera: Culicidae) on a microgeographic scale. PLoS ONE 2019, 14, e0220773. [Google Scholar] [CrossRef]
- Sherpa, S.; Rioux, D.; Pougnet-Lagarde, C.; Després, L. Genetic diversity and distribution differ between long-established and recently introduced populations in the invasive mosquito Aedes albopictus. Infect. Genet. Evol. 2018, 58, 145–156. [Google Scholar] [CrossRef]
- Gippet, J.M.; Liebhold, A.M.; Fenn-Moltu, G.; Bertelsmeier, C. Human-mediated dispersal in insects. Curr. Opin. Insect Sci. 2019, 35, 96–102. [Google Scholar] [CrossRef]
- Schmidt, T.L.; Rašić, G.; Zhang, D.; Zheng, X.; Xi, Z.; Hoffmann, A.A. Genome-wide SNPs reveal the drivers of gene flow in an urban population of the Asian Tiger Mosquito, Aedes albopictus. PLoS Negl. Trop. Dis. 2017, 11, e0006009. [Google Scholar] [CrossRef]
- Medley, K.A.; Jenkins, D.G.; Hoffman, E.A. Human-aided and natural dispersal drive gene flow across the range of an invasive mosquito. Mol. Ecol. 2014, 24, 284–295. [Google Scholar] [CrossRef]
- Zé-Zé, L.; Borges, V.; Osório, H.C.; Machado, J.; Gomes, J.P.; Alves, M.J. Mitogenome diversity of Aedes (Stegomyia) albopictus: Detection of multiple introduction events in Portugal. PLoS Negl. Trop. Dis. 2020, 14, e0008657. [Google Scholar] [CrossRef]
- Latreille, A.C.; Milesi, P.; Magalon, H.; Mavingui, P.; Atyame, C.M. High genetic diversity but no geographical structure of Aedes albopictus populations in Reunion Island. Parasit. Vectors 2019, 12, 597. [Google Scholar] [CrossRef]
- Eritja, R.; Palmer, J.R.B.; Roiz, D.; Sanpera-Calbet, I.; Bartumeus, F. Direct Evidence of Adult Aedes albopictus Dispersal by Car. Sci. Rep. 2017, 7, 14399. [Google Scholar] [CrossRef] [PubMed]
- Villena, O.C.; Terry, I.; Iwata, K.; Landa, E.R.; LaDeau, S.L.; Leisnham, P.T. Effects of tire leachate on the invasive mosquito Aedes albopictus and the native congener Aedes triseriatus. PeerJ 2017, 5, e3756. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.R. Genetic Variation in Insect Vectors: Death of Typology? Insects 2018, 9, 139. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.R.; Evans, B.R. How Much Does Inbreeding Reduce Heterozygosity? Empirical Results from Aedes aegypti. Am. J. Trop. Med. Hyg. 2017, 96, 157–158. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
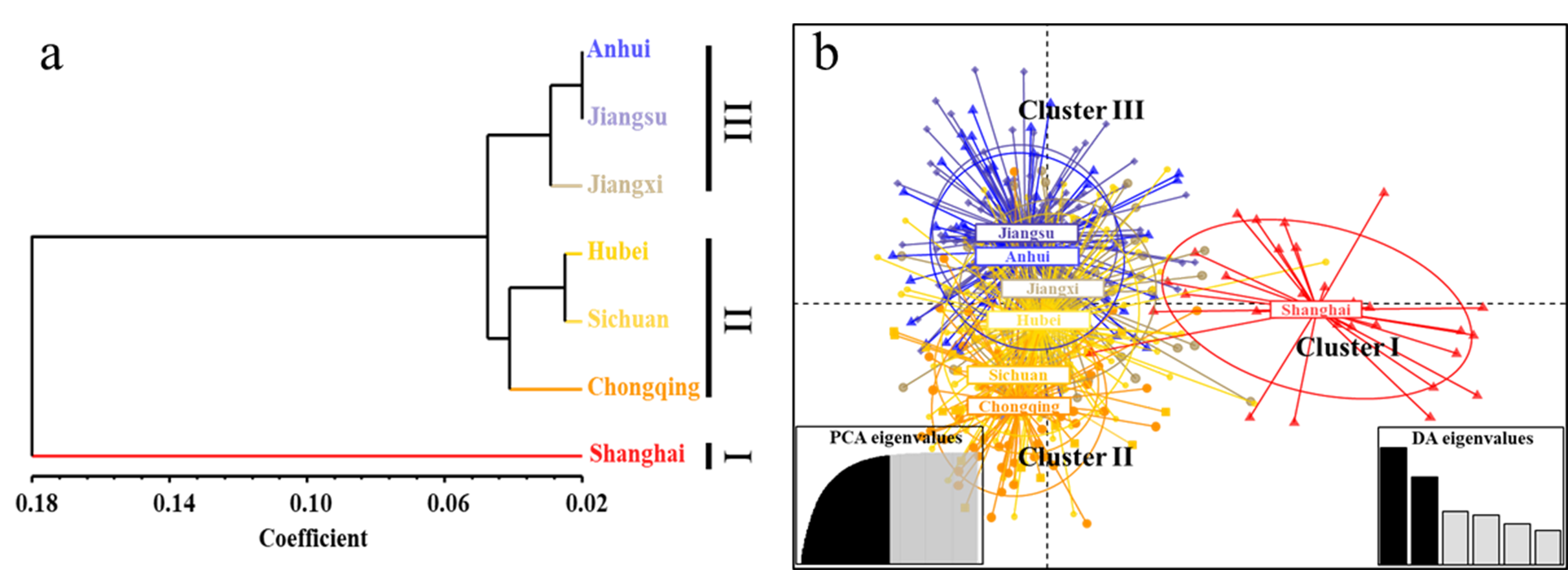{kind=link}
{kind=link}
| No. | Location | Population Code | Geographic Coordinate | Sample Size |
|---|---|---|---|---|
| 1 | Anhui (downstream) | AHBH | N30°53′10.41″ E117°46′18.71″ | 25 |
| 2 | AHYA | N30°57′31.85″ E117°47′14.68″ | 30 | |
| 3 | AHTL | N30°56′12.57″ E117°49′16.35″ | 30 | |
| 4 | Jiangsu (downstream) | NJDX | N32°03′25.02″ E118°46′27.42″ | 26 |
| 5 | JSJY | N31°54′05.29″ E120°16′20.34″ | 28 | |
| 6 | JYCS | N31°55′31.69″ E120°15′19.61″ | 30 | |
| 7 | NJZS | N32°05′19.69″ E118°43′54.56″ | 30 | |
| 8 | NJTH | N32°04′14.44″ E118°45′54.30″ | 30 | |
| 9 | Jiangxi (downstream) | JJLS | N29°40′07.78″ E115°56′54.98″ | 28 |
| 10 | JJXY | N29°40′45.15″ E116°00′17.82″ | 30 | |
| 11 | JJWF | N29°43′54.65″ E115°59′44.95″ | 30 | |
| 12 | Shanghai (downstream) (downstream) | SHJD | N31°12′03.49″ E121°25′41.03″ | 25 |
| 13 | Hubei (midstream) | WHWC | N30°33′18.14″ E114°17′35.34″ | 30 |
| 14 | WHDX | N30°31′52.51″ E114°21′12.32″ | 30 | |
| 15 | HBSX | N30°43′50.35″ E111°18′24.41″ | 26 | |
| 16 | HBQN | N30°43′13.14″ E111°16′16.61″ | 27 | |
| 17 | HBYC | N30°40′06.23″ E111°16′01.27″ | 30 | |
| 18 | HBBD | N30°19′33.84″ E112°14′28.09″ | 30 | |
| 19 | Chongqing (upstream) | CQDX | N29°34′01.11″ E106°27′51.77″ | 27 |
| 20 | CQWZ | N30°49′17.86″ E108°22′40.18″ | 30 | |
| 21 | Sichuan (upstream) | KZXZ | N30°39′57.73″ E104°03′00.46″ | 22 |
| 22 | SCDX | N30°37′57.88″ E104°04′52.49″ | 30 |
| Location | na | ne | I | Ho | He | WHD | FIS |
|---|---|---|---|---|---|---|---|
| Anhui | 11.455 ± 2.770 | 4.041 ± 2.208 | 1.610 ± 0.486 | 0.532 ± 0.228 | 0.688 ± 0.177 | 0.252 *** | 0.143 *** |
| Jiangsu | 11.273 ± 3.349 | 3.896 ± 1.841 | 1.578 ± 0.478 | 0.487 ± 0.229 | 0.678 ± 0.187 | 0.288 *** | 0.210 *** |
| Jiangxi | 9.818 ± 3.093 | 4.166 ± 2.227 | 1.576 ± 0.498 | 0.532 ± 0.229 | 0.692 ± 0.166 | 0.272 *** | 0.150 *** |
| Shanghai | 5.455 ± 2.115 | 3.106 ± 1.436 | 1.209 ± 0.469 | 0.545 ± 0.212 | 0.617 ± 0.187 | 0.123 | 0.047 |
| Hubei | 11.273 ± 3.319 | 4.223 ± 1.831 | 1.639 ± 0.456 | 0.513 ± 0.216 | 0.709 ± 0.161 | 0.329 *** | 0.212 *** |
| Chongqing | 8.455 ± 2.423 | 3.649 ± 1.634 | 1.460 ± 0.466 | 0.495 ± 0.244 | 0.659 ± 0.196 | 0.313 *** | 0.171 *** |
| Sichuan | 8.273 ± 1.849 | 3.910 ± 1.628 | 1.514 ± 0.430 | 0.522 ± 0.218 | 0.693 ± 0.176 | 0.342 ** | 0.168 *** |
| Anhui | Jiangsu | Jiangxi | Hubei | Chongqing | Sichuan | Shanghai | |
|---|---|---|---|---|---|---|---|
| Anhui | 0.155 ± 0.004 | 0.004 ± 0.001 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | |
| Jiangsu | 0.002 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | |
| Jiangxi | 0.008 | 0.013 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | |
| Hubei | 0.011 | 0.011 | 0.015 | 0.000 ± 0.000 | 0.002 ± 0.000 | 0.000 ± 0.000 | |
| Chongqing | 0.022 | 0.027 | 0.024 | 0.015 | 0.000 ± 0.000 | 0.000 ± 0.000 | |
| Sichuan | 0.018 | 0.021 | 0.019 | 0.009 | 0.015 | 0.000 ± 0.000 | |
| Shanghai | 0.073 | 0.075 | 0.067 | 0.067 | 0.108 | 0.082 |
| Yangtze River Basin | Anhui | Jiangsu | Jiangxi | Hubei | Chongqing | Sichuan | Shanghai | |
|---|---|---|---|---|---|---|---|---|
| S.M.M. | He < Heq * | 11 | 11 | 10 | 11 | 10 | 10 | 8 |
| He > Heq * | 0 | 0 | 1 | 0 | 1 | 1 | 3 | |
| p (He < Heq) | 0.000 | 0.000 | 0.001 | 0.000 | 0.001 | 0.001 | 0.032 | |
| T.P.M. | He < Heq | 9 | 9 | 6 | 8 | 7 | 6 | 4 |
| He > Heq | 2 | 2 | 5 | 3 | 4 | 5 | 7 | |
| p (He < Heq) | 0.007 | 0.007 | 0.265 | 0.031 | 0.111 | 0.265 | 0.514 | |
| NO. | Location | Region | H | Hd | π | k | Tajima’s D | Fu’s Fs | Haplotypes |
|---|---|---|---|---|---|---|---|---|---|
| 1 | downstream | Anhui | 9 | 0.315 | 0.001 | 0.341 | −1.976 * | −1.624 | H1(3),H2(67),H3(2),H4(2),H5(1),H6(1),H7(2),H8(2),H9(1) |
| 2 | downstream | Jiangsu | 6 | 0.179 | 0.0004 | 0.185 | −1.669 | −2.407 * | H2(105),H3(1),H15(1),H16(6),H23(2),H24(1) |
| 3 | downstream | Jiangxi | 7 | 0.369 | 0.0008 | 0.403 | −1.603 | −1.154 | H2(57),H9(4),H10(1),H15(2),H17(5),H20(2),H21(1), |
| 4 | downstream | Shanghai | 1 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | H2(25) |
| 5 | midstream | Hubei | 13 | 0.393 | 0.001 | 0.436 | −1.958 * | −2.362 * | H2(135),H4(2),H9(2),H10(4),H11(3),H12(1),H15(9),H16(1),H17(9),H18(5),H19(1),H21(1) |
| 6 | upstream | Chongqing | 4 | 0.074 | 0.0002 | 0.075 | −1.639 | −3.251 ** | H2(77),H12(1),H13(1),H14(1) |
| 7 | upstream | Sichuan | 7 | 0.315 | 0.0007 | 0.337 | −1.885 * | −3.229 * | H2(43),H3(1),H9(1),H15(1),H22(1),H25(4),H26(1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.-D.; Gao, J.; Li, C.-X.; Ma, Z.; Liu, Y.; Wang, G.; Liu, Q.; Xing, D.; Guo, X.-X.; Zhao, T.; et al. Genetic Diversity and Population Genetic Structure of Aedes albopictus in the Yangtze River Basin, China. Genes 2022, 13, 1950. https://doi.org/10.3390/genes13111950
Zhang H-D, Gao J, Li C-X, Ma Z, Liu Y, Wang G, Liu Q, Xing D, Guo X-X, Zhao T, et al. Genetic Diversity and Population Genetic Structure of Aedes albopictus in the Yangtze River Basin, China. Genes. 2022; 13(11):1950. https://doi.org/10.3390/genes13111950
Chicago/Turabian StyleZhang, Heng-Duan, Jian Gao, Chun-Xiao Li, Zu Ma, Yuan Liu, Ge Wang, Qing Liu, Dan Xing, Xiao-Xia Guo, Teng Zhao, and et al. 2022. "Genetic Diversity and Population Genetic Structure of Aedes albopictus in the Yangtze River Basin, China" Genes 13, no. 11: 1950. https://doi.org/10.3390/genes13111950
APA StyleZhang, H.-D., Gao, J., Li, C.-X., Ma, Z., Liu, Y., Wang, G., Liu, Q., Xing, D., Guo, X.-X., Zhao, T., Jiang, Y.-T., Dong, Y.-D., & Zhao, T.-Y. (2022). Genetic Diversity and Population Genetic Structure of Aedes albopictus in the Yangtze River Basin, China. Genes, 13(11), 1950. https://doi.org/10.3390/genes13111950