
Sheep Post-Domestication Expansion in the Context of Mitochondrial and Y Chromosome Haplogroups and Haplotypes



Abstract
:1. Introduction
2. Mitochondrial Haplogroups and Haplotypes
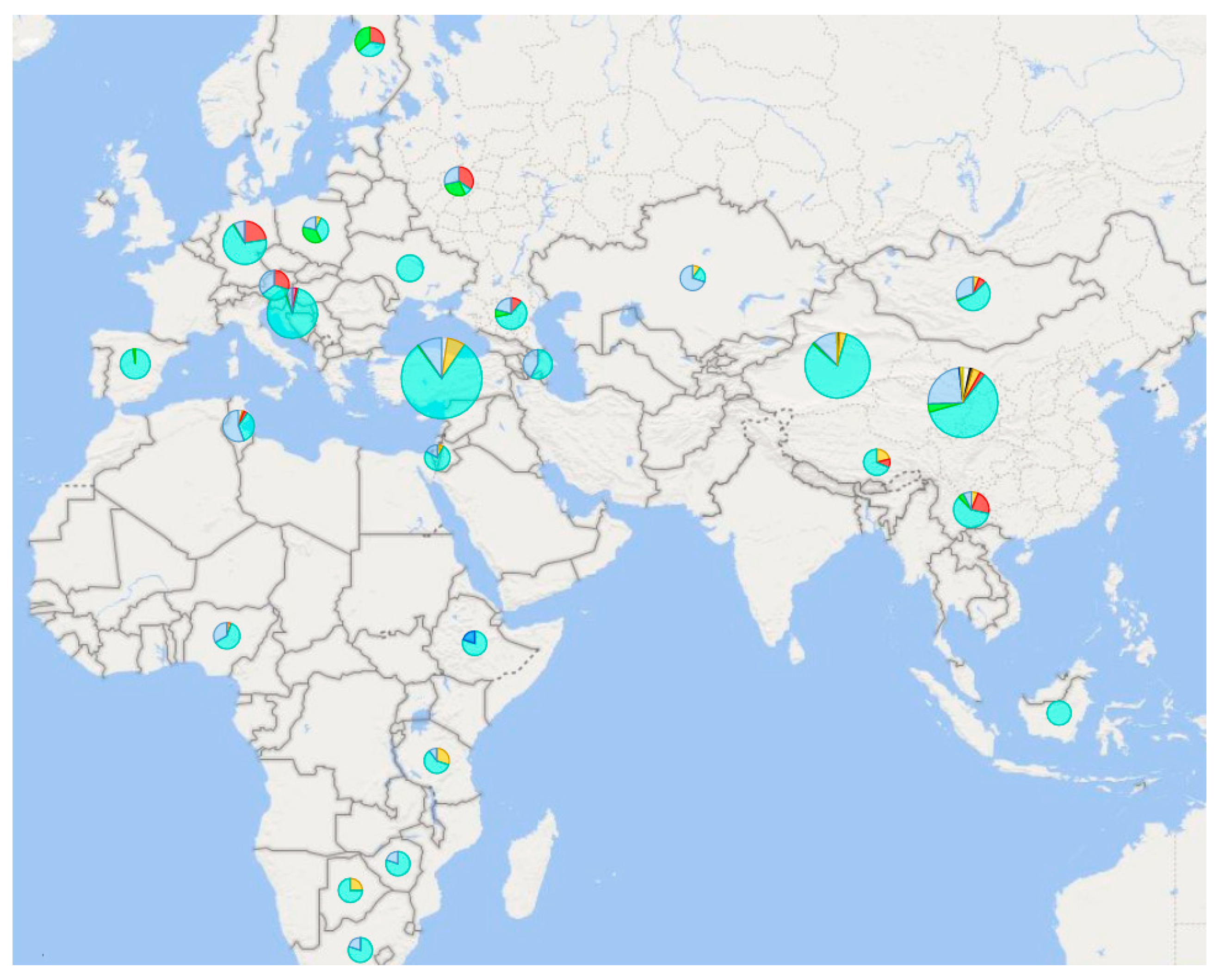
2.1. Europe
2.2. Asia
- Lineage A spread first to the Mongolian Plateau and the Indian subcontinent. Later, it expanded from the Mongolian Plateau to northern and southwestern China. According to [77], lineage A was the most abundant lineage in ancient Bronze Age China (95.5%). Its abundance increased from west to east.
- Lineage B headed first on the Mongolian Plateau and colonized northern and southwestern China and India from there.
- Lineage C also first colonized the Mongolian Plateau. From there it headed to northern China and then to the Indian subcontinent.
2.3. Africa
2.4. America and Australia
3. Haplogroups and Haplotypes of Male Y Chromosomes
4. Historical Background
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zeder, M.A. Out of the Fertile Crescent: The Dispersal of Domestic Livestock through Europe and Africa. In Human Dispersal and Species Movement; Boivin, N., Petraglia, M., Crassard, R., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 261–303. [Google Scholar]
- Zeder, M.A. Animal Domestication in the Zagros: A Review of Past and Current Research. Paléorient 1999, 25, 11–25. [Google Scholar] [CrossRef]
- Baird, D.; Fairbairn, A.; Jenkins, E.; Martin, L.; Middleton, C.; Pearson, J.; Asouti, E.; Edwards, Y.; Kabukcu, C.; Mustafaoğlu, G.; et al. Agricultural Origins on the Anatolian Plateau. Proc. Natl. Acad. Sci. USA 2018, 115, E3077–E3086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luigi-Sierra, M.G.; Mármol-Sánchez, E.; Amills, M. Comparing the Diversity of the Casein Genes in the Asian Mouflon and Domestic Sheep. Anim. Genet. 2020, 51, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Wright, D. Article Commentary: The Genetic Architecture of Domestication in Animals. Bioinform. Biol. Insights 2015, 9 (Suppl. S4), 11–20. [Google Scholar] [CrossRef] [Green Version]
- Lv, F.-H.; Agha, S.; Kantanen, J.; Colli, L.; Stucki, S.; Kijas, J.W.; Joost, S.; Li, M.-H.; Ajmone Marsan, P. Adaptations to Climate-Mediated Selective Pressures in Sheep. Mol. Biol. Evol. 2014, 31, 3324–3343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salces-Ortiz, J.; González, C.; Martínez, M.; Mayoral, T.; Calvo, J.H.; Serrano, M. Looking for Adaptive Footprints in the HSP90AA1 Ovine Gene. BMC Evol. Biol. 2015, 15, 7. [Google Scholar] [CrossRef] [Green Version]
- Kijas, J.W.; Lenstra, J.A.; Hayes, B.; Boitard, S.; Porto Neto, L.R.; San Cristobal, M.; Servin, B.; McCulloch, R.; Whan, V.; Gietzen, K.; et al. Genome-Wide Analysis of the World’s Sheep Breeds Reveals High Levels of Historic Mixture and Strong Recent Selection. PLoS Biol. 2012, 10, e1001258. [Google Scholar] [CrossRef] [Green Version]
- Ryder, M.L. Sheep. In Evolution of Domesticated Animals; Manson, I.L., Ed.; Longman Group Limited: London, UK; New York, NY, USA, 1984; pp. 63–84. [Google Scholar]
- Hiendleder, S.; Kaupe, B.; Wassmuth, R.; Janke, A. Molecular Analysis of Wild and Domestic Sheep Questions Current Nomenclature and Provides Evidence for Domestication from Two Different Subspecies. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2002, 269, 893–904. [Google Scholar] [CrossRef] [Green Version]
- Tapio, M.; Marzanov, N.; Ozerov, M.; Ćinkulov, M.; Gonzarenko, G.; Kiselyova, T.; Murawski, M.; Viinalass, H.; Kantanen, J. Sheep Mitochondrial DNA Variation in European, Caucasian, and Central Asian Areas. Mol. Biol. Evol. 2006, 23, 1776–1783. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, M.; Valentini, A.; Marsan, P.A.; Pariset, L. Mitochondrial DNA of Seven Italian Sheep Breeds Shows Faint Signatures of Domestication and Suggests Recent Breed Formation. Mitochondrial DNA 2013, 24, 577–583. [Google Scholar] [CrossRef]
- Rezaei, H.R.; Naderi, S.; Chintauan-Marquier, I.C.; Taberlet, P.; Virk, A.T.; Naghash, H.R.; Rioux, D.; Kaboli, M.; Pompanon, F. Evolution and Taxonomy of the Wild Species of the Genus Ovis (Mammalia, Artiodactyla, Bovidae). Mol. Phylogenet. Evol. 2010, 54, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Chessa, B.; Pereira, F.; Arnaud, F.; Amorim, A.; Goyache, F.; Mainland, I.; Kao, R.R.; Pemberton, J.M.; Beraldi, D.; Stear, M.J.; et al. Revealing the History of Sheep Domestication Using Retrovirus Integrations. Science 2009, 324, 532–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Chen, L.; Wang, W. Genomic Insights into Ruminant Evolution: From Past to Future Prospects. Zool. Res. 2019, 40, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Kijas, J.W.; Townley, D.; Dalrymple, B.P.; Heaton, M.P.; Maddox, J.F.; McGrath, A.; Wilson, P.; Ingersoll, R.G.; McCulloch, R.; McWilliam, S.; et al. A Genome Wide Survey of SNP Variation Reveals the Genetic Structure of Sheep Breeds. PLoS ONE 2009, 4, e4668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rannamäe, E.; Lõugas, L.; Niemi, M.; Kantanen, J.; Maldre, L.; Kadõrova, N.; Saarma, U. Maternal and Paternal Genetic Diversity of Ancient Sheep in Estonia from the Late Bronze Age to the Post-Medieval Period and Comparison with Other Regions in Eurasia. Anim. Genet. 2016, 47, 208–218. [Google Scholar] [CrossRef]
- Zhao, Y.-X.; Yang, J.; Lv, F.-H.; Hu, X.-J.; Xie, X.-L.; Zhang, M.; Li, W.-R.; Liu, M.-J.; Wang, Y.-T.; Li, J.-Q.; et al. Genomic Reconstruction of the History of Native Sheep Reveals the Peopling Patterns of Nomads and the Expansion of Early Pastoralism in East Asia. Mol. Biol. Evol. 2017, 34, 2380–2395. [Google Scholar] [CrossRef] [PubMed]
- Pardeshi, V.C.; Kadoo, N.Y.; Sainani, M.N.; Meadows, J.R.S.; Kijas, J.W.; Gupta, V.S. Mitochondrial Haplotypes Reveal a Strong Genetic Structure for Three Indian Sheep Breeds. Anim. Genet. 2007, 38, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Nigussie, H.; Mwacharo, J.M.; Osama, S.; Agaba, M.; Mekasha, Y.; Kebede, K.; Abegaz, S.; Pal, S.K. Genetic Diversity and Matrilineal Genetic Origin of Fat-Rumped Sheep in Ethiopia. Trop. Anim. Health Prod. 2019, 51, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-H.; Zhang, M.; Lv, F.-H.; Ren, X.; Li, W.-R.; Liu, M.-J.; Nam, K.; Bruford, M.W.; Li, M.-H. Contrasting Patterns of Genomic Diversity Reveal Accelerated Genetic Drift but Reduced Directional Selection on X-Chromosome in Wild and Domestic Sheep Species. Genome Biol. Evol. 2018, 10, 1282–1297. [Google Scholar] [CrossRef] [Green Version]
- Chessa, S.; Giambra, I.J.; Brandt, H.; Caroli, A.M.; Gootwine, E.; Erhardt, G. Genetic Diversity within Economically Important Loci in European, Middle Eastern, and African Sheep Breeds: An Insight into Their Development. Small Rumin. Res. 2017, 155, 72–80. [Google Scholar] [CrossRef]
- Brown, W.M.; George, M.; Wilson, A.C. Rapid Evolution of Animal Mitochondrial DNA. Proc. Natl. Acad. Sci. USA 1979, 76, 1967–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafia, P.; Tarang, A. Sequence Variations of Mitochondrial DNA Displacement-Loop in Iranian Indig…: Elektronické Publikace Dostupné Na ČZU. Iran. J. Appl. Anim. Sci. 2016, 6, 363–368. [Google Scholar]
- Mukhametzharova, I.; Islamov, Y.; Shauyenov, S.; Ibrayev, D.; Atavliyeva, S.; Tarlykov, P. Genetic Characterization of Kazakh Native Sheep Breeds Using Mitochondrial DNA. Online J. Biol. Sci. 2018, 18, 341–348. [Google Scholar] [CrossRef]
- Meadows, J.R.S.; Cemal, I.; Karaca, O.; Gootwine, E.; Kijas, J.W. Five Ovine Mitochondrial Lineages Identified from Sheep Breeds of the near East. Genetics 2007, 175, 1371–1379. [Google Scholar] [CrossRef] [Green Version]
- Yağci, S.; Baş, S.; Kiraz, S. Study of Mitochondrial DNA (MtDNA) D-Loop Region Polymorphism in Şavak Akkaraman Sheep. Turk. J. Vet. Anim. Sci. 2020, 44, 323–330. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information NCBI. Available online: https://www.ncbi.nlm.nih.gov/ipg/ADI57605.1 (accessed on 26 June 2020).
- Demirci, S.; Koban Baştanlar, E.; Dağtaş, N.D.; Pişkin, E.; Engin, A.; Özer, F.; Yüncü, E.; Doğan, Ş.A.; Togan, İ. Mitochondrial DNA Diversity of Modern, Ancient and Wild Sheep (Ovis gmelinii anatolica) from Turkey: New Insights on the Evolutionary History of Sheep. PLoS ONE 2013, 8, e81952. [Google Scholar] [CrossRef] [Green Version]
- Dymova, M.A.; Zadorozhny, A.V.; Mishukova, O.V.; Khrapov, E.A.; Druzhkova, A.S.; Trifonov, V.A.; Kichigin, I.G.; Tishkin, A.A.; Grushin, S.P.; Filipenko, M.L. Mitochondrial DNA Analysis of Ancient Sheep from Altai. Anim. Genet. 2017, 48, 615–618. [Google Scholar] [CrossRef]
- Wood, N.J.; Phua, S.H. Variation in the Control Region Sequence of the Sheep Mitochondrial Genome. Anim. Genet. 2009, 27, 25–33. [Google Scholar] [CrossRef]
- Sanna, D.; Barbato, M.; Hadjisterkotis, E.; Cossu, P.; Decandia, L.; Trova, S.; Pirastru, M.; Leoni, G.G.; Naitana, S.; Francalacci, P.; et al. The First Mitogenome of the Cyprus Mouflon (Ovis Gmelini Ophion): New Insights into the Phylogeny of the Genus Ovis. PLoS ONE 2015, 10, e0144257. [Google Scholar] [CrossRef] [Green Version]
- Meadows, J.R.S.; Hiendleder, S.; Kijas, J.W. Haplogroup Relationships between Domestic and Wild Sheep Resolved Using a Mitogenome Panel. Heredity 2011, 106, 700–706. [Google Scholar] [CrossRef] [Green Version]
- Lv, F.-H.; Peng, W.-F.; Yang, J.; Zhao, Y.-X.; Li, W.-R.; Liu, M.-J.; Ma, Y.-H.; Zhao, Q.-J.; Yang, G.-L.; Wang, F.; et al. Mitogenomic Meta-Analysis Identifies Two Phases of Migration in the History of Eastern Eurasian Sheep. Mol. Biol. Evol. 2015, 32, 2515–2533. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Xie, X.-L.; Wang, D.-F.; Zhao, C.; Lv, F.-H.; Li, X.; Yang, J.; Yu, J.-L.; Shen, M.; Gao, L.; et al. Paternal Origins and Migratory Episodes of Domestic Sheep. Curr. Biol. 2020, 30, 4085–4095.e6. [Google Scholar] [CrossRef]
- Çakırlar, C. The Evolution of Animal Husbandry in Neolithic Central-West Anatolia: The Zooarchaeological Record from Ulucak Höyük (c. 7040–5660 Cal. BC, Izmir, Turkey). Anatol. Stud. 2012, 62, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Mereu, P.; Pirastru, M.; Barbato, M.; Satta, V.; Hadjisterkotis, E.; Manca, L.; Naitana, S.; Leoni, G.G. Identification of an Ancestral Haplotype in the Mitochondrial Phylogeny of the Ovine Haplogroup B. PeerJ 2019, 7, e7895. [Google Scholar] [CrossRef]
- Dotsev, A.V.; Kunz, E.; Shakhin, A.V.; Petrov, S.N.; Kostyunina, O.V.; Okhlopkov, I.M.; Deniskova, T.E.; Barbato, M.; Bagirov, V.A.; Medvedev, D.G.; et al. The First Complete Mitochondrial Genomes of Snow Sheep (Ovis Nivicola) and Thinhorn Sheep (Ovis Dalli ) and Their Phylogenetic Implications for the Genus Ovis. Mitochondrial DNA Part B 2019, 4, 1332–1333. [Google Scholar] [CrossRef]
- Scheu, A.; Geörg, C.; Schulz, A.; Burger, J.; Benecke, N. The Arrival of Domesticated Animals in South-Eastern Europe as Seen from Ancient DNA. In Population Dynamics in Prehistory and Early History; Kaiser, E., Burger, J., Schier, W., Eds.; Walter de Gruyter: Berlin, Germany, 2012; pp. 45–54. [Google Scholar] [CrossRef] [Green Version]
- Molaee, V.; Bazzucchi, M.; De Mia, G.M.; Otarod, V.; Abdollahi, D.; Rosati, S.; Lühken, G. Phylogenetic Analysis of Small Ruminant Lentiviruses in Germany and Iran Suggests Their Expansion with Domestic Sheep. Sci. Rep. 2020, 10, 2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivieri, C.; Ermini, L.; Rizzi, E.; Corti, G.; Luciani, S.; Marota, I.; De Bellis, G.; Rollo, F. Phylogenetic Position of a Copper Age Sheep (Ovis aries) Mitochondrial DNA. PLoS ONE 2012, 7, e33792. [Google Scholar] [CrossRef] [Green Version]
- Niemi, M.; Bläuer, A.; Iso-Touru, T.; Nyström, V.; Harjula, J.; Taavitsainen, J.-P.; Storå, J.; Lidén, K.; Kantanen, J. Mitochondrial DNA and Y-Chromosomal Diversity in Ancient Populations of Domestic Sheep (Ovis aries) in Finland: Comparison with Contemporary Sheep Breeds. Genet. Sel. Evol. 2013, 45, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gáspárdy, A.; Berger, B.; Zabavnik-Piano, J.; Kovács, E.; Annus, K.; Zenke, P.; Sáfár, L.; Maróti-Agóts, Á. Comparison of MtDNA Control Region among Descendant Breeds of the Extinct Zaupel Sheep Revealed Haplogroup C and D in Central Europe. Vet. Med. Sci. 2021, 7, 2330–2338. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ding, X.; Zeng, Y.; Yue, Y.; Guo, X.; Guo, T.; Chu, M.; Wang, F.; Han, J.; Feng, R.; et al. Genetic Diversity and Phylogenetic Evolution of Tibetan Sheep Based on MtDNA D-Loop Sequences. PLoS ONE 2016, 11, e0159308. [Google Scholar] [CrossRef]
- Tapio, I.; Värv, S.; Bennewitz, J.; Maleviciute, J.; Fimland, E.; Grislis, Z.; Meuwissen, T.H.E.; Miceikiene, I.; Olsaker, I.; Viinalass, H.; et al. Prioritization for Conservation of Northern European Cattle Breeds Based on Analysis of Microsatellite Data. Conserv. Biol. 2006, 20, 1768–1779. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Kumar, S., Jr.; Kolte, A.P.; Kumar, S. Extensive Variation and Sub-Structuring in Lineage A MtDNA in Indian Sheep: Genetic Evidence for Domestication of Sheep in India. PLoS ONE 2013, 8, e77858. [Google Scholar] [CrossRef] [PubMed]
- Muigai, A.W.T.; Hanotte, O. The Origin of African Sheep: Archaeological and Genetic Perspectives. Afr. Archaeol. Rev. 2013, 30, 39–50. [Google Scholar] [CrossRef]
- Gornas, N.; Weimann, C.; El Hussien, A.; Erhardt, G. Genetic Characterization of Local Sudanese Sheep Breeds Using DNA Markers. Small Rumin. Res. 2011, 95, 27–33. [Google Scholar] [CrossRef]
- Resende, A.; Gonçalves, J.; Muigai, A.W.T.; Pereira, F. Mitochondrial DNA Variation of Domestic Sheep (Ovis aries) in Kenya. Anim. Genet. 2016, 47, 377–381. [Google Scholar] [CrossRef]
- Álvarez, I.; Capote, J.; Traoré, A.; Fonseca, N.; Pérez, K.; Cuervo, M.; Fernández, I.; Goyache, F. Mitochondrial Analysis Sheds Light on the Origin of Hair Sheep. Anim. Genet. 2013, 44, 344–347. [Google Scholar] [CrossRef]
- Kim, Y.S.; Tseveen, K.; Batsukh, B.; Seong, J.; Kong, H.S. Origin-Related Study of Genetic Diversity and Heteroplasmy of Mongolian Sheep (Ovis arie) Using Mitochondrial DNA. J. Anim. Reprod. Biotechnol. 2020, 35, 198–206. [Google Scholar] [CrossRef]
- Ganbold, O.; Lee, S.-H.; Seo, D.; Paek, W.K.; Manjula, P.; Munkhbayar, M.; Lee, J.H. Genetic Diversity and the Origin of Mongolian Native Sheep. Livest. Sci. 2019, 220, 17–25. [Google Scholar] [CrossRef]
- Xiao, P.; Niu, L.L.; Zhao, Q.J.; Chen, X.Y.; Wang, L.J.; Li, L.; Zhang, H.P.; Guo, J.Z.; Xu, H.Y.; Zhong, T. New Insights into Mitogenomic Phylogeny and Copy Number in Eight Indigenous Sheep Populations Based on the ATP Synthase and Cytochrome c Oxidase Genes. Animal 2018, 12, 1341–1349. [Google Scholar] [CrossRef]
- Ibrahim, A.; Budisatria, I.G.S.; Widayanti, R.; Artama, W.T. The Genetic Profiles and Maternal Origin of Local Sheep Breeds on Java Island (Indonesia) Based on Complete Mitochondrial DNA D-Loop Sequences. Vet. World 2020, 13, 2625–2634. [Google Scholar] [CrossRef]
- Arora, R.; Yadav, H.S.; Mishra, B.P. Mitochondrial DNA Diversity in Indian Sheep. Livest. Sci. 2013, 153, 50–55. [Google Scholar] [CrossRef]
- Ćinkulov, M.; Popovski, Z.; Porcu, K.; Tanaskovska, B.; Hodžić, A.; Bytyqi, H.; Mehmeti, H.; Margeta, V.; Djedović, R.; Hoda, A.; et al. Genetic Diversity and Structure of the West Balkan Pramenka Sheep Types as Revealed by Microsatellite and Mitochondrial DNA Analysis. J. Anim. Breed. Genet. 2008, 125, 417–426. [Google Scholar] [CrossRef]
- Dudu, A.; Ghiţă, E.; Costache, M.; Georgescu, S.E. Origin and Genetic Diversity of Romanian Racka Sheep Using Mitochondrial Markers. Small Rumin. Res. 2016, 144, 276–282. [Google Scholar] [CrossRef]
- Ferencakovic, M.; Curik, I.; Pérez-Pardal, L.; Royo, L.J.; Cubric-Curik, V.; Fernández, I.; Álvarez, I.; Kostelic, A.; Sprem, N.; Krapinec, K.; et al. Mitochondrial DNA and Y-Chromosome Diversity in East Adriatic Sheep. Anim. Genet. 2013, 44, 184–192. [Google Scholar] [CrossRef]
- Ghernouti, N.; Bodinier, M.; Ranebi, D.; Maftah, A.; Petit, D.; Gaouar, S.B.S. Control Region of MtDNA Identifies Three Migration Events of Sheep Breeds in Algeria. Small Rumin. Res. 2017, 155, 66–71. [Google Scholar] [CrossRef]
- Gorkhali, N.A.; Han, J.L.; Ma, Y.H. Mitochondrial DNA Variation in Indigenous Sheep (Ovis aries) Breeds of Nepal. Trop. Agric. Res. 2015, 26, 632. [Google Scholar] [CrossRef] [Green Version]
- Guangxin, E.; Yong-Ju, Z.; Ri-Su, N.; Yue-Hui, M.; Jia-Hua, Z.; Li-Peng, C.; Xiao-Yu, Q.; Zhong-Quan, Z.; Ya-Wang, S.; Xin, W.; et al. Meta-Analysis Evidence of Maternal Lineages in Chinese Tibetan Sheep Using MtDNA D-Loop Panel. Mitochondrial DNA Part A 2017, 28, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Du, L.-X.; Ma, Y.-H.; Guan, W.-J.; Li, H.-B.; Zhao, Q.-J.; Li, X.; Rao, S.-Q. A Novel Maternal Lineage Revealed in Sheep (Ovis aries). Anim. Genet. 2005, 36, 331–336. [Google Scholar] [CrossRef]
- Hamadalahmad, A.; Almeziad, M.; Javadmanesh, A. Genetic Similarity Comparison between Some Iranian and Middle Eastern Sheep Breeds Using Mitochondrial Control Region Sequencing. DYSONA Life Sci. 2020, 1, 20–24. [Google Scholar] [CrossRef]
- Hussain, T.; Babar, M.E.; Wajid, A. Extra Nuclear DNA Control Region and Cytochome b Gene Based Phylogeny Kashmir: Implications Towards Conservation. JAPS J. Anim. Plant Sci. 2016, 26, 1890–1893. [Google Scholar]
- Kandoussi, A.; Boujenane, I.; Auger, C.; Serranito, B.; Germot, A.; Piro, M.; Maftah, A.; Badaoui, B.; Petit, D. The Origin of Sheep Settlement in Western Mediterranean. Sci. Rep. 2020, 10, 10225. [Google Scholar] [CrossRef]
- Kirikci, K.; Noce, A.; Cam, M.A.; Mercan, L.; Amills, M. The Analysis of Mitochondrial Data Indicates the Existence of Population Substructure in Karayaka Sheep. Small Rumin. Res. 2018, 162, 25–29. [Google Scholar] [CrossRef]
- Koseniuk, A.; Słota, E. Mitochondrial Control Region Diversity in Polish Sheep Breeds. Arch. Anim. Breed. 2016, 59, 227–233. [Google Scholar] [CrossRef]
- Liu, J.; Lu, Z.; Yuan, C.; Wang, F.; Yang, B. Phylogeography and Phylogenetic Evolution in Tibetan Sheep Based on MT-CYB Sequences. Animals 2020, 10, 1177. [Google Scholar] [CrossRef] [PubMed]
- Meadows, J.R.S.; Li, K.; Kantanen, J.; Tapio, M.; Sipos, W.; Pardeshi, V.; Gupta, V.; Calvo, J.H.; Whan, V.; Norris, B.; et al. Mitochondrial Sequence Reveals High Levels of Gene Flow Between Breeds of Domestic Sheep from Asia and Europe. J. Hered. 2005, 96, 494–501. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, S.I.; Schwarzacher, T.; Heslop-Harrison, J.S. Complete Mitogenomes from Kurdistani Sheep: Abundant Centromeric Nuclear Copies Representing Diverse Ancestors. Mitochondrial DNA Part A 2018, 29, 1180–1193. [Google Scholar] [CrossRef] [Green Version]
- Oner, Y.; Calvo, J.H.; Elmaci, C. Investigation of the Genetic Diversity among Native Turkish Sheep Breeds Using MtDNA Polymorphisms. Trop. Anim. Health Prod. 2013, 45, 947–951. [Google Scholar] [CrossRef]
- Othman, O.E.; Pariset, L.; Balabel, E.A.; Marioti, M. Genetic Characterization of Egyptian and Italian Sheep Breeds Using Mitochondrial DNA. J. Genet. Eng. Biotechnol. 2015, 13, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Pariset, L.; Mariotti, M.; Gargani, M.; Joost, S.; Negrini, R.; Perez, T.; Bruford, M.; Ajmone Marsan, P.; Valentini, A. Genetic Diversity of Sheep Breeds from Albania, Greece, and Italy Assessed by Mitochondrial DNA and Nuclear Polymorphisms (SNPs). Sci. World J. 2011, 11, 1641–1659. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Ahlawat, S.; Sharma, H.; Sharma, P.; Panchal, P.; Arora, R.; Tantia, M.S. Microsatellite and Mitochondrial DNA Analyses Unveil the Genetic Structure of Native Sheep Breeds from Three Major Agro-Ecological Regions of India. Sci. Rep. 2020, 10, 20422. [Google Scholar] [CrossRef]
- Sulaiman, Y.; Wu, C.; Zhao, C. Phylogeny of 19 Indigenous Sheep Populations in Northwestern China Inferred from Mitochondrial DNA Control Region. Asian J. Anim. Vet. Adv. 2011, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Yüncü, E.; Demirci, S.; Koban Baştanlar, E.; Doğan, Ş.A.; Taşdemir, U.; Togan, İ. Comparative Study of Three Simple Molecular Approaches in Search of MtDNA Haplogroup Identification of Domestic Sheep. Small Rumin. Res. 2013, 114, 64–71. [Google Scholar] [CrossRef]
- Cai, D.; Tang, Z.; Yu, H.; Han, L.; Ren, X.; Zhao, X.; Zhu, H.; Zhou, H. Early History of Chinese Domestic Sheep Indicated by Ancient DNA Analysis of Bronze Age Individuals. J. Archaeol. Sci. 2011, 38, 896–902. [Google Scholar] [CrossRef]
- Chen, S.-Y.; Duan, Z.-Y.; Sha, T.; Xiangyu, J.; Wu, S.-F.; Zhang, Y.-P. Origin, Genetic Diversity, and Population Structure of Chinese Domestic Sheep. Gene 2006, 376, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Udo, H.M.J.; Budisatria, I.G.S. Fat-Tailed Sheep in Indonesia; an Essential Resource for Smallholders. Trop. Anim. Health Prod. 2011, 43, 1411–1418. [Google Scholar] [CrossRef] [Green Version]
- Horsburgh, K.A.; Rhines, A. Genetic Characterization of an Archaeological Sheep Assemblage from South Africa’s Western Cape. J. Archaeol. Sci. 2010, 37, 2906–2910. [Google Scholar] [CrossRef]
- Alonso, R.A.; Ulloa-Arvizu, R.; Gayosso-Vázquez, A. Mitochondrial DNA Sequence Analysis of the Mexican Creole Sheep (Ovis aries) Reveals a Narrow Iberian Maternal Origin. Mitochondrial DNA Part A 2017, 28, 793–800. [Google Scholar] [CrossRef]
- Delgado, J.V.; Perezgrovas, R.; Camacho, M.E.; Fresno, M.; Barba, C. The Wool-Less Canary Sheep and Their Relationship with the Present Breeds in America. Anim. Genet. Resour. Inf. 2000, 28, 27–34. [Google Scholar] [CrossRef]
- Spangler, G.L.; Rosen, B.D.; Ilori, M.B.; Hanotte, O.; Kim, E.-S.; Sonstegard, T.S.; Burke, J.M.; Morgan, J.L.M.; Notter, D.R.; Van Tassell, C.P. Whole Genome Structural Analysis of Caribbean Hair Sheep Reveals Quantitative Link to West African Ancestry. PLoS ONE 2017, 12, e0179021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muigai, A.W.T.; Hirbo, J.; Sharkey, S.; Rege, J.E.O.; Blackburn, H.; Hanotte, O. Genetic diversity and relationships of hair sheep breeds of the Americas: First results. In Proceedings of the 7th World Congress on Genetics Applied to Livestock Production, Montpellier, France, 19–23 August 2002; pp. 1–4. [Google Scholar]
- Campos, E.; Cuéllar, J.; Salvador, O.; García-Trejo, E.A.; Pereira, F. The Genetic Diversity and Phylogeography of Mexican Domestic Sheep. Small Rumin. Res. 2020, 187, 106109. [Google Scholar] [CrossRef]
- Parsons, D.; Nicholson, C.F. Assessing Policy Options for Agricultural Livestock Development: A Case Study of Mexico’s Sheep Sector. Cogent Food Agric. 2017, 3, 1313360. [Google Scholar] [CrossRef]
- Macias-Cruz, U.; Álvarez-Valenzuela, F.D.; Correa-Calderón, A.; Molina-Ramírez, L.; González-Reyna, A.; Soto-Navarro, S.; Avendaño-Reyes, L. Pelibuey Ewe Productivity and Subsequent Pre-Weaning Lamb Performance Using Hair-Sheep Breeds Under a Confinement System. J. Appl. Anim. Res. 2009, 36, 255–260. [Google Scholar] [CrossRef]
- Revelo, H.A.; López-Alvarez, D.; Landi, V.; Rizzo, L.; Alvarez, L.A. Mitochondrial DNA Variations in Colombian Creole Sheep Confirm an Iberian Origin and Shed Light on the Dynamics of Introduction Events of African Genotypes. Animals 2020, 10, 1594. [Google Scholar] [CrossRef] [PubMed]
- Bravo, S.; Larama, G.; Quiñones, J.; Paz, E.; Rodero, E.; Sepúlveda, N. Genetic Diversity and Phylogenetic Relationship among Araucana Creole Sheep and Spanish Sheep Breeds. Small Rumin. Res. 2019, 172, 23–30. [Google Scholar] [CrossRef]
- Hu, X.-J.; Yang, J.; Xie, X.-L.; Lv, F.-H.; Cao, Y.-H.; Li, W.-R.; Liu, M.-J.; Wang, Y.-T.; Li, J.-Q.; Liu, Y.-G.; et al. The Genome Landscape of Tibetan Sheep Reveals Adaptive Introgression from Argali and the History of Early Human Settlements on the Qinghai–Tibetan Plateau. Mol. Biol. Evol. 2019, 36, 283–303. [Google Scholar] [CrossRef] [Green Version]
- Barbato, M.; Hailer, F.; Orozco-terWengel, P.; Kijas, J.; Mereu, P.; Cabras, P.; Mazza, R.; Pirastru, M.; Bruford, M.W. Genomic Signatures of Adaptive Introgression from European Mouflon into Domestic Sheep. Sci. Rep. 2017, 7, 7623. [Google Scholar] [CrossRef] [Green Version]
- Meadows, J.R.S.; Hanotte, O.; Drögemüller, C.; Calvo, J.; Godfrey, R.; Coltman, D.; Maddox, J.F.; Marzanov, N.; Kantanen, J.; Kijas, J.W. Globally Dispersed Y Chromosomal Haplotypes in Wild and Domestic Sheep. Anim. Genet. 2006, 37, 444–453. [Google Scholar] [CrossRef]
- Meadows, J.R.S.; Kijas, J.W. Re-Sequencing Regions of the Ovine Y Chromosome in Domestic and Wild Sheep Reveals Novel Paternal Haplotypes. Anim. Genet. 2009, 40, 119–123. [Google Scholar] [CrossRef]
- Parmaksiz, A.; Oymak, A.; Yüncü, E.; Demirci, S.; Koban Baştanlar, E.; Özkan Ünal, E.; Togan, İ.; Özer, F. Türkiye’den 12 Yerli, Karagül, Karacabey Merinosu ve Anadolu Yaban Koyununda (Ovis gmelinii anatolica) Y-Kromozom Polimorfizmleri. Kafkas Univ. Vet. Fak. Derg. 2018, 24, 821–828. [Google Scholar] [CrossRef]
- Zhang, M.; Peng, W.-F.; Yang, G.-L.; Lv, F.-H.; Liu, M.-J.; Li, W.-R.; Liu, Y.-G.; Li, J.-Q.; Wang, F.; Shen, Z.-Q.; et al. Y Chromosome Haplotype Diversity of Domestic Sheep (Ovis aries) in Northern Eurasia. Anim. Genet. 2014, 45, 903–907. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, L.; Yan, W.; Li, S.; Wang, J.; Liu, X.; Hu, J.; Luo, Y. Y Chromosomal Haplotype Characteristics of Domestic Sheep (Ovis aries) in China. Gene 2015, 565, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Z.; Absher, D.M.; Tang, H.; Southwick, A.M.; Casto, A.M.; Ramachandran, S.; Cann, H.M.; Barsh, G.S.; Feldman, M.; Cavalli-Sforza, L.L.; et al. Worldwide Human Relationships Inferred from Genome-Wide Patterns of Variation. Science 2008, 319, 1100–1104. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.; Benecke, N.; Grabundžija, A.; Küchelmann, H.-C.; Pollock, S.; Schier, W.; Schoch, C.; Schrakamp, I.; Schütt, B.; Schumacher, M. The Textile Revolution. Research into the Origin and Spread of Wool Production between the Near East and Central Europe. eTopoi J. Anc. Stud. 2016, 6, 102–151. [Google Scholar]
- Sabatini, S.; Bergerbrant, S.; Brandt, L.Ø.; Margaryan, A.; Allentoft, M.E. Approaching Sheep Herds Origins and the Emergence of the Wool Economy in Continental Europe during the Bronze Age. Archaeol. Anthropol. Sci. 2019, 11, 4909–4925. [Google Scholar] [CrossRef] [Green Version]
- Kyselý, R. The Size of Domestic Cattle, Sheep, Goats and Pigs in the Czech Neolithic and Eneolithic Periods: Temporal Variations and Their Causes. Archaeofauna 2016, 25, 33–78. [Google Scholar]
- Bokonyi, S. The Development and History of Domestic Animals in Hungary: The Neolithic Through the Middle Ages. Am. Anthropol. 1971, 73, 640–674. [Google Scholar] [CrossRef]
- Landi, V.; Lasagna, E.; Ceccobelli, S.; Martinez, A.; Santos-Silva, F.; Vega-Pla, J.L.; Panella, F.; Allain, D.; Palhiere, I.; Murawski, M.; et al. An Historical and Biogeographical Assessment of European Merino Sheep Breeds by Microsatellite Markers. Small Rumin. Res. 2019, 177, 76–81. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Data Origin | Times of Divergence in Mya | ||
|---|---|---|---|---|
| Argali | Urial | Branch of A, B, D haplogroups | ||
| Rezaei et al. (2010) [13] | Cytochrome b sequence | 1.72 | 1.26 | - |
| Meadows et al. (2011) [33] | Whole mitogenome | 2.13 | - | 0.92 |
| Lv et al. (2015) [34] | Whole mitogenome | 2.93 | 2.60 | 0.89 |
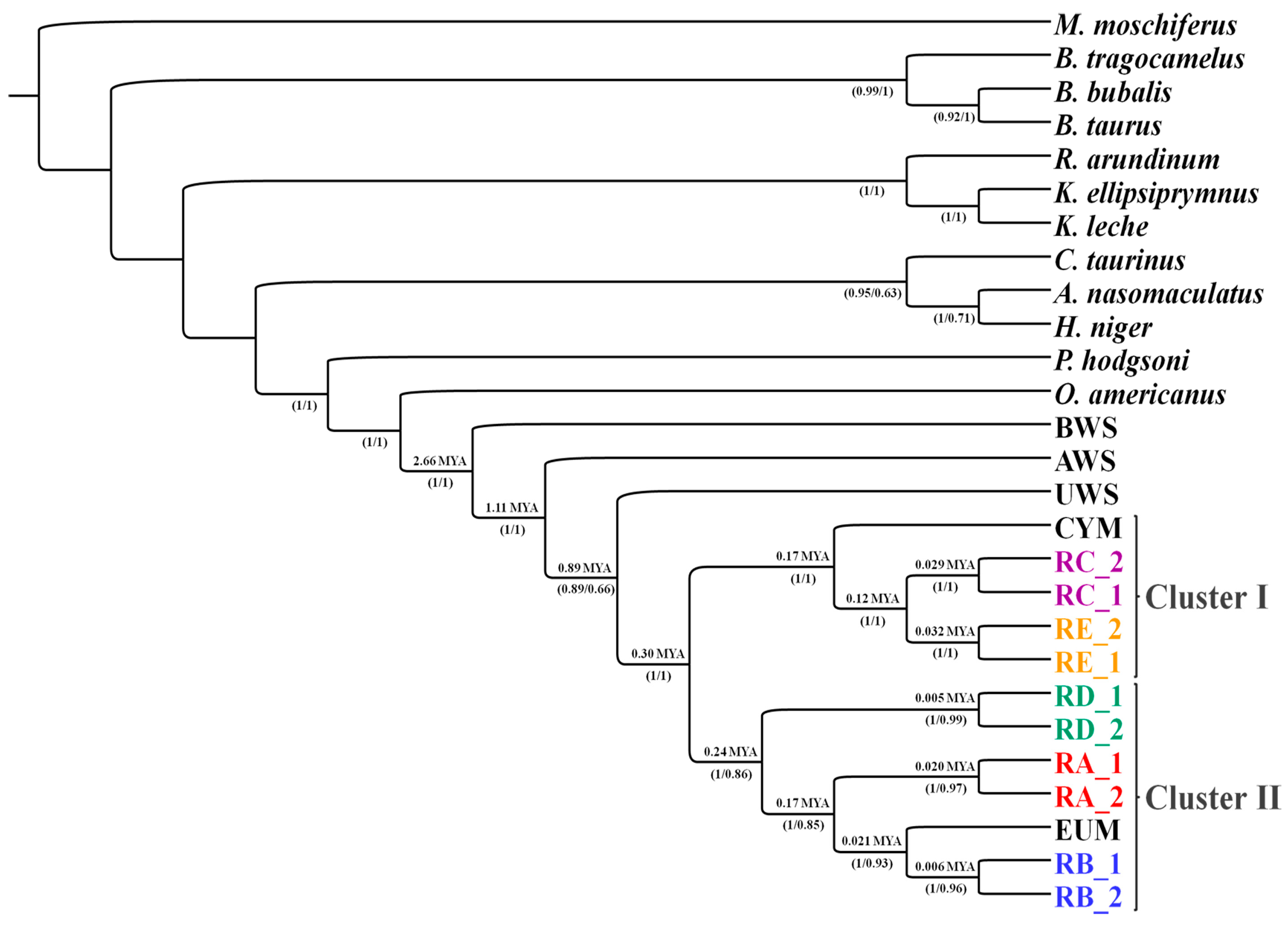
| Sanna et al. (2015) [32] | Whole mitogenome | 1.11 | 0.89 | 0.30 |
| Deng et al. (2020) [35] | Whole mitogenome | 2.93 | 2.60 | 1.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machová, K.; Málková, A.; Vostrý, L. Sheep Post-Domestication Expansion in the Context of Mitochondrial and Y Chromosome Haplogroups and Haplotypes. Genes 2022, 13, 613. https://doi.org/10.3390/genes13040613
Machová K, Málková A, Vostrý L. Sheep Post-Domestication Expansion in the Context of Mitochondrial and Y Chromosome Haplogroups and Haplotypes. Genes. 2022; 13(4):613. https://doi.org/10.3390/genes13040613
Chicago/Turabian StyleMachová, Karolína, Anežka Málková, and Luboš Vostrý. 2022. "Sheep Post-Domestication Expansion in the Context of Mitochondrial and Y Chromosome Haplogroups and Haplotypes" Genes 13, no. 4: 613. https://doi.org/10.3390/genes13040613
APA StyleMachová, K., Málková, A., & Vostrý, L. (2022). Sheep Post-Domestication Expansion in the Context of Mitochondrial and Y Chromosome Haplogroups and Haplotypes. Genes, 13(4), 613. https://doi.org/10.3390/genes13040613